Introduction
Glioma is the most common form of central nervous
system (CNS) neoplasm. This tumor is mostly expanded or
infiltrated, and has no clear boundary with normal brain tissues.
Therefore, it has a high degree of malignancy and poor prognosis
(1). In addition, according to the
World Health Organization (WHO) glioma grading criteria, gliomas
can be classified as low-grade (WHO I–II) and high-grade (WHO
III–IV) according to their degree of malignancy (2). Despite considerable advances in surgical
resection (or a biopsy if surgery cannot be performed),
radiotherapy and chemotherapy, the prognosis for glioma has not
significantly improved (2,3). Median survival time of patients with
glioblastoma multiforme, the most common and malignant type of
glioma, is only 14.6 months, and the 5-year survival rate is less
than 10% (4–6). In order to solve the problem of glioma
recurrence after surgical resection, gene therapy against glioma
has become a hot topic. Therefore, identification of tumor
biomarkers as target genes is essential for developing targeted
therapy against glioma (7). One
promising family for gene targeting is the collagen family.
According to the distribution in the body, collagen can be divided
into interstitial collagen, basement membrane collagen and
peripheral collagen. So far, 29 types of collagen have been
identified and described. A typical collagen molecule, also called
the triple helix, is made up of three left hand spirals (proline
II), intertwined and bonded to each other, forming a long and tough
right hand spiral structure (8–10).
COL3A1, also known as collagen α-1(III), is a
precursor of collagen III-α protein found in extensible connective
tissues such as skin, lung, and vascular system, and frequently in
association with type I collagen (COL1A1). An increase of COL3A1
and COL1A1 is found in some cancers, resulting as markers of poor
prognosis (11,12). COL3A1 is a fibrillary collagen
molecule that has been linked to myocardial infarction (13) and increased risk of stroke recurrence
and prognosis in Chinese patients (14). Interestingly, COL3A1 is the target of
miR29 family and downregulation of these microRNA (miRNA) family
members is responsible for the increased invasiveness of
nasopharyngeal carcinoma (15).
Let-7d suppresses growth, metastasis, and tumor macrophage
infiltration by targeting COL3A1 and CCL7 in renal cell carcinoma
(16). Additionally, an integrated
computational analysis of mutations regarding miRNA and mRNA
expression in a TCGA dataset of glioblastoma multiforme patients
revealed interesting insights into collagen regulation in glioma
(17). In our previous study, we also
revealed that COL3A1 and SNAP91 may serve as suitable biomarkers
for diagnostic or therapeutic strategies against glioma by using
bioinformatics (18). The current
research on COL3A1 is only in bioinformatics analysis. However, the
role of COL3A1 in tumorigenesis and progression, particularly in
glioma, is still limited.
Therefore, in this study, we examined the role of
COL3A1 in glioma tumorigenesis and progression. Our results showed
that COL3A1 expression was upregulated and directly correlated with
glioma grade. Using COL3A1-siRNA, we discovered that COL3A1
expression silencing resulted in an inhibition of cell
proliferation and migration. Additionally, COL3A1 mRNA levels were
inversely correlated with miR128-3p level in glioma. These findings
identify COL3A1 as an oncogene in glioma that could potentially
serve as a therapeutic target for glioma treatment.
Materials and methods
Cell culture and transient
transfection
Human glioma cell lines U87, U251, U343 and Hs683
were obtained from the Institute of Biochemistry and Cell Biology
of the Chinese Academy of Sciences (Shanghai, China) and maintained
as adherent cultures in Dulbecco's modified Eagle's medium (DMEM
C11995500BT; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS, 10099-141;
Gibco; Thermo Fisher Scientific, Inc.). Cells were cultured at 37°C
in a humidified atmosphere of 5% CO2. Cells were
detached using 0.25% trypsin when they were approximately 80%
confluent and consequently sub-cultured.
A COL3A1-siRNA and negative control siRNA (NC-siRNA)
were purchased from Santa-Cruz Biotechnology, Inc. (Dallas, TX, USA
(sc-43062; sc-637007). miRNA mimics and negative control miRNA were
purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
Cells were transfected using Lipofectamine® RNAiMAX
reagent (13778150; Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol.
Patient samples and quantitative
real-time PCR (qPCR)
The consecutive newly diagnosed 33 glioma patients
included in this study were recruited from the Affiliated Cancer
Hospital of XiangYa School of Medicine, Central South University
(Changsha, Hunan, China) and informed consent was obtained from all
patients prior to the beginning of the study (CTXY-1300041-3).
Samples were collected after surgical resection, snap-frozen in
liquid nitrogen and stored at −80°C before RNA extraction. Total
RNA was extracted by trizol reagent according to the manufacturer's
protocol. Two micrograms of RNA were reverse-transcribed into cDNA
using the Primescript RT reagent kit with gDNA Eraser (Takara Bio
Inc., Otsu, Japan). To increase the output of the microRNAs, RNA
was reverse-transcribed into cDNA using Mir-X™ miRNA FirstStrand
Synthesis kits (Clontech Laboratories, Inc., Mountainview, CA,
USA). qPCR was performed using SYBR Premix DimerEraser kit (Takara
Bio Inc.). qPCR conditions were the following: 95°C for 30 sec, 40
cycles at 95°C for 5 sec, 55°C for 30 sec and 72°C for 30 sec.
Primers used for qPCR were as follows: COL3A1 forward,
TTGAAGGAGGATGTTCCCATCT; reverse, ACAGACACATATTTGGCATGGTT. GAPDH
forward, GAGTCAACGGATTTGGTCGT; reverse, TTGATTTTGGAGGGATCTCG. U6
forward, CTCGCTTCGGCAGCACA; reverse, AACGCTTCACGAATTTGCGT. Mature
miRNA was used as miRNA specific 5′ primer. The 3′ primer for qPCR
was supplied with the kit. All the analyses were performed in
duplicate. The relative expression of COL3A1 mRNA/microRNA was
normalized to the expression level of GAPDH/U6 mRNA using the
2−ΔCq method (19).
Immunohistochemistry
Paraffin embedded glioma tissues were cut into 5 µm
sections and stained to evaluate COL3A1 expression. The whole
process was performed according to standard protocols. The primary
antibody (HPA007583; Sigma-Aldrich, St. Louis, MO, USA) was diluted
to 1:100 and incubated overnight at 4°C. Color development was
performed by using the DAB kit (P0203; Beyotime Biotechnology,
Shanghai, China) according to the manufacturer's protocol. Two
pathologists evaluated the immunohistochemistry results
independently, according to the percentage of COL3A1 positive cells
per field of vision at ×400 magnification. Positive cell rate was
measured by Image pro-plus 6.0 software.
Western blot analysis
Total cell protein extracts were obtained using RIPA
buffer purchased from Beyotime Biotechnology. Protein lysates were
separated by centrifugation, and the concentrations were qualified
using Pierce BCA Protein assay kit (P0012; Beyotime Biotechnology).
Protein lysates were loaded into a 10% SDS-PAGE gel. The gel was
transferred to a 0.45 µm PVDF membrane (Millipore, Billerica, MA,
USA) for 2 h. Primary antibodies anti-COL3A1 (1:1,000, HPA007583;
Sigma-Aldrich), and anti-GAPDH (1:5,000, G8795; Sigma-Aldrich) were
incubated overnight at 4°C. Secondary HRP-conjugated antibodies
(A0208/A0216; Beyotime Biotechnology) were added at 1:2,000
dilution and incubated for 1 h at 25°C. The bound antibodies were
visualized using an enhanced chemiluminescence reagent (RPN2232; GE
Healthcare Life Sciences, Little Chalfont, UK) and quantified by
densitometry using ChemiDoc XRS+ image analyzer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Densitometric band analyses
were adjusted to the GAPDH loading control.
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]
MTS assay
Hs683 and U251 were seeded onto 96-well culture
plates (1,000 cells/well/100 µl) and cell proliferation was
measured at 24, 48, 72, and 96 h after transfection using a MTS
reagent (1:9; Promega Corporation, Madison, WI, USA) according to
the manufacturer's instructions. Absorbance was measured at 490 nm
by the BioTek®Eon (Synergy™ HT; BioTek Instruments,
Inc., Winooski, VT, USA). Background absorbance of the medium was
subtracted.
Colony formation assay
U251 were transfected with siRNA for 48 h and were
collected and seeded in triplicate into 6-well plates at a density
of 1,000 cells/well/ml. Cells were incubated for 10 days at 37°C
under 5% CO2 atmosphere. Cells were then fixed with 4%
paraformaldehyde for 30 min and stained with Giemsa (C0121;
Beyotime Biotechnology) for 20 min. After washing with
double-distilled H2O several times, images from the cell
plates were obtained using a camera (Canon, Inc., Tokyo,
Japan).
Wound healing assay
U251 were seeded onto a 6-well plate overnight. The
confluent monolayers were scratched using sterile pipette tips and
washed with phosphate-buffered saline (PBS) several times to remove
detached cells. Cells were then transfected with siRNA for 48 h in
medium without serum. Photographs of the wounded areas were
obtained using a Leica DMI3000 B inverted microscope (Leica
Microsystems GmbH, Wetzlar, Germany). The migration rate was
calculated according to the scratched surfaces, which were
quantified using ImageJ Version 1.41o software (National Institutes
of Health, Bethesda, MD, USA).
Bioinformatics analysis
Three online miRNA research databases, TargetScan
(http://www.targetscan.org), microRNA.org (http://34.236.212.39/microrna/home.do), and miRBase
(http://www.mirbase.org/), were used to predict
miRNA regulators of the COL3A1 gene. The predicted miRNAs were
selected based on the coincident results of the three databases.
Overlapping results were extracted (excluding those that have been
studied) for subsequent analysis.
Dual luciferase activity assay
U251 cells were seeded onto a 24-well plate
overnight. Cells were co-transfected with pmiR-RB-reporter vectors
(WT/MUT) and miRNA mimics (Guangzhou RiboBio Co., Ltd.) using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
After 48 h, luciferase activity was measured using the Dual
Luciferase Reporter assay system (Promega). Firefly luciferase
activity was normalized to renilla luciferase activity for each
transfected well. The results were obtained from three independent
experiments and each one was performed in triplicate.
Statistical analysis
SPSS 19.0 software was used for statistical
analyses. The significance of the differences two groups was
estimated by Student's t-test. One-way ANOVA followed by Tukey's
post hoc test was used to analyze differences between three or more
groups. The association between COL3A1 and clinical features was
analyzed using Chi-square test. Pearson correlation analyses were
performed to investigate the correlation between COL3A1 and
miR128-3p. All tests performed were two-sided and statistical
significance was set at P<0.05.
Results
COL3A1 is strongly upregulated in
glioma
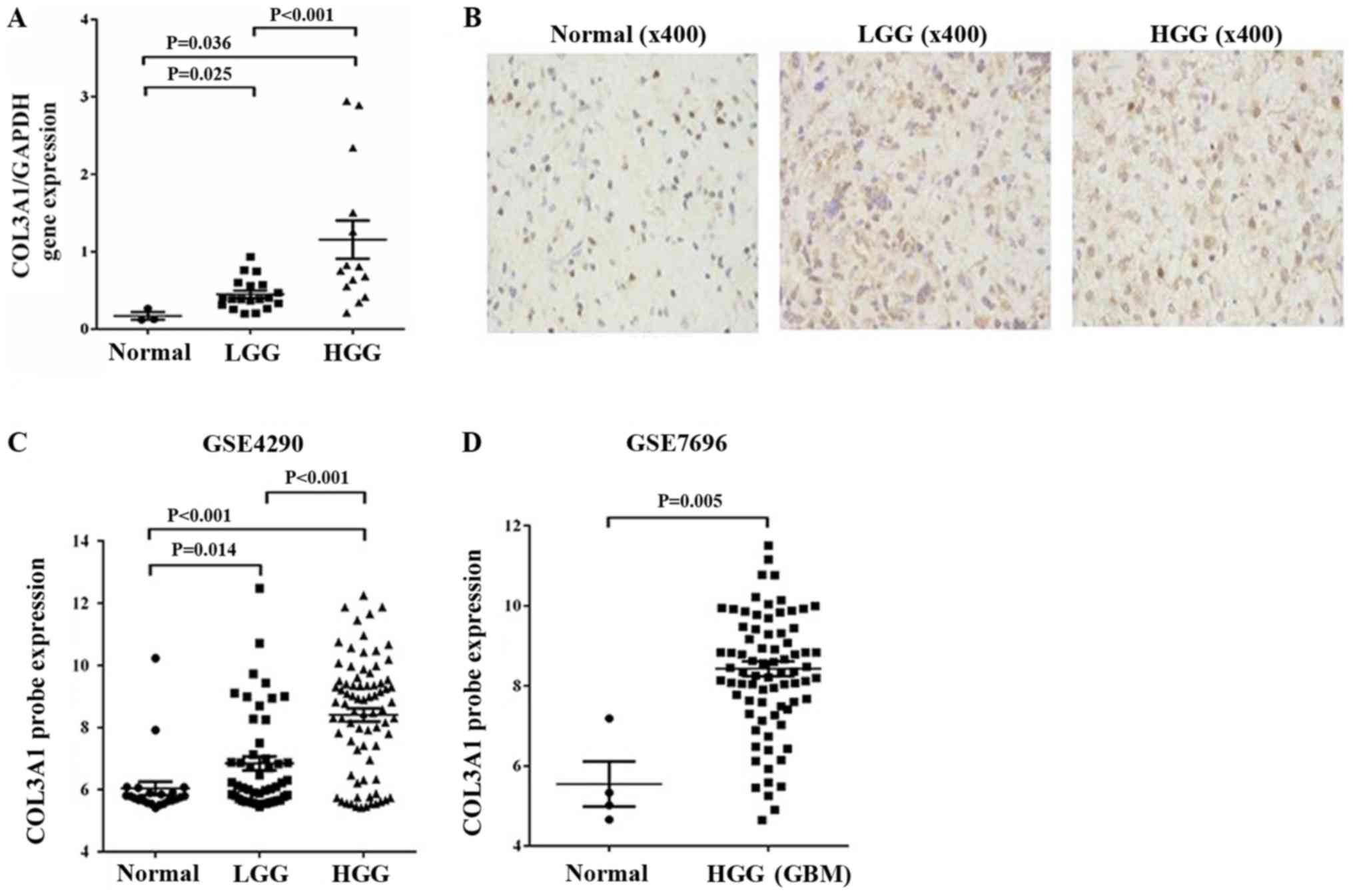
To evaluate if COL3A1 is upregulated in glioma and
directly correlated with glioma grade, we examined COL3A1 in 33
glioma tissues and normal brain tissues using quantitative qPCR.
COL3A1 expression was significantly higher in glioma tissues [low
grade glioma (LGG), P=0.025; high grade glioma (HGG), P=0.036]
compared with normal brain samples (Fig.
1A). Thirty-three glioma tissues were divided into two group,
based on the median expression level of all gliomas. Chi-square
test was performed to analyze the data shown in Table I. As summarized, COL3A1 was
significantly associated with WHO grade (LGG vs. HGG, P=0.0008).
Further examination of COL3A1 protein expression (positive cell
rate) resulted in ‘normal brain’ (7.71%), LGG (18.58%), HGG
(24.03%), separately (Fig. 1B). In
our previous study, we used TCGA data to verify previous COL3A1
expression. Here we evaluated its expression in GSE4290 and GSE7696
expression profiles acquired from the Gene Expression Omnibus (GEO,
http://www.ncbi.nlm.nih.gov/geo/)
database. Upregulated COL3A1 (>2-fold upregulated) expression
was observed in malignant gliomas compared to LGG and non-tumor
brain tissue and resulted directly correlated with the glioma grade
in GSE4290 dataset (Fig. 1C).
Analysis of the GSE7696 data also revealed a >2-fold COL3A1
upregulation at the transcription level and resulted drastically
increased in malignant gliomas when compared to non-tumor brain
tissue (Fig. 1D). These results
suggested a correlation between increased COL3A1 expression and
glioma tumor grade.
 | Table I.Correlation between COL3A1 expression
and glioma clinicopathologic features in 33 patients. |
Table I.
Correlation between COL3A1 expression
and glioma clinicopathologic features in 33 patients.
|
|
| COL3A1 expression
levels |
|
|
|---|
|
|
|
|
|
|
|---|
| Characteristic | N (%) | High | Low | Ratio (high/low) | P-value |
|---|
| Sex |
|
|
|
|
|
| Male | 12 (36.36) | 4 | 8 | 0.500 | 0.462 |
|
Female | 21 (63.64) | 5 | 16 | 0.312 |
|
| Age, years |
|
|
|
|
|
|
<45 | 16 (48.49) | 5 | 11 | 0.454 | 0.257 |
| ≥45 | 17 (51.51) | 4 | 13 | 0.307 |
|
| Grade |
|
|
|
|
|
| Low
(I+II) | 19 (57.57) | 3 | 16 | 0.187 | 0.0008 |
| High
(III+IV) | 14 (42.43) | 6 | 8 | 0.750 |
|
SiRNA inhibits COL3A1 expression
efficiently in human glioma cell lines
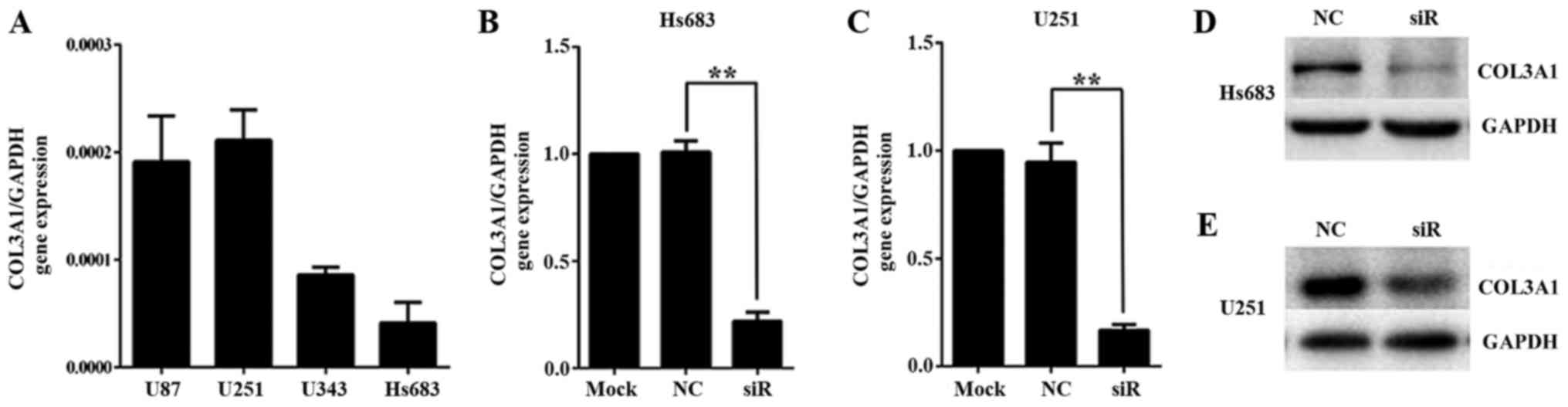
To investigate the role of COL3A1 in glioma, we
knocked down COL3A1 expression using a COL3A1-siRNA (Fig. 2). COL3A1 gene expression was higher in
U87 and U251, and the lowest in Hs683 (Fig. 2A). After transfection of Hs683 and
U251 with COL3A1-siRNA or NC-siRNA, COL3A1 mRNA expression was
significantly reduced by ~80% and protein expression in Hs683
(Fig. 2B and D) and U251 (Fig. 2C and E) was nearly depleted.
Therefore, the present results demonstrated that a highly efficient
COL3A1 knockdown at the mRNA and protein level was achieved after
transfection.
COL3A1 knockdown suppresses
proliferation and migration in glioma cells
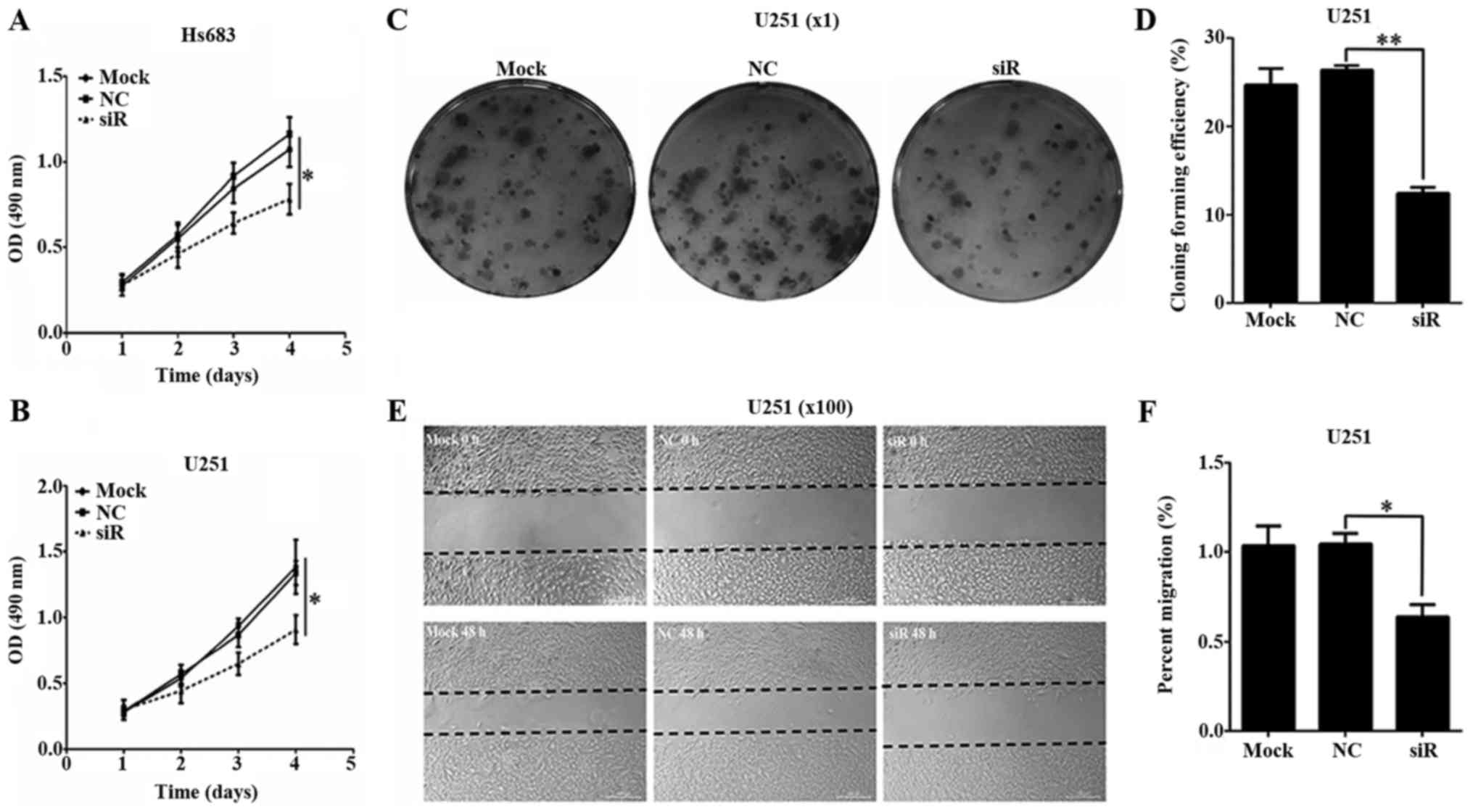
To investigate the effect of COL3A1 silencing on
glioma cell proliferation, Hs683 and U251 transfected with
COL3A1-siRNA or NC-siRNA were analyzed by MTS assay. Proliferation
of Hs683 cells treated with COL3A1-siRNA began to decrease 72 h
after transfection. On day 4, proliferation of silenced cells was
significantly reduced compared to those treated with NC-siRNA or
untreated cells (P<0.05; Fig. 3A).
These results were consistent with the results obtained in U251
cells (P<0.05; Fig. 3B). Moreover,
no significant difference was observed between NC-siRNA treated
cells and untreated cells. The relatively long-term effects of
COL3A1 knockdown on glioma cell proliferation were also examined
using a colony formation assay. As shown in Fig. 3C and D, COL3A1 silencing in U251 cells
substantially reduced colony formation (P<0.01).
Next, we investigated the effect of COL3A1 silencing
on cell migration. COL3A1 knockdown dramatically reduced glioma
cell migration 48 h after transfection with COL3A1-siRNA (Fig. 3E). Migration rates were significantly
lower in silenced cells compared to untreated and NC-siRNA treated
cells (P<0.05; Fig. 3F). No
significant differences were found between untreated and NC-siRNA
treated cells. These results suggested that COL3A1 contributed to
glioma cell migration. Taken together, MTS, Colony formation assay
and Wound healing assay were used to illustrate the relationship
between COL3A1 and cell proliferation in Fig. 3. These findings identify COL3A1 as an
oncogene in glioma.
miR128-3p overexpression inhibits
COL3A1 mRNA and protein expression
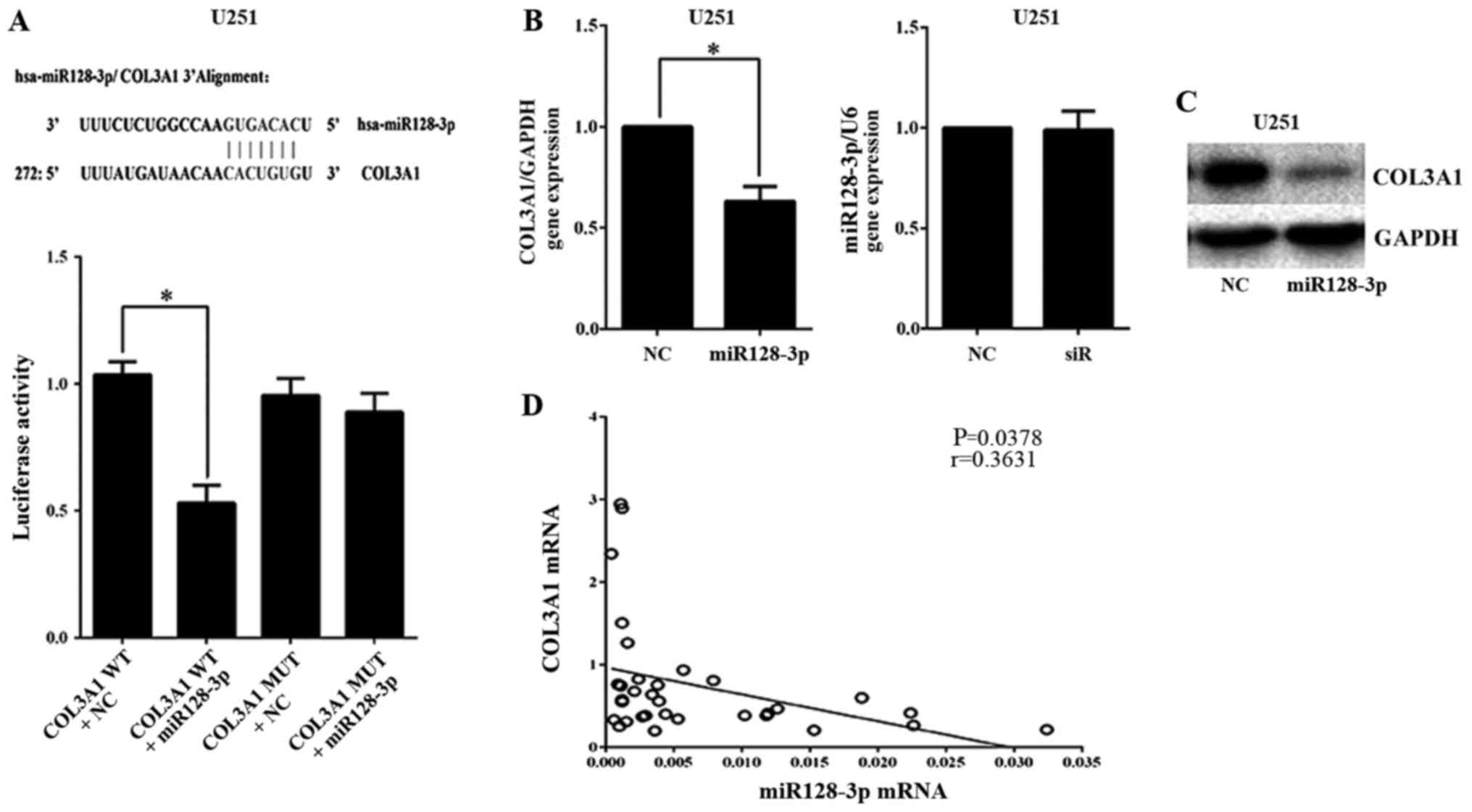
To identify potential miRNA regulators of the COL3A1
gene, three online miRNA research databases, TargetScan, microRNA.org, and miRBase, were analyzed using
internetwork. Among the predicted miRNAs, we were particularly
interested in miR128-3p because of our previous results indicating
this miRNA as associated with COL3A1. To validate miR128-3p
binding, we identified a putative consensus site for miR128-3p
binding the 3UTR of COL3A1 using microRNA.org.
Furthermore, luciferase activity decreased significantly in the
cells transfected with the wild-type reporter plasmids but not in
those transfected with mutant miR128-3p plasmids in U251 cells
(P<0.05; Fig. 4A). Moreover,
COL3A1 mRNA was inhibited by miR128-3p overexpression (P<0.05),
while miR128-3p mRNA showed no significant differences when
transfection with COL3A1-siRNA was performed in U251 cells
(Fig. 4B). COL3A1 protein was also
inhibited by the miR128-3p mimics in U251 cells (Fig. 4C). Additionally, COL3A1 mRNA levels
were inversely correlated with miR128-3p level in glioma tissues
(Fig. 4D).
Discussion
Gliomas is one of the most common malignant tumors
with high recurrence, fatality and low recovery rates. Glioma cells
are highly invasiveness and infiltrative multiple brain regions
(20–23). Targeting specific tumor-promoting
genes may provide a promising alternative treatment to the current
standards (24). Collagen is a
biological macromolecular protein synthetized and widely present in
animal cells, which content and sort is various. It has a
significant impact on cells, tissues and organs. Previously, we
identified COL3A1 and SNAP91 as potential biomarkers for diagnostic
or therapeutic strategies against glioma using bioinformatics
(18). In the current study, we
confirmed that COL3A1 was upregulated in glioma by analyzing COL3A1
expression in 33 glioma samples and two datasets (GSE4412,
GSE7696). Additionally, silencing COL3A1 expression in vitro
inhibited glioma cell proliferation and migration. This might be
closely related to the following three primary functions of
collagen in brain tumors: to act as: i) a scaffold and provide
sites for cell adhesion; ii) a reservoir for extracellular matrix
proteins, proteoglycans and growth factors; and iii) a ligand to
activate signal transduction networks required for tumor growth,
differentiation and invasion (25–29).
Although the underlying mechanism for these effects still needs to
be studied, our results gave a hint on the role of COL3A1 in
collagen and brain tumor. Taken together, the results suggested
that COL3A1 functions as an oncogene in glioma.
Interestingly, COL3A1 is the target of the miR29
family members, and downregulation of these members is responsible
for the increased invasiveness of nasopharyngeal carcinoma
(15). Boxing Su et al
validated COL3A1 as a direct, functional target gene for let-7d in
renal cell carcinoma, in vitro and in vivo (16). Our results also found that the 3′UTRs
in COL3A1 were the target sites through which miR128-3p modulates
the expressions of COL3A1. Importantly, COL3A1 mRNA was inversely
correlated with miR128-3p levels in glioma clinical specimens. Our
findings suggested that miR128-3p-COL3A1 regulatory pathway plays a
role in glioma growth. These findings indicated COL3A1 as an
oncogene in glioma that could potentially serve as a target in the
therapy for glioma patients.
Since COL3A1 is involved in the invasion of
nasopharyngeal carcinoma progression (15), invasion studies should be conducted.
In addition, any candidate miRNA, which may involve the COL3A1
should be taken into consideration. Unfortunately, because of the
financial strain, in the current research there is a lack of data
on miR128-3p expression in glioma tissue and invasion studies. We
mainly discussed the role of COL3A1 in glioma. We have done the MTS
test with U87 and the results was consistent with the trend of U251
and Hs683 (data not shown). Although U87 cells (that matches U87 MG
ATCC cells) have been reported to be misidentified (30), we confirm that such misidentification
issue is unlikely to affect the outcomes of our study. In addition,
due to the lack of patient's prognostic information, survival
analysis could not be performed. For this reason, we will continue
to collect data regarding the patient's prognostic information.
Then, future studies should be performed in glioma cells on other
COL3A1 miRNA regulators reported in other tumors.
Although glioma gene therapy is beneficial for
patients, it is still under test and did not achieve the desired
effect in clinical trials. As a result, an improvement of gene
therapy clinical application to ameliorate the effect and the
development of more efficient methods of using gene therapy is the
focus of our future research. Our goal is a continuous gene therapy
innovation to provide more scientific and feasible strategies for
the clinical treatment of glioma.
Acknowledgements
Not applicable.
Funding
This study was supported by the Key specialty of
clinical pharmacy of Hunan administration of traditional Chinese
medicine (grant no. 2017-11), the Education Bureau of Hunan
Province (grant no. 17A156) and the Hunan administration of
traditional Chinese medicine (grant no. 201709).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RO and YG conceived the project, designed
experiments, analyzed data, and wrote the manuscript. YG, TZ, JC,
and LL performed experiments and analyzed data.
Ethics approval and consent to
participate
All glioma human samples were taken in accordance
with the ethical guidelines of the Affiliated Cancer Hospital of
XiangYa School of Medicine, Central South University. The present
study was approved by the Ethics Committee of Central South
Unviersity and written informed consent was obtained from all
patients prior to their inclusion within the present study.
Consent for publication
All patients agreed to the publication of their
data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dodgshun AJ, Maixner WJ, Heath JA,
Sullivan MJ and Hansford JR: Single agent carboplatin for pediatric
low-grade glioma: A retrospective analysis shows equivalent
efficacy to multiagent chemotherapy. Int J Cancer. 138:481–488.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gao YF, Zhu T, Mao CX, Liu ZX, Wang ZB,
Mao XY, Li L, Yin JY, Zhou HH and Liu ZQ: PPIC, EMP3 and CHI3L1 are
novel prognostic markers for high grade glioma. Int J Mol Sci.
17(pii): E18082016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Karsy M, Guan J, Sivakumar W, Neil JA,
Schmidt MH and Mahan MA: The genetic basis of intradural spinal
tumors and its impact on clinical treatment. Neurosurg Focus.
39:E32015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Batista A, Riedemann L, Vardam T and Jain
RK: Targeting the tumor microenvironment to enhance pediatric brain
cancer treatment. Cancer J. 21:307–313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Domingues P, González-Tablas M, Otero Á,
Pascual D, Miranda D, Ruiz L, Sousa P, Ciudad J, Gonçalves JM,
Lopes MC, et al: Tumor infiltrating immune cells in gliomas and
meningiomas. Brain Behav Immun. 53:1–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Panditharatna E, Yaeger K, Kilburn LB,
Packer RJ and Nazarian J: Clinicopathology of diffuse intrinsic
pontine glioma and its redefined genomic and epigenomic landscape.
Cancer Genet. 208:367–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tobias A, Ahmed A, Moon KS and Lesniak MS:
The art of gene therapy for glioma: A review of the challenging
road to the bedside. J Neurol Neurosurg Psychiatry. 84:213–322.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dodd A and Daniels TR: Injectable
recombinant human platelet-derived growth factor in collagen
carrier for hindfoot fusion. Foot Ankle Clin. 21:777–791. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang S, Vader D, Wang Z,
Stemmer-Rachamimov A, Weitz DA, Dai G, Rosen BR and Deisboeck TS:
Using magnetic resonance microscopy to study the growth dynamics of
a glioma spheroid in collagen I: A case study. BMC Med Imaging.
8:32008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liang Y, Diehn M, Bollen AW, Israel MA and
Gupta N: Type I collagen is overexpressed in medulloblastoma as a
component of tumor microenvironment. J Neurooncol. 86:133–141.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Z, Wang Y, Zhang J, Zhong J and Yang
R: COL1A1 promotes metastasis in colorectal cancer by regulating
the WNT/PCP pathway. Mol Med Rep. 17:5037–5042. 2018.PubMed/NCBI
|
|
12
|
Wang XQ, Tang ZX, Yu D, Cui SJ, Jiang YH,
Zhang Q, Wang J, Yang PY and Liu F: Epithelial but not stromal
expression of collagen alpha-1(III) is a diagnostic and prognostic
indicator of colorectal carcinoma. Oncotarget. 7:8823–8838.
2016.PubMed/NCBI
|
|
13
|
Kong CH, Lin XY, Woo CC, Wong HC, Lee CN,
Richards AM and Sorokin VA: Characteristics of aortic wall
extracellular matrix in patients with acute myocardial infarction:
tissue microarray detection of collagen I, collagen III and elastin
levels. Interact Cardiovasc Thorac Surg. 16:11–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lv W, Lin Y, Song W, Sun K, Yu H, Zhang Y,
Zhang C, Li L, Suo M, Hui R and Chen J: Variants of COL3A1 are
associated with the risk of stroke recurrence and prognosis in the
Chinese population: A prospective study. J Mol Neurosci.
53:196–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qiu F, Sun R, Deng N, Guo T, Cao Y, Yu Y,
Wang X, Zou B, Zhang S, Jing T, et al: miR-29a/b enhances cell
migration and invasion in nasopharyngeal carcinoma progression by
regulating SPARC and COL3A1 gene expression. PLoS One.
10:e01209692015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su B, Zhao W, Shi B, Zhang Z, Yu X, Xie F,
Guo Z, Zhang X, Liu J, Shen Q, et al: Let-7d suppresses growth,
metastasis, and tumor macrophage infiltration in renal cell
carcinoma by targeting COL3A1 and CCL7. Mol Cancer. 13:2062014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong H, Luo L, Hong S, Siu H, Xiao Y, Jin
L, Chen R and Xiong M: Integrated analysis of mutations, miRNA and
mRNA expression in glioblastoma. BMC Syst Biol. 4:1632010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao YF, Mao XY, Zhu T, Mao CX, Liu ZX,
Wang ZB, Li L, Li X, Yin JY, Zhang W, et al: COL3A1 and SNAP91:
Novel glioblastoma markers with diagnostic and prognostic value.
Oncotarget. 7:70494–70503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xue S, Song G and Yu J: The prognostic
significance of PD-L1 expression in patients with glioma: A
meta-analysis. Sci Rep. 7:42312017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xue J, Zhao Z, Zhang L, Xue L, Shen S, Wen
Y, Wei Z, Wang L, Kong L, Sun H, et al: Neutrophil-mediated
anticancer drug delivery for suppression of postoperative malignant
glioma recurrence. Nat Nanotechnol. 12:692–700. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang P and Heitman J: The cyclophilins.
Genome Biol. 6:2262005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qi XT, Zhan JS, Xiao LM, Li L, Xu HX, Fu
ZB, Zhang YH, Zhang J, Jia XH, Ge G, et al: The unwanted cell
migration in the brain: Glioma metastasis. Neurochem Res.
42:1847–1863. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan YC, Cui CC, Zhu YS, Zhang L, Shi M, Yu
JS, Bai J and Zheng JN: Overexpression of CAP1 and its significance
in tumor cell proliferation, migration and invasion in glioma.
Oncol Rep. 36:1619–1625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lucey P and Goldberg DJ: Complications of
collagen fillers. Facial Plast Surg. 30:615–622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Raglow Z and Thomas SM: Tumor matrix
protein collagen XIα1 in cancer. Cancer Lett. 357:448–453. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Konstantopoulos A and Mehta JS:
Conventional versus accelerated collagen cross-linking for
keratoconus. Eye Contact Lens. 41:65–71. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jokerst C, Purdy H and Bhalla S: An
overview of collagen vascular disease-associated interstitial lung
disease. Semin Roentgenol. 50:31–39. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Snedeker JG and Gautieri A: The role of
collagen crosslinks in ageing and diabetes-the good, the bad, and
the ugly. Muscles Ligaments Tendons J. 4:303–308. 2014.PubMed/NCBI
|
|
30
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|