Introduction
Hepatoblastoma (HB) is one of the most common solid
tumors, which caused cancer-associated mortality worldwide
(1). As the most common malignant
liver tumor of childhood, the incidence peaks in the first two
years of life, with the majority of cases presenting by 5 years of
age. Slight male preponderance is observed. HB may be present at
birth and prenatal cases have been reported (2,3). The
molecular pathogensis of HB is remained to be elucidated. Different
molecular alterations have been identified as being involved in the
genesis of HB. Particularly, a deregulation of different signaling
pathways has been described among which Wnt signaling (4–6), Sonic
Hedgehog (7), Notch and
phosphatidylinositol 3-kinase/protein kinase B/mammalian target of
rapamycin are counted as the main players (8,9). It is of
great importance to understand the molecular mechanisms to explore
more targeted therapeutic strategies. HepG2 cells were isolated
from a human liver biopsy of a 15-year old male with HB, which were
shown to be a HB cell line (10–12).
DNA-damage regulated autophagy modulator 1 (DRAM1)
is an evolutionarily conserved transmembrane protein, which
localizes predominantly to lysosomes and acts as a target of tumor
protein p53 (TP53)-mediated autophagy and programmed cell death
(13). Recent reports have
demonstrated that high levels of DRAM1 were associated with shorter
overall survival in glioblastoma multiforme (GBM) patients, and
knockdown of DRAM1 inhibited the migration and invasion abilities
of glioblastoma stem cells (GSCs) (14). However, the precise contribution of
DRAM1 to cancer cell invasion and migration and the underlying
mechanisms remain unclear. Autophagy is an evolutionally conserved
process in which amino acids, nutrients, and lipids are recycled
when cells go through nutrient and oxygen deprivations (15). DRAM1 acts as a regulator of autophagy
mediated by p53 in response to genotoxic stresses through
regulation of the clearance of autophagosomes by promoting
lysosomal acidification and inducing the activation of lysosomal
enzymes (16).
Epithelial-mesenchymal-transition (EMT) represents a
process of fast changes, during which the cell phenotype changes
from epithelial to mesenchymal, the expression of mesenchymal
markers were upregulated, the actin cytoskeleton was
reorganization, cell-cell adhesion structures were destructed, and
pseudopod formation emerges (17).
During the EMT, the migratory ability of epithelial cells is
enhanced (17). Autophagy is a
catabolic process that mediates degradation of unnecessary or
dysfunctional cellular components (18,19). These
two important processes in cancer are linked in an intricate
relationship. EMT requires autophagy to support viability of
potentially metastatic cancer cells, while a number of additional
evidence indicates that autophagy acts to prevent EMT and that the
activation of the autophagic machinery may determine reversion of
the EMT phenotype in cancer cells (20–24).
This study was designed to investigate the effect of
DRAM1 on cell invasion and migration, as well as explore the
underlying mechanisms involved in cell autophagy and EMT. We
provided the evidence that DRAM1 knockdown inhibited cell invasion
and migration abilities of HB cells by inhibiting the autophagy-EMT
pathway.
Materials and methods
Antibodies and reagents
Antibodies for E-Cadherin and Vimentin were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Antibodies for DRAM1, P62 and LC3 were purchased from Abcam
(Cambridge, MA, USA). Antibody for β-actin was purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). Rapamycin was purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Transwell
chamber was purchased from Corning Incorporated (Corning, NY, USA).
Matrigel was purchased from BD Biosciences (Franklin Lakes, NJ,
USA).
Cell culture
Human HB derived HepG2 cells were obtained from the
American Type Culture Collection and cultured in Dulbecco's
modified Eagle's medium (DMEM; 11965500; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (FBS; 086150008; Wisten Inc., Anthem, AZ, USA), 100 IU/ml
penicillin, and 100 IU/ml streptomycin in a humidified incubator at
37°C under 5% CO2 atmosphere, and passaged at
preconfluent densities using 0.25% trypsin solution every 2 to 3
days. Cells were stored and used within 3 months after
resuscitation from cryopreservation status.
Transfection and RNA interference
The DRAM1 shRNA (shDRAM1: Tracking number
TRCN0000161451, clone ID NM 018370.1–1356s1c1) were purchased from
Sigma-Aldrich and the small interfering (si)RNA targeting DRAM1 was
purchased from Shanghai GenePharma Co., Ltd. (Shanghai, China). For
establish stable DRAM1 knockdown cells, HepG2 cells were
transfected with DRAM1 shRNA and were selected in cell culture
medium containing 1.5 mg/ml poromycin for 1 week. Cells were then
cultured in culture medium for in vivo experiments. For
transfection, cells were plated in 6-well plates at 30% confluency,
and siRNA duplexes were introduced into the cells using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's recommendation. The siRNAs
targeting sense sequences were as follows: DRAM1-1,
5′-CCACGAUGUAUACAAGAUA-3′; DRAM1-2, 5′-CCACGAAAUCAAUGGUGA-3′;
Negative control, 5′-UAAGGCUAUGAAGAGAUAC-3′. The knockdown
efficiency of specific proteins was determined by western blot
analysis 48 h after shRNA or siRNA treatment.
Short hairpin RNA (shRNA) lentiviruses
production and transduction
A shRNA against DRAM1 was cloned into the pLKO.1
vector according to the manufacturer's protocol (Addgene, Inc.,
Cambridge, MA, USA). pLKO.1, scrambled shRNA (negative control),
pMD2.G (used for viruspackaging) and psPAX2 (used for virus
packaging) were purchased from Addgene, Inc. All constructs were
verifed by sequencing. Lentiviruses were produced by
co-transfecting 293FT cells (Invitrogen; Thermo Fisher Scientifc,
Inc.) in 10-cm dishes with 10 µg pLKO.1-shRNA, 2.5 µg pMD2.G and
7.5 µg psPAX2 using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). HepG2 cells were infected with DRAM1 shRNA.
Virus-containing medium was removed after 16 h and replaced with
fresh DMEM. After 24 h, the cells were used experimentally.
Transwell assay
The migration and invasion abilities were detected
using Transwell chambers with 8 µm pore filters. The filters used
for invasion assays were coated with 30 µl pre-diluted Matrigel
(diluted at a ratio of 1:6 with serum-free DMEM medium). Cells
(1×105 cell/well for migration assay, 2.5×105
cell/well for invasion assay) were appropriately added to the upper
chambers. And then 0.6 ml DMEM medium supplemented with 10% FBS was
added to the lower chambers as a chemoattractant. After incubation
for 24 h, migrated or invaded cells were stained with DAPI. The
cell number was counted under an optical microscope (5 different
visual fields were randomly selected for each membrane and observed
under ×600 magnification lenses). All experiments were performed at
least in triplicate.
Western blot analysis
Protein was extracted from cells using cell lysis
solutions containing protease inhibitors and phosphorylase
inhibitors. Equal amounts of protein were fractionated on
Tris-glycine SDS-polyacrylamide gels and subjected to
electrophoresis and transferred to NC membranes. Membranes were
blocked with 5% non-fat milk with Tris buffered Saline-Tween-20
(TBS-T), and then incubated with primary antibodies against DRAM1,
P62, LC3, E-Cadherin, or Vimentin. After washing in TBST, membranes
were incubated with fluorescent secondary antibodies. β-actin was
used as the loading control. Immunoreactivity was detected using
ODYSSEY INFRARED IMAGER (Li-COR Biosciences, Nebraska, NE, USA).
The signal intensity of primary antibody binding was quantitatively
analyzed with Image J software (W.S. Rasband, Image J; National
Institutes of Health, Bethesda, MD, USA).
Immunofluorescence
The HepG2 cells were seeded onto cover glass in 24
well plates and cultured to the appropriate confluency. Thereafter,
cells were washed with phosphate-buffered saline (PBS) for 5 min ×3
times. Then cells were treated with precooled alcohol for 15 min
before blocked in PBS containing 1% BSA and 0.1% Triton X-100 for 1
h at room temperature. Then the cells were incubated with primary
antibody overnight at 4°C. After washed 3 times with PBS for 10
min, the cells were incubated with Cy3-conjugated donkey
anti-rabbit IgG for 1 h at room temperature. After another 10 min ×
3 times of washing with PBS, cells were incubated with DAPI for 10
min, and dehydrated in increasing grades of ethanol and
cover-slipped with Fluoromount Aqueous Mounting Medium (Sigma
F4680; Sigma-Aldrich; Merck KGaA). The slices were analyzed with a
laser scanning confocal unit (Zeiss LSM 710; Carl Zeiss, Jena,
Germany).
In vivo tumor growth analysis
The control shRNA-transfected cells and DRAM1
shRNA-transfected cells (1×107) were intravenously
injected into the tail vein of 6-week-old female athymic nude mice
(Shanghai SLAC Laboratory Animal Co. Ltd., Shanghai, China). At 4
weeks later, the mice were anesthetized and photographed. After the
mice were sacrificed, the livers were removed and photographed. All
animal procedures were approved and monitored by the local Animal
Care and Use Committee in Soochow University (License no. Syxk;
Su-0062).
Statistical analysis
All data were presented as means ± SEM. Data were
subjected to one-way ANOVA using the GraphPad Prism software
statistical package (GraphPad Software; GraphPad Software, Inc., La
Jolla, CA, USA). When a significant group effect was found, post
hoc comparisons were performed using the Student-Newman-Keuls test
to examine special group differences. Independent group t-tests
were used for comparing two groups. Significant differences with
P<0.05, 0.01, and 0.001 are indicated by *, **, ***,
respectively. All calculations were performed using the 14.0 SPSS
software package (SPSS, Inc., Chicago, IL, USA).
Results
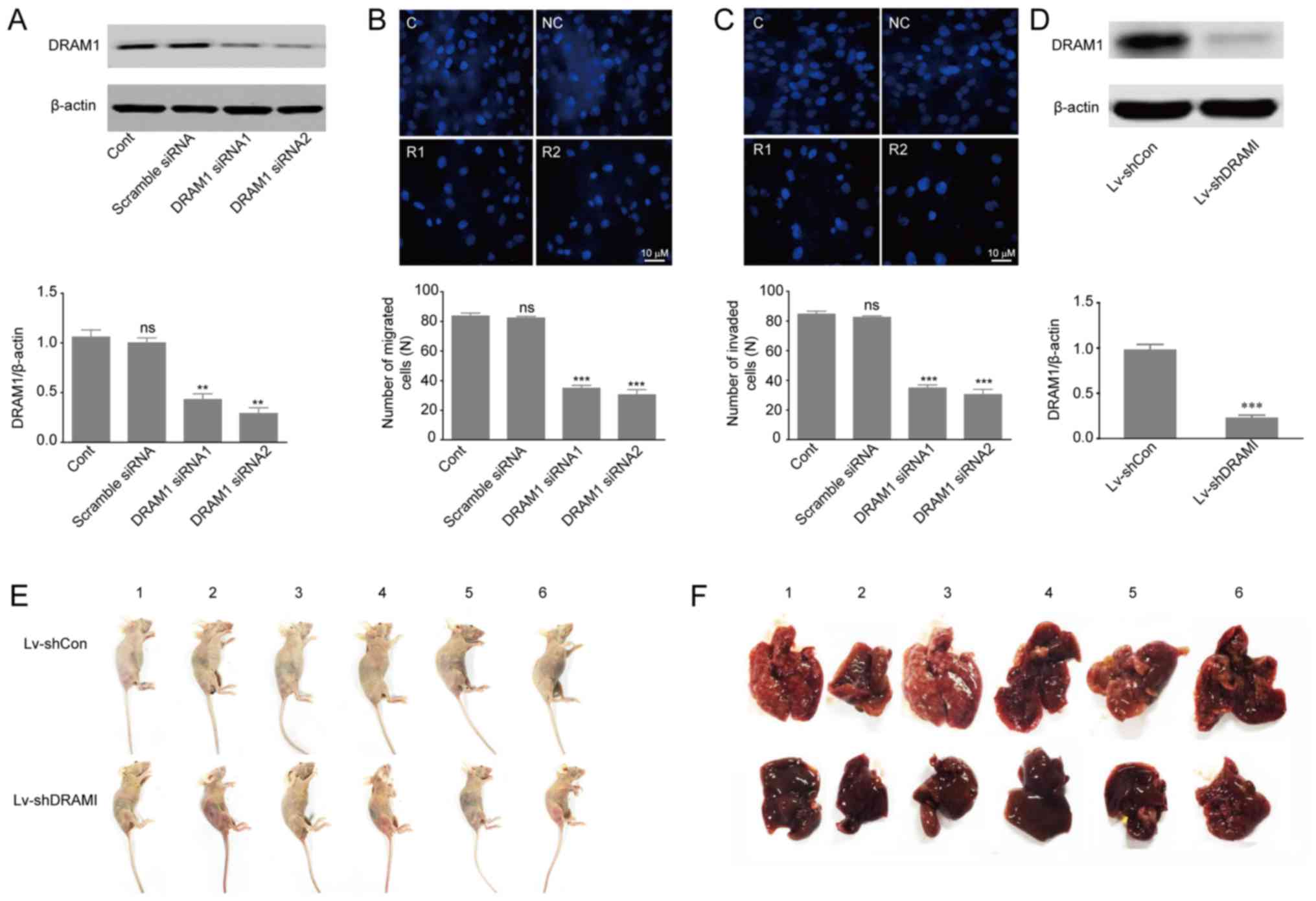
DRAM1 knockdown inhibits the invasion
and metastasis of HepG2 cells in vivo and in vitro
Previous studies reported that high expression of
DRAM1 indicated poor prognosis in GBM tumors, and that inhibition
of DRAM1 expression reduced invasion and metastasis of GSCs
(14,25). In order to further investigate the
effects of DRAM1 on invasion and metastasis of HepG2 cells, we
designed two different DRAM1 siRNA molecules (DRAM1 siRNA1, DRAM1
siRNA2) to knockdown DRAM1 expression. HepG2 cells were transfected
with scramble siRNA, DRAM1 siRNA1 or DRAM1 siRNA2 at the
concentration of 80 nM for 48 h. Western blot analysis of DRAM1
protein levels showed 57 and 71% of silence efficiency with DRAM1
siRNA1 and DRAM1 siRNA2, respectively (Fig. 1A). Transwell chambers for migratory
and invasive culture system were used to detect the migration and
invasion abilities of HepG2 cells after DRAM1 was knocked down. The
results showed that knockdown of DRAM1 significantly reduced the
number of migrated (Fig. 1B) and
invaded (Fig. 1C) cells, which
indicated that DRAM1 knockdown inhibited migration and invasion
abilities of HepG2 cells. Moreover, we transduced HepG2 cells with
vectors expressing shRNAs against DRAM1 or the scramble shRNA
(Fig. 1D) and grafted these cells
into nude mice by tail vein injection (1×107 cells per
nude mouse). 4 weeks later the DRAM1 knockdown cells exhibited
slower growth and lower metastasis compared to the control
shRNA-transfected cells (Fig. 1E and
F). Collectively, these results suggested that knockdown of
DRAM1 inhibited the migration and invasion abilities of HepG2 cells
both in vitro and in vivo.
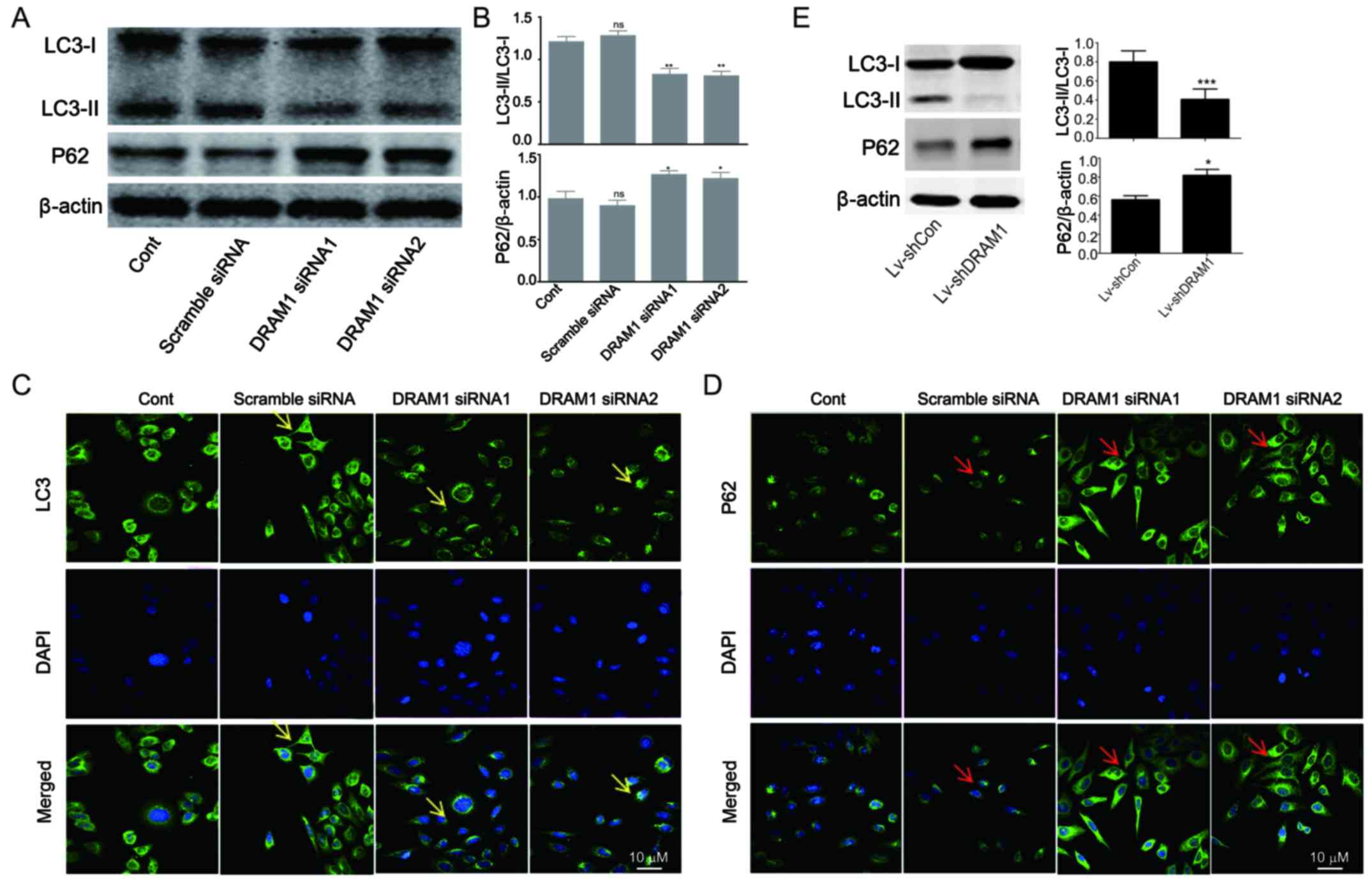
DRAM1 knockdown inhibits autophagy in
HepG2 cells
In order to investigate the mechanisms underlying
the inhibitory effects of DRAM1 knockdown on cell invasion and
metastasis, the levels of autophagy-related proteins were detected.
As shown in Fig. 2A and B, decreased
transformations of LC3I to LC3II as well as increased expressions
of p62 were observed in DRAM1 siRNA groups, indicating that
autophagy was inhibited by DRAM1 knockdown (Fig. 2A and B). We next performed
immunofluorescence to further detect the distribution of LC3 and
p62 in HepG2 cells. In consistent with Western blot analysis, the
results showed decreased distribution of LC3II and increased
expression of p62 in the cytoplasm in DRAM1 knockdown cells
(Fig. 2C and D), which suggested that
DRAM1 knockdown inhibited cell autophagy. Also, similar result was
observed in knockdown cells by shDRAM1 (Fig. 2E).
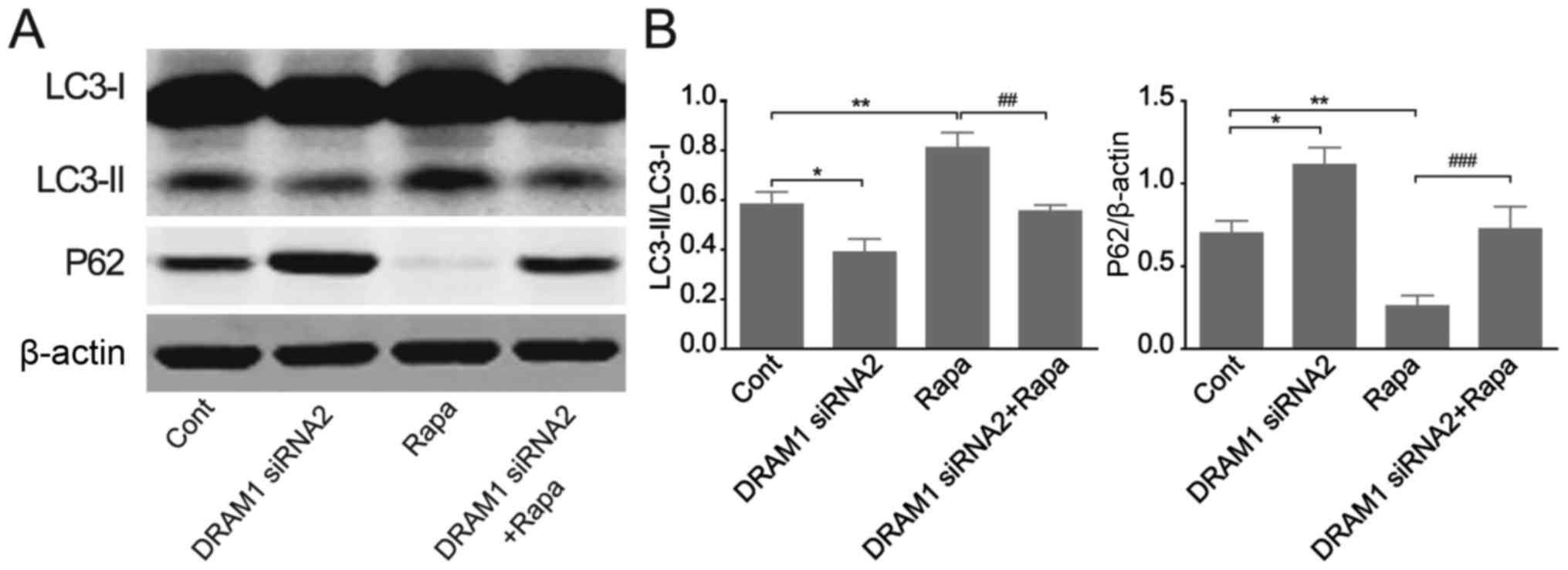
DRAM1 knockdown abrogates RAPA-induced
autophagy in HepG2 cells
We continued to verify the effects of DRAM1
knockdown on cell autophagy by applying RAPA, an autophagy inducer.
HepG2 cells were transfected with DRAM1 siRNAs for 48 h with or
without RAPA treatment and autophagy related protein levels were
detected by Western blots. The results showed that the autophagy
induced by RAPA stimulation was inhibited by DRAM1 knockdown. RAPA
treatment caused an increased transformation of LC3I to LC3II and a
reduced expression of p62 (Fig. 3A and
B), and the stimulatory effects of RAPA on cell autophagy were
obviously inhibited when DRAM1 was knocked down. These results
further verified the inhibitory potential of DRAM1 knockdown on
cell autophagy.
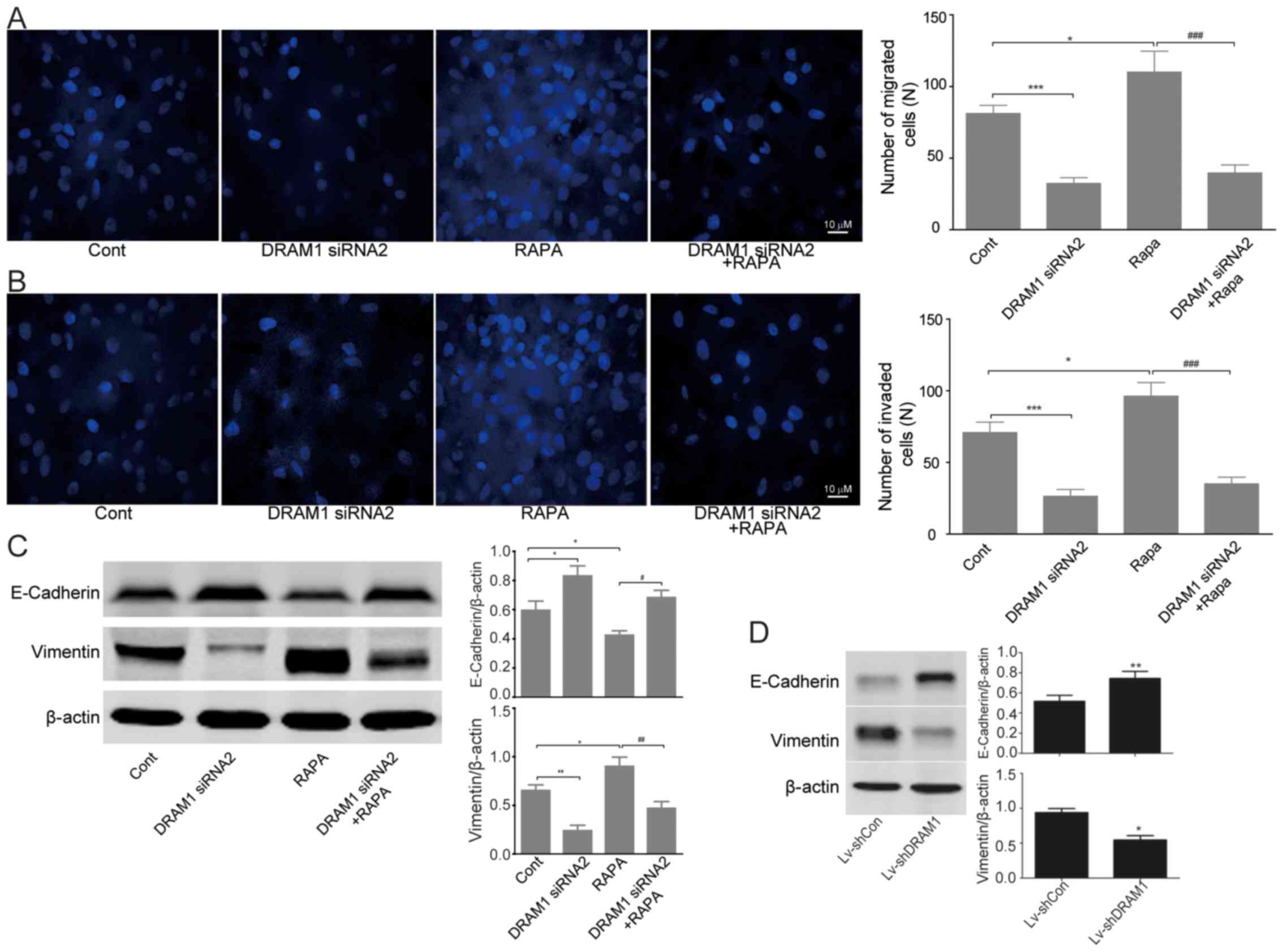
Invasion and migration of HepG2 cells
were inhibited through autophagy-EMT pathway when DRAM1 was knocked
down
We next set out to discover the mechanisms
underlying the inhibitory effects of DRAM1 knockdown on the cell
invasion and metastasis. Transwell chamber migratory and invasive
culture systems were adopted to detect the migratory and invasive
ability of HepG2 cells after DRAM1 was knocked down. As showed in
Fig. 4A and B, cell invasion and
metastasis were inhibited after DRAM1 knockdown. Moreover, the
stimulatory effects of RAPA on cell invasion and metastasis were
also obviously rescued by DRAM1 knockdown (Fig. 4A and B).
Since we have observed significant inhibition of
DRAM1 knockdown on the invasion and metastasis abilities of HepG2
cells both in vitro and in vivo, we next detected the
expression of EMT related proteins by western blots. The results
showed an increased expression of E-cadherin and a decreased
expression of vimentin in DRAM1 knockdown HepG2 cells, which
indicated an inhibitory effect of DRAM1 knockdown on EMT. On the
contrary, RAPA treatment caused a decreased expression of
E-cadherin and an increased expression of vimentin, both of which
were significantly reversed by the combined treatment of DRAM1
siRNA (Fig. 4C). Similar result was
obtained by shDRAM1 (Fig. 4D).
Collectively, these results suggested that the inhibitory effects
of DRAM1 knockdown on the invasion and migration abilities of HepG2
cells likely worked through the autophagy-EMT pathway.
Discussion
HepG2 cells were shown to be a HB cell line, which
did not affected the outcomes of the study. HB is the most common
liver malignant tumor diagnosed by the age of 4 years, accounting
for 80% of liver cancers in children under the age of 15 years,
among patients with localized HB, surgical resection is a common
treatment option (26,27). Liver transplantation is considered to
be the only curative therapy, however, the majority of patients
with advanced HB are not suitable for transplantation (1). Therefore, a more comprehensive
understanding of the regulatory mechanisms of HB invasion and
migration is beneficial for the survival improvement of HB
patients. Previous reports have shown that DRAM1 played an
important role in the migration and invasion of GSCs (14). Our previous results also revealed that
DRAM1 was highly expressed in intestinal cancer. However, little is
known about the precise contribution of DRAM1 in the migration and
invasion of HB cells. In this study, we found that DRAM1 knockdown
could inhibit the migration and invasion of HepG2 cells in the
setting of transwell assay, which was further confirmed by animal
experiments.
EMT is a natural process in which epithelial cells
obtain characteristics of mesenchymal cells, which is recognized as
an important procedure in cell invasion and metastasis. EMT happens
in normal cellular processes, however, it may also be exploited by
cancer cells, which triggers them to invade and form metastases at
distant sites. EMT was reported to motivate the expression of
mesenchymal markers in epithelial cells (28). The molecular regulators involved in
EMT include specific molecules which distinguish epithelial cells
from mesenchymal cells, and other constituents that can drive cells
towards the targeted sites. Cells undergoing EMT typically show an
increase in the expression of vimentin, fibronectin and integrin
αvβ6, as well as a decrease in the expression of E-cadherin and
cytokeratins. In our present study, with the application of
rapamycin, the expression of vimentin was significantly upregulated
while E-cadherin was downregulated in HepG2 cells. Meanwhile, the
concomitant knockdown of DRAM1 reversed the alterations of these
EMT markers. Our results suggested that DRAM1 knockdown regulated
EMT thus inhibiting the invasion and metastasis of HepG2 cells.
Growing evidence has suggested that autophagy is
closely related to human physiology and diseases including cancers.
Pervious researches revealed that DRAM1 was a lysosomal protein
associated with cell autophagy (13,29,30).
Therefore, we speculated that autophagy may be involved in the
inhibition of DRAM1 knockdown on the migration and invasion of HB
cells. The results in this study demonstrated that the elevation of
LC3-II and the decline of P62 stimulated by rapamycin treatment
were blocked by DRAM1 knockdown, which suggested that cell
autophagy was inhibited after DRAM1 knockdown. However, it is still
controversial on the function of autophagy in regulating cell
migration and invasion. Robin and colleagues showed that autophagy
inhibition significantly reduced the invasion of tumor cells
(31). Zhan et al reported
that autophagy induced by the activation of toll-like receptor 3 or
4 could enhance the production of various cytokines and thereby
facilitating the migration and invasion of lung cancer cells
(32). On the contrary, there were
also several researches suggested that autophagy could inhibit
tumor cell migration and invasion (22,33). In
this study, we found that DRAM1 knockdown significantly decreased
the levels of EMT markers and inhibited the migration and invasion
of HepG2 cells, which could be reversed by rapamycin treatment. It
needs to be clarified in future studies whether we could reverse
the inhibition of migration and invasion in DRAM1-silenced HepG2
cells by activating autophagy.
In summary, DRAM1 played an important role in the
regulation of HB migration and invasion. Our study demonstrated
that DRAM1 knockdown inhibited cell invasion and migration by
inhibiting the autophagy-EMT pathway, which could provide basic
knowledge for the development of new therapies for HB in clinical
practice.
Acknowledgements
Not applicable.
Funding
This work was supported by the Medicine and
Technology Project For Youth Changshu (CSWSQ201707) and the
National Natural Science Foundation of China (grant no.
81602613).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CC contributed to the idea conception, experimental
work and manuscript preparation. QYL, HKC, PFW, ZYF, XMM and HRW
contributed to the experimental work and manuscript preparation.
GQZ guided the idea conception, experimental work and manuscript
preparation. All authors reviewed the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sia D, Villanueva A, Friedman SL and
Llovet JM: Liver cancer cell of origin, molecular class, and
effects on patient prognosis. Gastroenterology. 152:745–761. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Avni FE, Massez A and Cassart M: Tumours
of the fetal body: A review. Pediatr Radiol. 39:1147–1157. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Catanzarite V, Hilfiker M, Daneshmand S
and Willert J: Prenatal diagnosis of fetal hepatoblastoma: Case
report and review of the literature. J Ultrasound Med.
27:1095–1098. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Apte U, Zeng G, Thompson MD, Muller P,
Micsenyi A, Cieply B, Kaestner KH and Monga SP: Beta-catenin is
critical for early postnatal liver growth. Am J Physiol
Gastrointest Liver Physiol. 292:G1578–G1585. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Monga SP: Role and regulation of β-catenin
signaling during physiological liver growth. Gene Expr. 16:51–62.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan X, Yuan Y, Zeng G, Apte U, Thompson
MD, Cieply B, Stolz DB, Michalopoulos GK, Kaestner KH and Monga SP:
Beta-catenin deletion in hepatoblasts disrupts hepatic
morphogenesis and survival during mouse development. Hepatology.
47:1667–1679. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tanimizu N and Miyajima A: Notch signaling
controls hepatoblast differentiation by altering the expression of
liver-enriched transcription factors. J Cell Sci. 117:3165–3174.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hartmann W, Küchler J, Koch A, Friedrichs
N, Waha A, Endl E, Czerwitzki J, Metzger D, Steiner S, Wurst P, et
al: Activation of phosphatidylinositol-3′-kinase/AKT signaling is
essential in hepatoblastoma survival. Clin Cancer Res.
15:4538–4545. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Testa JR and Tsichlis PN: AKT signaling in
normal and malignant cells. Oncogene. 24:7391–7393. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aden DP, Fogel A, Plotkin S, Damjanov I
and Knowles BB: Controlled synthesis of HBsAg in a differentiated
human liver carcinoma-derived cell line. Nature. 282:615–616. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
12
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR and Freshney RI: Check your cultures! A list of
cross-contaminated or misidentified cell lines. Int J Cancer.
127:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Crighton D, Wilkinson S, O'Prey J, Syed N,
Smith P, Harrison PR, Gasco M, Garrone O, Crook T and Ryan KM:
DRAM, a p53-induced modulator of autophagy, is critical for
apoptosis. Cell. 126:121–134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Galavotti S, Bartesaghi S, Faccenda D,
Shaked-Rabi M, Sanzone S, McEvoy A, Dinsdale D, Condorelli F,
Brandner S, Campanella M, et al: The autophagy-associated factors
DRAM1 and p62 regulate cell migration and invasion in glioblastoma
stem cells. Oncogene. 32:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang XD, Qi L, Wu JC and Qin ZH: DRAM1
regulates autophagy flux through lysosomes. PLoS One. 8:e632452013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Klymkowsky MW and Savagner P:
Epithelial-mesenchymal transition: A cancer researcher's conceptual
friend and foe. Am J Pathol. 174:1588–1593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boya P, Reggiori F and Codogno P: Emerging
regulation and functions of autophagy. Nat Cell Biol. 15:713–720.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kroemer G, Mariño G and Levine B:
Autophagy and the integrated stress response. Mol Cell. 40:280–293.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lv Q, Wang W, Xue J, Hua F, Mu R, Lin H,
Yan J, Lv X, Chen X and Hu ZW: DEDD interacts with PI3KC3 to
activate autophagy and attenuate epithelial-mesenchymal transition
in human breast cancer. Cancer Res. 72:3238–3250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qiang L, Zhao B, Ming M, Wang N, He TC,
Hwang S, Thorburn A and He YY: Regulation of cell proliferation and
migration by p62 through stabilization of Twist1. Proc Natl Acad
Sci USA. 111:9241–9246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Catalano M, D'Alessandro G, Lepore F,
Corazzari M, Caldarola S, Valacca C, Faienza F, Esposito V,
Limatola C, Cecconi F and Di Bartolomeo S: Autophagy induction
impairs migration and invasion by reversing EMT in glioblastoma
cells. Mol Oncol. 9:1612–1625. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li G, Li CX, Xia M, Ritter JK, Gehr TW,
Boini K and Li PL: Enhanced epithelial-to-mesenchymal transition
associated with lysosome dysfunction in podocytes: Role of
p62/sequestosome 1 as a signaling hub. Cell Physiol Biochem.
35:1773–1786. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gugnoni M, Sancisi V, Gandolfi G, Manzotti
G, Ragazzi M, Giordano D, Tamagnini I, Tigano M, Frasoldati A,
Piana S and Ciarrocchi A: Cadherin-6 promotes EMT and cancer
metastasis by restraining autophagy. Oncogene. 36:667–677. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Humbert M, Mueller C, Fey MF and Tschan
MP: Inhibition of damage-regulated autophagy modulator-1 (DRAM-1):
Impairs neutrophil differentiation of NB4 APL cells. Leuk Res.
36:1552–1556. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Herzog CE, Andrassy RJ and Eftekhari F:
Childhood cancers: Hepatoblastoma. Oncologist. 5:445–453. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Darbari A, Sabin KM, Shapiro CN and
Schwarz KB: Epidemiology of primary hepatic malignancies in U.S.
children. Hepatology. 38:560–566. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu J, Lamouille S and Derynck R:
TGF-beta-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Crighton D, Wilkinson S and Ryan KM: DRAM
links autophagy to p53 and programmed cell death. Autophagy.
3:72–74. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mah LY, O'Prey J, Baudot AD, Hoekstra A
and Ryan KM: DRAM-1 encodes multiple isoforms that regulate
autophagy. Autophagy. 8:18–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Macintosh RL, Timpson P, Thorburn J,
Anderson KI, Thorburn A and Ryan KM: Ryan, Inhibition of autophagy
impairs tumor cell invasion in an organotypic model. Cell Cycle.
11:2022–2029. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhan Z, Xie X, Cao H, Zhou X, Zhang XD,
Fan H and Liu Z: Autophagy facilitates TLR4- and TLR3-triggered
migration and invasion of lung cancer cells through the promotion
of TRAF6 ubiquitination. Autophagy. 10:257–268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu S, Xie F, Wang H, Liu Z, Liu X, Sun L
and Niu Z: Ubenimex inhibits cell proliferation, migration and
invasion in renal cell carcinoma: The effect is
autophagy-associated. Oncol Rep. 33:1372–1380. 2015. View Article : Google Scholar : PubMed/NCBI
|