Introduction
Pancreatic cancer is one of the most aggressive
human cancers and is a leading cause of cancer-associated
mortality. Surgical resection is the most effective treatment;
however, tumors have already advanced locally or metastasized to
distant organs at the time of diagnosis in most cases (1,2).
Gemcitabine is the current standard treatment for advanced or
resected pancreatic cancer (3).
However, as a large number of patients are resistant to gemcitabine
treatment, the drug provides only modest survival benefits. To
develop novel therapeutic strategies for pancreatic cancer, it is
necessary to gain further insight into the molecular mechanisms
supporting the malignant characteristics of pancreatic cancer.
ATPase family AAA domain containing protein 2
(ATAD2), which is also known as AAA+ nuclear co-regulator cancer
associated (ANCCA) is highly conserved in a wide range of species
and is associated with multiple biological processes, including
cell proliferation, migration and invasion (4). ATAD2 has an AAA+ domain in the central
region and a bromodomain in the C-terminal region (5). AAA+ domains mediate protein
multimerization and promote ATP hydrolysis to produce chemical
energy, which subsequently induces conformational changes of
substrate proteins to regulate their functions (6,7).
Bromodomains are small helical interacting modules that bind to
acetylated lysines in histones or other proteins (8). ATAD2 has been demonstrated to regulate
gene expression or chromatin modification in cooperation with
multiple transcription factors (9–13).
Accumulating studies have demonstrated that ATAD2 is associated
with the progression of numerous cancers. For example, high
expression of ATAD2 is a strong predictor of poor prognosis in
breast cancer (14,15). ATAD2 promotes proliferation and
survival of breast cancer cells by regulating the expression of
B-Myb, enhancer of zeste 2 polycomb repressive complex 2 subunit or
nuclear receptor coactivator 3 (16).
High expression of ATAD2 has also been reported in endometrial
carcinoma, hepatocellular carcinoma, cervical cancer and lung
cancer (17–28). These studies clearly demonstrate that
ATAD2 is critical for cancer progression and is a promising target
for cancer therapy. The present study examined whether ATAD2 was
involved in the progression of pancreatic cancer, and demonstrated
that ATAD2 is important for survival, invasion and drug resistance
of pancreatic cancer cells.
Materials and methods
Cells and antibodies
The pancreatic cancer cell lines, KP4, PK9,
MIAPaCa-2, PK8, RI151, PANC1 and KML1 were cultured in RPMI medium
(Wako, Osaka, Japan) with 10% fetal bovine serum (FBS) (Equitec,
Hendra, Australia) and antibiotics (penicillin, 50 IU/ml,
streptomycine, 50 µg/ml) at 37°C with 5% carbon dioxide
(CO2). The cell lines were obtained from RIKEN
BioResource Center (Tsukuba, Japan). ATAD2 (AMAB90541) and β-actin
(A5316) antibodies were obtained from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany), and cleaved poly (ADP ribose) polymerase
antibody (9541) was obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Western blotting
Cells were lysed with Laemmli sample buffer (20%
glycerol, 135 mM Tris-HCl, pH 6.8, 4% SDS, 10% 2-Mercaptoethanol,
and 0.003% bromophenol blue) and boiled for 5 min. The protein
concentrations of the lysates were measured using the RC-DC Protein
Assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Equal
protein quantities (30 µg) were separated by SDS-PAGE (10%) and
transferred to PVDF membranes (Merck KGaA). The membranes were
blocked with 0.5% non-fat skim milk at 37°C for 30 min, incubated
with each primary antibody (1,000 dilution) at room temperature for
1 h, washed with TBS-T buffer (10 mM Tris-HCl, pH 7.4, 150 mM NaCl,
and 0.05% Tween-20) and then incubated with HRP-conjugated
anti-mouse IgG (31430) or anti-rabbit IgG (31460) secondary
antibodies (Thermo Fisher Scientific, Inc., Waltham, MA, USA) in
3,000 dilution at room temperature for 1 h. The proteins were
visualized using enhanced chemiluminescence (GE Healthcare Life
Sciences, Uppsala, Sweden).
Small interfering RNA (siRNA)
transfection
siRNAs were obtained from Sigma-Aldrich; Merck KGaA.
The sequences of siRNAs for ATAD2 were as follows: ATAD2-1,
5′-CCAGCUGUCAUUCAUGCUUTT-3′ and ATAD2-2,
5′-GCCUACACCCUCACUUGUUTT-3′. The sequence of the control siRNA
targeting luciferase was 5′-CUUACGCUGAGUACUUCGATT-3′. Cells were
transfected with 20 nM siRNA using Lipofectamine RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol.
Proliferation assay
Cells (1,000 cells/well) were cultured in 96-well
plates 24 h following transfection with siRNAs, and the number of
viable cells at 24, 48 and 72 h were evaluated using a Cell
Counting Kit-8 assay, according to manufacturer's protocol (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan). To evaluate the
proliferation of siRNA-transfected cells in the presence of
gemcitabine, siRNA-transfected cells (1,000 cell/well) were
cultured in triplicate in 96-well plates with various
concentrations of gemcitabine (0.05, 0.1, 0.2, 0.4, 0.6, 0.8 and
1.0 µM) at 37°C for 48 h. Gemcitabine was obtained from Wako
(Osaka, Japan). The number of viable cells was evaluated using Cell
Counting Kit-8 assays 3 days following this.
TUNEL assay
Cells were fixed with 4% paraformaldehyde and
subjected to TUNEL assays using the In Situ Cell Death
Detection kit and fluorescein (Roche Diagnostics, Basel,
Switzerland) 72 h following transfection with siRNAs, according to
the manufacturer's protocol. Cells in five randomly selected fields
were evaluated using fluorescent microscope (BX60; Olympus, Tokyo,
Japan) at ×100 magnification, and three independent experiments
were performed.
Migration and invasion assay
Cell migration and invasion were measured using
Boyden chambers (8 µm pore size and 6.5 mm membrane diameter;
Corning Incorporated, Corning, NY, USA). To evaluate cell
migration, 72 h following transfection with siRNAs, cells
(4×104) were placed on the upper surface of a filter
coated with fibronectin and then allowed to migrate to the bottom
surface. Upper chamber and lower chamber were filled with RPMI
medium without serum. Cells were fixed with 70% ethanol and stained
with 0.5% crystal violet 6 h subsequent to this. Cells that
migrated to the lower surface of the filters were quantified in
five randomly selected fields using a microscope (BX60; Olympus) at
×40 magnification, three independent experiments were performed. To
evaluate cell invasion, cells were placed on the upper surface of a
filter coated with Matrigel (BD Bioscience, San Jose, CA, USA) 72 h
following transfection with siRNAs. The upper chamber was filled
with RPMI and the lower chamber was filled with RPMI supplemented
with 10% FBS. Cells were fixed with 70 % ethanol and stained 24 h
subsequent to this to count cells that had invaded to the lower
surface of the filter.
Colony formation assay
Cells were transfected with siRNAs, and 24 h
subsequent to this, cells (1×104) were mixed with 0.36%
agar in RPMI medium supplemented with 10% FBS, and overlaid onto a
0.72% agarose layer in 6-well plates. Following incubation at 37°C
for 2 weeks, colonies in five randomly selected fields were counted
using a microscope (BX60; Olympus). Three independent experiments
were performed.
Statistical analysis
The data were expressed as the mean ± standard
deviation. Comparisons between the groups were performed using
unpaired Student's t-tests using Excel software (Microsoft
Corporation, Redmond, WA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
ATAD2 knockdown promotes
apoptosis
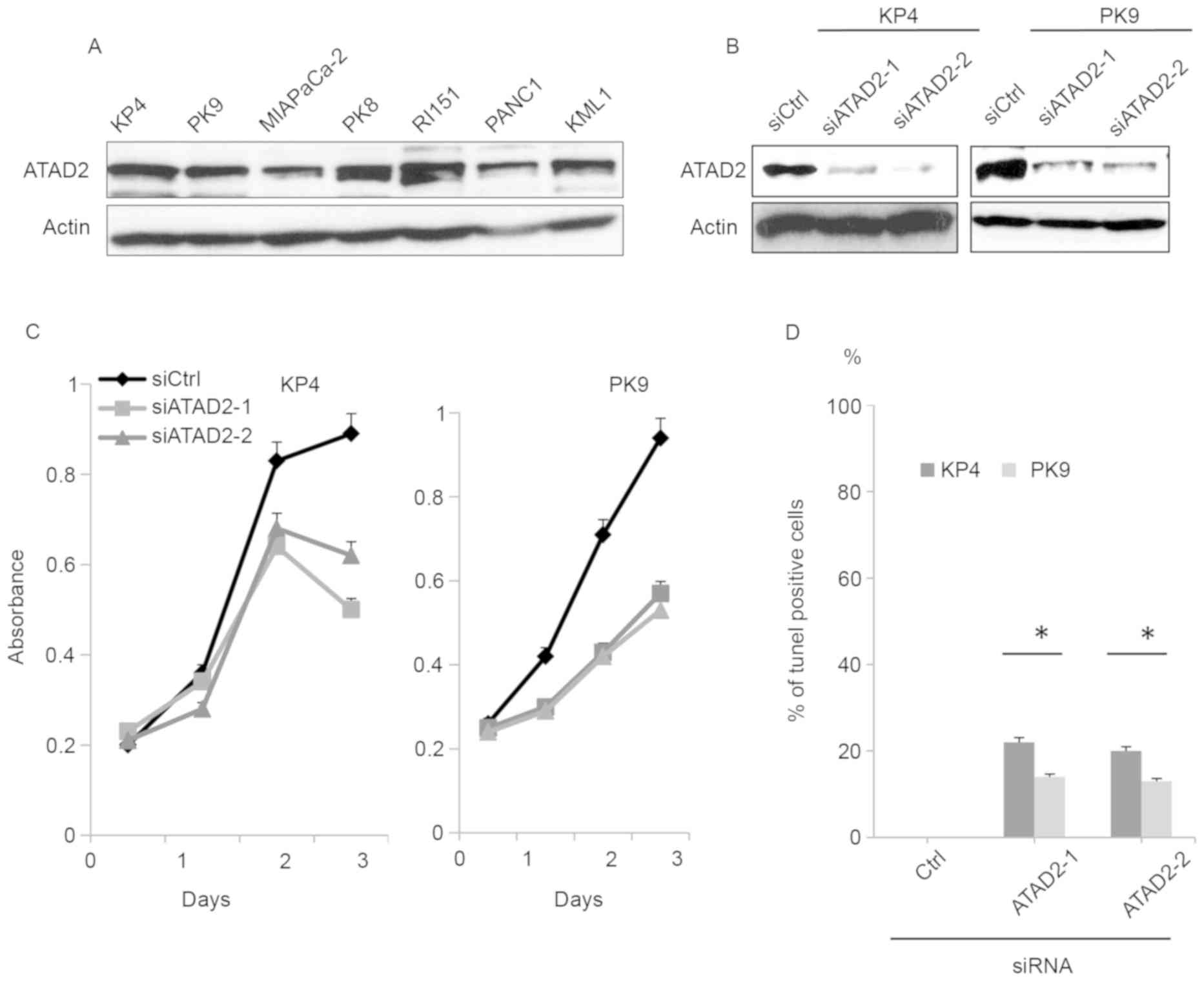
To determine the involvement of ATAD2 in pancreatic
cancer progression, the expression of ATAD2 was examined in KP4,
PK9, MIAPaCa-2, PK8, RI151, PANC1 and KML1 pancreatic cancer cell
lines. ATAD2 was expressed at similar levels in multiple pancreatic
cancer cell lines (Fig. 1A). The KP4
and PK9 cell lines were selected for further analysis. Transfection
of two different siRNAs sufficiently reduced the level of
expression of ATAD2 in KP4 and PK9 cell lines (Fig. 1B). Depletion of ATAD2 significantly
reduced the proliferation of either KP4 or PK9 cells (Fig. 1C). To determine whether reduced
proliferation resulted from an increase in apoptotic cells, cells
transfected with siRNAs were stained for DNA breaks using the TUNEL
assay. ATAD2 knockdown promoted apoptosis of both cell lines
(Fig. 1D).
ATAD2 knockdown suppresses cell
migration and invasion
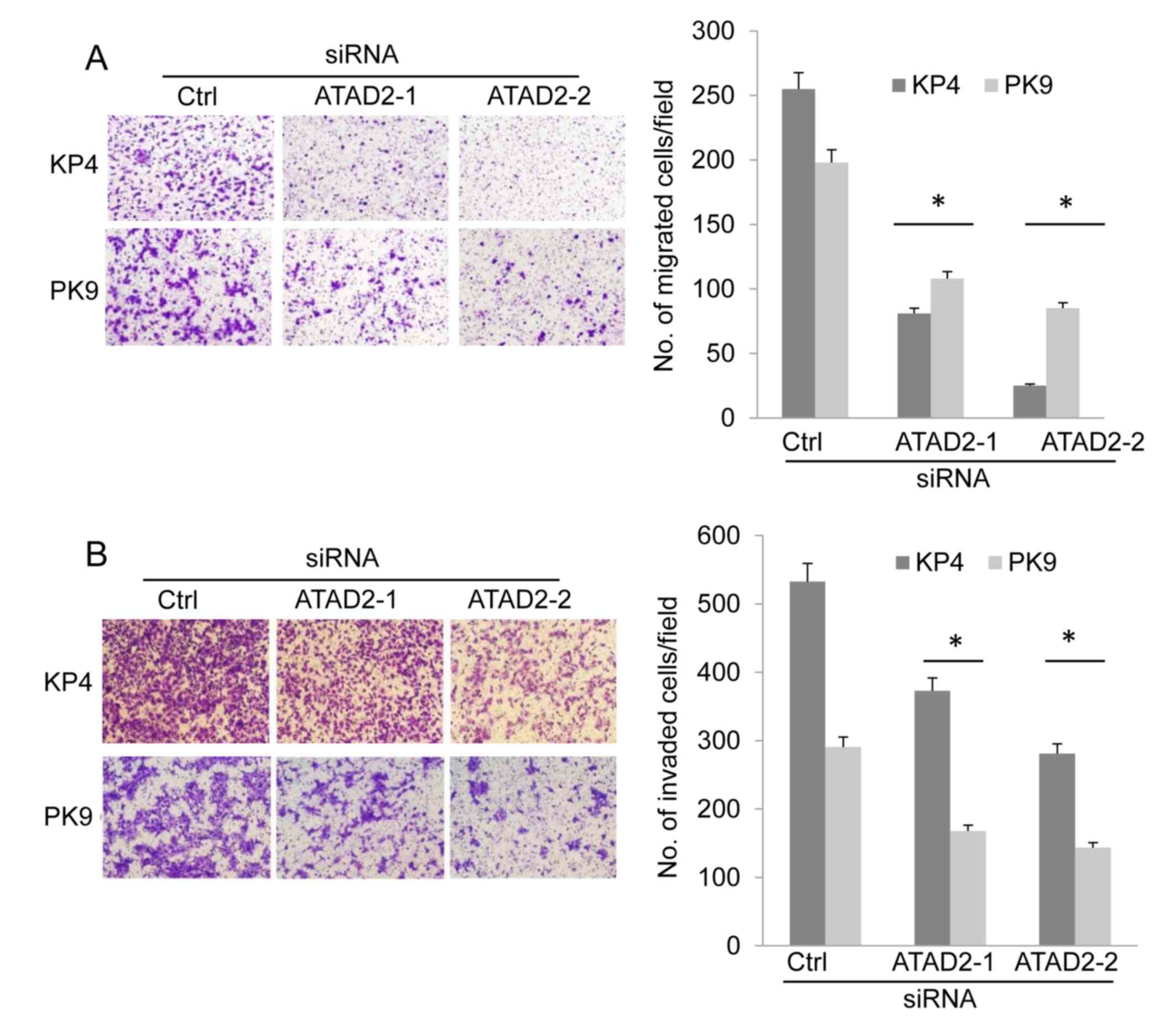
Cell migration and invasion of ATAD2-depleted cells
was examined using a modified Boyden chamber. KP4 and PK9 cells
were transfected with siRNAs and, 72 h following this, cells were
suspended and placed in the upper chambers of the filters. The
cells were allowed to migrate to the bottom surface of the filter,
which was coated with fibronectin. The migrated cells were counted
6 h subsequent to this to evaluate cell migration. The migration of
KP4 and PK9 cells was decreased following ATAD2 knockdown compared
with cells transfected with control siRNA (Fig. 2A). To determine cell invasive ability,
Matrigel-coated Boyden chambers were used. ATAD2 depletion
significantly suppressed the invasion of KP4 and PK9 cells compared
with cells transfected with control siRNA (Fig. 2B). These results demonstrated that
ATAD2 was associated with the migration and invasion of pancreatic
cancer cells.
ATAD2 depletion suppresses
anchorage-independent growth
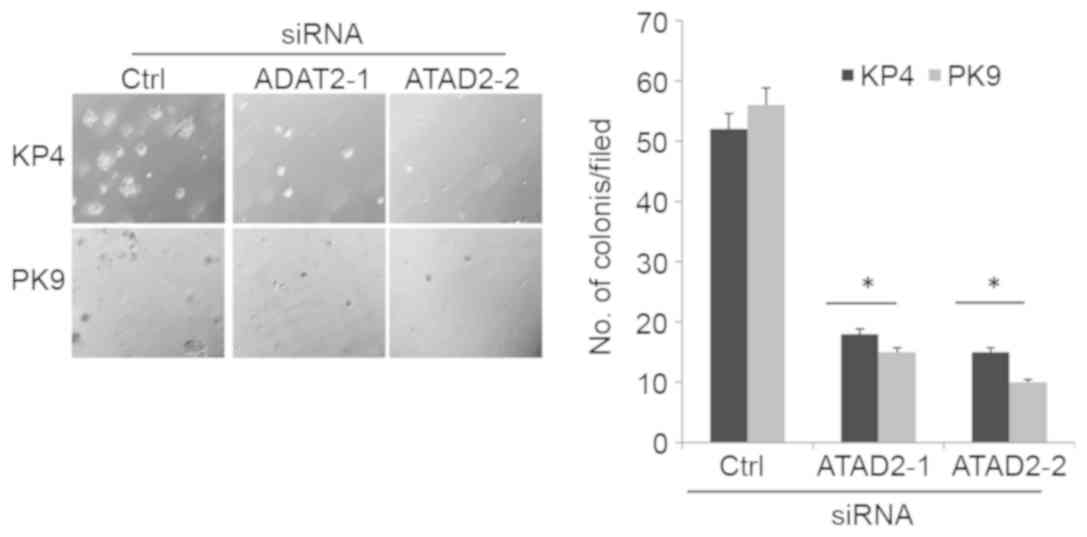
One of the malignant characteristics of cancer cells
is the ability to grow in the absence of cell adhesion to the
extracellular matrix, known as anchorage-independent growth. To
determine the ability of cells to grow in the absence of cell
adhesion, siRNA-transfected cells were cultured in soft agar for
two weeks, and then the formation of colonies was examined. ATAD2
depletion significantly suppressed anchorage-independent growth of
KP4 and PK9 cells compared with cells transfected with control
siRNA (Fig. 3).
ATAD2 depletion sensitizes cells to
gemcitabine
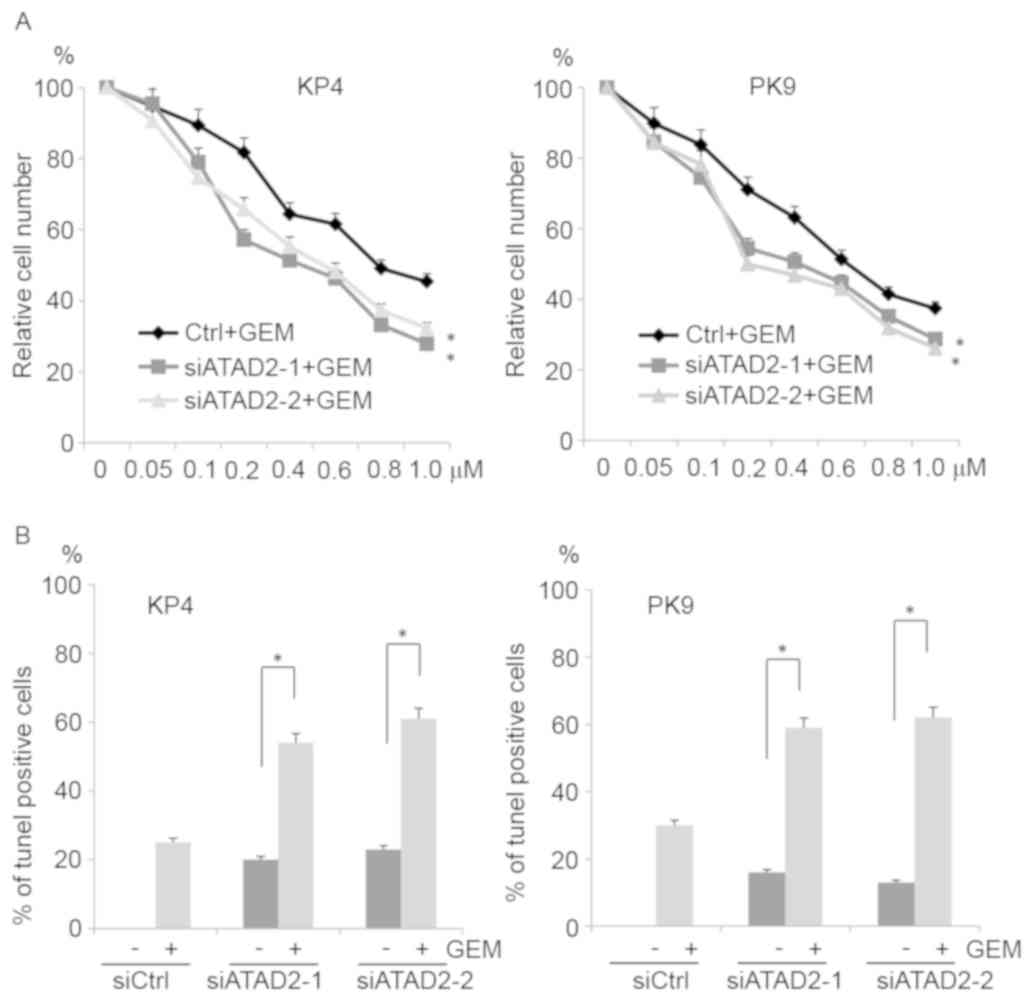
To determine whether ATAD2 knockdown sensitized
pancreatic cancer cells to gemcitabine, which is a nucleoside
analog frequently used for pancreatic cancer treatment,
siRNA-transfected cells were cultured in the presence of 0–1.0 µM
gemcitabine and cell growth was assessed. ATAD2 siRNA-transfected
cells were more sensitive to gemcitabine than control
siRNA-transfected cells (Fig. 4A). To
further confirm this result, TUNEL assays were performed.
siRNA-transfected cells were cultured in the presence of 1.0 µM
gemcitabine, and the apoptotic cells were examined. More apoptotic
cells were observed among the ATAD2-depleted cells than the control
cells (Fig. 4B). These results
indicated that ATAD2 knockdown sensitized pancreatic cancer cells
to gemcitabine.
Discussion
The present study provided evidence that ATAD2 is
associated with the progression of pancreatic cancer. Western
blotting analysis demonstrated that ATAD2 was expressed in all the
pancreatic cancer cell lines that were examined. To determine the
function of ATAD2 in the malignant characteristics of pancreatic
cancer cells, two siRNAs that targeted different regions of the
ATAD2 mRNA were used, to exclude the possibility of off-target
effects. These siRNAs sufficiently decreased ATAD2 expression.
Depletion of ATAD2 by these siRNAs significantly reduced invasion
and migration in KP4 and PK9 cells. In addition, ATAD2 knockdown
suppressed anchorage-independent growth of pancreatic cancer cells.
These results indicated that ATAD2 is associated with the
malignancy of pancreatic cancer.
ATAD2 suppression has previously been indicated to
facilitate P53 and P38-dependent apoptosis of hepatocellular
carcinoma cells (27). ATAD2 is also
essential for the survival and proliferation of breast cancer
cells. ATAD2 was required for the expression of members of the
kinesin families, including kinesin family member (KIF) 4A, KIF20A,
and KIF23, that are essential for the progression of mitosis
(15). As a result, depletion of
ATAD2 induced apoptosis of drug-sensitive and drug-resistant cancer
cells (15). ATAD2 depletion was
demonstrated to significantly induce apoptosis of pancreatic cancer
cells in the present study. These results indicate that ATAD2 is
essential for the survival of multiple cancer cells.
ATAD2 depletion increased the sensitivity of
pancreatic cancer cells to gemcitabine. Although gemcitabine is
often used for pancreatic cancer treatment, its effect is far from
satisfactory. Accumulating studies have demonstrated that chemical
inhibitors for AAA+ proteins are useful for cancer treatment
(29–31). p97, a member of the AAA+ family of
proteins, is essential for the regulation of protein homeostasis.
Inhibition of p97 by small chemicals induced unfolded protein
responses and promoted apoptosis in multiple cancers (32). Thus, chemical inhibitors that disrupt
the enzymatic activity of ATAD2 may be useful for the treatment of
pancreatic cancer in combination with gemcitabine. In addition to
the AAA+ domain, ATAD2 has a conserved domain called the
bromodomain, which is essential for the binding of ATAD2 to
acetylated histone. Previous studies have reported that certain
chemicals specifically inhibit the bromodomain of ATAD2, further
demonstrating that ATAD2 may be a promising target for the
development of chemical inhibitors (33,34). The
results of the present study and other studies indicate that
combination therapy of gemcitabine and ATAD2 inhibitors may be a
potential therapeutic strategy for the treatment of pancreatic
cancers.
Acknowledgements
The authors would like to thank the members of the
Division of Cancer Biology, Nagoya University Graduate School of
Medicine (Nagoya, Japan) for their helpful discussions and
technical assistance. The present study was funded by a grant from
the Naito Foundation.
References
|
1
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berlin J and Benson AB III: Chemotherapy:
Gemcitabine remains the standard of care for pancreatic cancer. Nat
Rev Clin Oncol. 7:135–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boussouar F, Jamshidikia M, Morozumi Y,
Rousseaux S and Khochbin S: Malignant genome reprogramming by
ATAD2. Biochim Biophys Acta. 1829:1010–1014. 2103. View Article : Google Scholar
|
|
5
|
Cattaneo M, Morozumi Y, Perazza D,
Boussouar F, Jamshidikia M, Rousseaux S, Verdel A and Khochbin S:
Lessons from yeast on emerging roles of the ATAD2 protein family in
gene regulation and genome organization. Mol Cells. 37:851–856.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ogura T and Wilkinson AJ: AAA+ superfamily
ATPases: Common structure-diverse function. Genes Cells. 6:575–597.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hanson PI and Whiteheart SW: AAA+
proteins: Have engine, will work. Nat Rev Mol Cell Biol. 6:519–529.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ntranos A and Casaccia P: Bromodomains:
Translating the words of lysine acetylation into myelin injury and
repair. Neurosci Lett. 625:4–10. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zou JX, Guo L, Revenko AS, Tepper CG, Gemo
AT, Kung HJ and Chen HW: Androgen-induced coactivator ANCCA
mediates specific androgen receptor signaling in prostate cancer.
Cancer Res. 69:3339–3346. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ciró M, Prosperini E, Quarto M, Grazini U,
Walfridsson J, McBlane F, Nucifero P, Pacchiana G, Capra M,
Christensen J and Helin K: ATAD2 is a novel cofactor for MYC,
overexpressed and amplified in aggressive tumors. Cancer Res.
69:8491–8498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hsia EY, Kalashnikova EV, Revenko AS, Zou
JX, Borowsky AD and Chen HW: Deregulated E2F and the AAA+
coregulator ANCCA drive proto-oncogene ACTR/AIB1 overexpression in
breast cancer. Mol Cancer Res. 8:183–193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Revenko AS, Kalashnikova EV, Gemo AT, Zou
JX and Chen HW: Chromatin loading of E2F-MLL complex by
cancer-associated coregulator ANCCA via reading a specific histone
mark. Mol Cell Biol. 30:5260–5272. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duan Z, Zou JX, Yang P, Wang Y, Borowsky
AD, Gao AC and Chen HW: Developmental and androgenic regulation of
chromatin regulators EZH2 and ANCCA/ATAD2 in the prostate Via MLL
histone methylase complex. Prostate. 73:455–466. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Caron C, Lestrat C, Marsal S, Escoffier E,
Curtet S, Virolle V, Barbry P, Debernardi A, Brambilla C, Brambilla
E, et al: Functional characterization of ATAD2 as a new
cancer/testis factor and a predictor of poor prognosis in breast
and lung cancers. Oncogene. 29:5171–5181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zou JX, Duan Z, Wang J, Sokolov A, Xu J,
Chen CZ, Li JJ and Chen HW: Kinesin family deregulation coordinated
by bromodomain protein ANCCA and histone methyltransferase MLL for
breast cancer cell growth, survival, and tamoxifen resistance. Mol
Cancer Res. 12:539–549. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kalashnikova EV, Revenko AS, Gemo AT,
Andrews NP, Tepper CG, Zou JX, Cardiff RD, Borowsky AD and Chen HW:
ANCCA/ATAD2 overexpression identifies breast cancer patients with
poor prognosis, acting to drive proliferation and survival of
triple-negative cells through control of B-Myb and EZH2. Cancer
Res. 70:9402–9412. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fouret R, Laffaire J, Hofman P,
Beau-Faller M, Mazieres J, Validire P, Girard P, Camilleri-Bröet S,
Vaylet F, Leroy-Ladurie F, et al: A comparative and integrative
approach identifies ATPase family, AAA domain containing 2 as a
likely driver of cell proliferation in lung adenocarcinoma. Clin
Cancer Res. 18:5606–5616. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Raeder MB, Birkeland E, Trovik J, Krakstad
C, Shehata S, Schumacher S, Zack TI, Krohn A, Werner HM, Moody SE,
et al: Integrated genomic analysis of the 8q24 amplification in
endometrial cancers identifies ATAD2 as essential to MYC-dependent
cancers. PLoS One. 8:e548732013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Sun Y, Li Y, Fang Z, Wang R, Pan
Y, Hu H, Luo X, Ye T, Li H, et al: ANCCA protein expression is a
novel independent poor prognostic marker in surgically resected
lung adenocarcinoma. Ann Surg Oncol. 20 Suppl 3:S577–S582. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu G, Liu H, He H, Wang Y, Lu X, Yu Y, Xia
S, Meng X and Liu Y: miR-372 down-regulates the oncogene ATAD2 to
influence hepatocellular carcinoma proliferation and metastasis.
BMC Cancer. 14:1072014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wan WN, Zhang YX, Wang XM, Liu YJ, Zhang
YQ, Que YH and Zhao WJ: ATAD2 is highly expressed in ovarian
carcinomas and indicates poor prognosis. Asian Pac J Cancer Prev.
15:2777–2783. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu G, Lu X, Wang Y, He H, Meng X, Xia S,
Zhen K and Liu Y: Epigenetic high regulation of ATAD2 regulates the
Hh pathway in human hepatocellular carcinoma. Int J Oncol.
45:351–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hwang HW, Ha SY, Bang H and Park CK: ATAD2
as a Poor prognostic marker for hepatocellular carcinoma after
curative resection. Cancer Res Treat. 47:853–861. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zheng L, Li T, Zhang Y, Guo Y, Yao J, Dou
L and Guo K: Oncogene ATAD2 promotes cell proliferation, invasion
and migration in cervical cancer. Oncol Rep. 33:2337–2344. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shang P, Meng F, Liu Y and Chen X:
Overexpression of ANCCA/ATAD2 in endometrial carcinoma and its
correlation with tumor progression and poor prognosis. Tumour Biol.
36:4479–4485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krakstad C, Tangen IL, Hoivik EA, Halle
MK, Berg A, Werner HM, Ræder MB, Kusonmano K, Zou JX, Øyan AM, et
al: ATAD2 overexpression links to enrichment of B-MYB-translational
signatures and development of aggressive endometrial carcinoma.
Oncotarget. 6:28440–28452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu WJ, Chua MS and So SK: Suppression of
ATAD2 inhibits hepatocellular carcinoma progression through
activation of p53- and p38-mediated apoptotic signaling.
Oncotarget. 6:417222015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang M, Zhang C, Du W, Yang X and Chen Z:
ATAD2 is overexpressed in gastric cancer and serves as an
independent poor prognostic biomarker. Clin Transl Oncol.
18:776–781. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chou TF, Brown SJ, Minond D, Nordin BE, Li
K, Jones AC, Chase P, Porubsky PR, Stoltz BM, Schoenen FJ, et al:
Reversible inhibitor of p97, DBeQ, impairs both ubiquitin-dependent
and autophagic protein clearance pathways. Proc Natl Acad Sci USA.
108:4834–4849. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chou TF, Li K, Frankowski KJ, Schoenen FJ
and Deshaies RJ: Structure-activity relationship study reveals
ML240 and ML241 as potent and selective inhibitors of p97 ATPase.
Chem Med Chem. 8:297–312. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Magnaghi P, D'Alessio R, Valsasina B,
Avanzi N, Rizzi S, Asa D, Gasparri F, Cozzi L, Cucchi U, Orrenius
C, et al: Covalent and allosteric inhibitors of the ATPase
VCP/p97induce cancer cell death. Nat Chem Biol. 9:548–556. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Anderson DJ, Le Moigne R, Djakovic S,
Kumar B, Rice J, Wong S, Wang J, Yao B, Valle E, Kiss von Soly S,
et al: Targeting the AAA ATPase p97 as an approach to treat cancer
through disruption of protein homeostasis. Cancer Cell. 28:653–665.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Demont EH, Chung CW, Furze RC, Grandi P,
Michon AM, Wellaway C, Barrett N, Bridges AM, Craggs PD, Diallo H,
et al: Fragment-Based Discovery of Low-Micromolar ATAD2Bromodomain
Inhibitors. J Med Chem. 58:5649–5673. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bamborough P, Chung CW, Furze RC, Grandi
P, Michon AM, Sheppard RJ, Barnett H, Diallo H, Dixon DP, Douault
C, et al: Structure-based optimization of naphthyridones into
PotentATAD2 bromodomain inhibitors. J Med Chem. 58:6151–6178. 2015.
View Article : Google Scholar : PubMed/NCBI
|