Introduction
A recent study estimated that >65,000 male and
female individuals would likely be diagnosed with colorectal cancer
(CRC) in the USA in 2014 (1). With
the number of mortalities ranging between 24,000 and 26,000
individuals per year in the USA, CRC is emerging as one of the
three leading causes of adult cancer. Various cellular factors have
recently emerged as important elements in maintaining the survival
and proliferation of CRC tumor cells, including signal transducer
and activator of transcription 3 (STAT3) (2). The expression of STAT3 and
phosphorylated STAT3 (p-STAT3) has been demonstrated to be
significantly higher in CRC tissues compared with healthy
intestinal mucosa (3). As STAT3 is
activated by numerous growth factors and cytokines, including
interleukin-6 (IL-6), the local cytokine environment may have a
significant role in the malignancy of CRC (4).
The binding of IL-6 to the IL-6 receptor (IL-6R)
initiates an intracellular signaling cascade that activates STAT3
and enhances the localized inflammatory environment, contributing
toward c ancer progression (5,6). This
association was highlighted by a study that measured the cytokine
levels in the sera of CRC patients and identified a direct
correlation between IL-6 expression levels and CRC progression
(7). Furthermore, activation of the
intracellular Janus kinase (JAK)/STAT3 signaling pathway by IL-6
results in the expression of various genes involved in cancer
growth and development (8). The
phosphorylation of STAT3 in the cytoplasm induces its
homodimerization, nuclear translocation and DNA binding (9). p-STAT3 acts as a transcriptional
activator of numerous genes, including cyclin D1 and B-cell
lymphoma-1 (Bcl-1), with its anti-apoptotic effects significantly
contributing to cell proliferation, and tumorigenesis (10,11).
Thus, the IL-6/STAT3 pathway is an emerging therapeutic target for
CRC.
Standard treatment strategies for CRC include a
combination of radiotherapy and chemotherapy, however, the
prognosis and survival rates of patients with advanced CRC is poor.
The common chemotherapeutic regimens used to treat CRC include
5-fluorouracil (5-FU)/leucovorin, capecitabine, irinotecan,
oxaliplatin, bevacizumab and cetuximab (12). In addition to these compounds,
various traditional Chinese medicines (TCMs) are currently being
evaluated as effective alternatives to the standard
chemotherapeutic arsenal. However, the precise mechanism of action
of TCMs, as well as the specific pathways that lead to their
tumor-suppressive activities, remain unclear.
Hedyotis diffusa Willd. [HDW; also known as
Oldenlandia diffusa (Willd.)] of the Rubiaceae family, is a
traditional Chinese herbal medicine that is reported to exhibit a
range of pharmacological roles, including anticancer,
anti-inflammatory, anti-oxidative, neuroprotective and
hepatoprotective activities (13,14).
Furthermore, numerous prescriptions of HDW have been demonstrated
to provide therapeutic efficacy (15). Our previous studies demonstrated
that ethanol extracts obtained from HDW (EEHDW) can induce
apoptosis via a mitochondria-dependent pathway in human colon
carcinoma HT-29 cells. In addition, treatment with EEHDW appeared
to inhibit CRC growth in vivo via the inhibition of the
STAT3 signaling pathway and suppress tumor angiogenesis via the
hedgehog signaling pathway (16–20).
Although our previous studies indicated that the activity of EEHDW
disrupted the STAT3 pathway, the anticancer efficacy of EEHDW
during cytokine-mediated STAT3 activation (such as by IL-6) was
largely unclear. Thus, to elucidate the mechanism of the
tumoricidal activity of EEHDW, the present study investigated its
effects on the IL-6-mediated activation of HT-29 cells in
vitro. Specifically, cell proliferation and apoptosis, the
phosphorylation levels and transcriptional activity of STAT3, and
the expression of various target genes of the IL-6/STAT3 signaling
pathway were examined to determine the efficacy of EEHDW during
cytokine-mediated STAT3 activation.
Materials and methods
Materials and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), penicillin-streptomycin, trypsin-EDTA, TRIzol
reagent, and caspase-9 and -3 activation kits were purchased from
Invitrogen Life Technology, Inc. (Carlsbad, CA, USA). Bcl-2,
Bcl-2-associated X protein (Bax), cyclin D1 and cyclin-dependent
kinase 4 (CDK4) monoclonal antibodies, as well as horseradish
peroxidase (HRP)-conjugated monoclonal secondary antibodies, were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
SuperScript II reverse transcriptase was obtained from Promega
Corporation (Madison, WI, USA), the DAPI staining kit was obtained
from Nanjing KeyGen Biotech Co., Ltd., (Nanjing, China) and the
bicinchoninic acid (BCA) protein assay kit was purchased from
Tiangen Biotech (Beijing) Co., Ltd. (Beijing, China). All the other
chemicals, unless otherwise stated, were obtained from
Sigma-Aldrich (St. Louis, MO, USA).
EEHDW preparation
The HDW plant material was purchased from a
commercial supplier (Guo Yi Tang Chinese Herbal Medicine Store,
Fujian, China) and the EEHDW was obtained as previously described
(20). Stock solutions of EEHDW
were prepared by dissolving the EEHDW powder in 40% dimethyl
sulfoxide (DMSO) to a final concentration of 400 mg/ml, and stock
solutions were stored at −20°C. The working concentrations of EEHDW
were made by diluting the stock solution in the culture medium to a
final concentration of <0.5% DMSO in the medium.
Cell culture
Human colon carcinoma HT-29 cells were purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). Cells were grown as adherent monolayers in DMEM culture
media containing 10% (v/v) FBS, 100 U/ml penicillin and 100 μg/ml
streptomycin at 37°C in a humidified incubator with an atmosphere
of 5% CO2.
EEHDW and IL-6 treatment
The HT-29 cells were cultured with DMEM medium
containing 10% FBS and 1% penicillin/streptomycin. When the cells
reached ~50% confluency, the complete medium was removed and
FBS-free medium was added prior to overnight incubation. The cells
were pre-treated with 1, 3 or 5 mg/ml EEHDW in complete DMEM medium
for 1 h, followed by stimulation with 10 ng/ml IL-6 for 15 min or
24 h.
Evaluation of cell viability using an MTT
assay
Cell viability was assessed by performing an MTT
colorimetric assay. The cells were harvested and resuspended at a
final concentration of 1×105 cells/ml, then seeded into
96-well tissue culture plates at a concentration of 100 μl/well.
Subsequent to incubating for 24 h at 37°C, the cells were treated
with 1, 3 or 5 mg/ml EEHDW and/or 10 ng/ml IL-6 for an additional
24 h. Next, 100 μl MTT (0.5 mg/ml) was added to each well, the
plates were incubated at 37°C for 4 h and 100 μl DMSO was added to
dissolve the purple formazan crystals. Finally, the absorbance was
read at a wavelength of 570 nm using an ELISA reader (Model EXL800;
BioTek Instruments, Inc., Winooski, VT, USA).
Colony formation
The HT-29 cells from exponentially growing cultures
were seeded into 12-well culture plates at a density of
1×105 cells/well and were treated with 1, 3 or 5 mg/ml
EEHDW and/or IL-6 for 24 h, using the aforementioned protocol. The
cells were subsequently harvested and seeded into six-well plates
at a final concentration of 1×103 cells/well in 2 ml
fresh medium. Following incubation for eight days in a 37°C
humidified incubator with an atmosphere of 5% CO2, the
formed colonies were fixed in MeOH-HAc (v/v dilution, 3:1) for 10
min, stained with crystal violet and counted. The data were
normalized to the viability or survival of control cells, set as
100%.
Cell cycle analysis
A total of 2.5×105 HT-29 cells were
seeded into six-well plates in 2 ml medium and treated with 1, 3 or
5 mg/ml EEHDW and/or IL-6 for 24 h. The cells were harvested and
adjusted to a concentration of 2×105 cells/ml. Following
cell staining with a propidium iodide (PI) cell cycle assay kit,
the cell cycle progression of the HT-29 cells was determined using
fluorescence-activated cell sorting (FACS). Briefly, the cells were
fixed in 70% ethanol at 4°C overnight, then the fixed cells were
washed twice with cold PBS, and incubated for 30 min with RNase (8
μg/ml) and PI (10 μg/ml). The fluorescence signal was detected
through the FL1 channel of the flow cytometer (FACSCalibur; BD
Biosciences, Franklin Lakes, NJ, USA) and the proportion of DNA in
various phases of the cell cycle was analyzed using ModFit LT
software (version 3.0; Verity Software House, Inc., Topsham, ME,
USA).
Detection of apoptosis by FACS with
Annexin V/PI and DAPI staining
A total of 2×105 HT-29 cells were seeded
into six-well plates in 2 ml medium and treated with 1, 3 or 5
mg/ml EEHDW and/or IL-6 for 24 h. Subsequently, the apoptosis rate
of the HT-29 cells was determined by performing FACS, using a
FACSCalibur cell analyzer (BD Biosciences) and an Annexin
V-fluorescein isothiocyanate/PI kit, according to the
manufacturer’s instructions. In this assay, an Annexin V/PI
double-negative population indicates viable cells, and Annexin
V-positive/PI-negative or Annexin V/PI double-positive populations
represent cells undergoing early or late apoptosis,
respectively.
To verify the role of EEHDW in inducing HT-29 cell
apoptosis, apoptotic morphology (chromatin condensation and/or
nuclear fragmentation) was monitored in DAPI-stained cells. The
HT-29 cells were seeded into 12-well culture plates at a density of
1×105 cells/well, and treated with 1, 3 or 5 mg/ml EEHDW
and/or IL-6 for 24 h. Subsequently, the cells were washed in PBS,
fixed with 4% paraformaldehyde for 10 min and stained with DAPI (4
μg/ml) for 10 min at room temperature. Cover slips containing the
cells were washed with PBS and observed under fluorescence
microscopy (Leica DMI4000B; Leica Camera AG, Solms, Germany). Cells
with clearly defined, condensed nuclei were considered to be
apoptotic cells.
Analysis of caspase-9/-3 activation
The activity of caspase-9 and -3 was determined by
performing a colorimetric assay provided in the caspase-9 and-3
activation kit (Invitrogen Life Technologies), in accordance with
the manufacturer’s instructions. Briefly, following treatment with
1, 3 or 5 mg/ml EEHDW and/or IL-6 for 24 h, the HT-29 cells were
lysed with the provided lysis buffer for 30 min on ice. The lysed
cells were centrifuged at 16,000 × g for 10 min and the protein
concentration of the clarified supernatant was determined using the
BCA assay, according to the manufacturer’s instructions.
Subsequently, 100 μg protein was incubated with 50 μl of the
specific colorimetric tetrapeptides [Leu-Glu-His-Asp-p-nitroaniline
(pNA; specific substrate of caspase-9) or Asp-Glu-Val-Asp-pNA
(specific substrate of caspase-3)] at 37°C in the dark for 2 h.
Samples were read at a wavelength of 405 nm in an ELISA reader
(Model EXL800; BioTek Instruments, Inc.,). The data were normalized
to the activity of the caspases in control cells (treated with PBS
vehicle) and represented as a fold value of the control.
Reverse transcription-polymerase chain
reaction (RT-PCR) analysis
A total of 2×105 HT-29 cells were seeded
into six-well plates in 2 ml medium and treated with 1, 3 or 5
mg/ml EEHDW and/or IL-6 for 24 h. Total RNA was isolated using
TRIzol reagent (Invitrogen Life Technologies) and 1 μg
oligo(dT)-primed RNA was reverse-transcribed using SuperScript
reverse transcriptase (Promega Corporation), according to the
manufacturer’s instructions. PCR was performed on the complementary
DNA to determine the quantity of cyclin D1, CDK4, Bcl-1 and Bax
mRNA, with GAPDH used as an internal control. Samples were analyzed
by gel electrophoresis (1.5% agarose) and the DNA bands were
analyzed using a Gel Documentation system (Model Gel Doc 2000;
Bio-Rad Laboratories, Hercules, CA, USA).
Western blot analysis
A total of 2×105/ml HT-29 cells were
seeded into flasks and pre-treated with 1, 3 or 5 mg/ml EEHDW for 1
h. Subsequently IL-6 stimulation was performed for 15 min for
pSTAT3 and STAT3 detection, or 24 h for the analysis of the protein
expression of cyclin D1, CDK4, Bcl-2, Bax and Bcl-2. The treated
cells were lysed with mammalian cell lysis buffer containing
various protein inhibitors and the total protein concentrations
were determined by performing a BCA assay. An equal quantity of
protein from each cell lysate was subjected to SDS-PAGE and
transferred onto polyvinylidene difluoride membranes. The membranes
were blocked for 2 h with 5% skimmed dry milk and incubated with
the appropriate primary antibody directed against STAT3, p-STAT3,
cyclin D1, CDK4, Bcl-2, Bax or β-actin (dilution, 1:1,000)
overnight at a temperature of 4°C. Appropriate HRP-conjugated
secondary antibodies with chemiluminescence detection were used to
image the antibody-detected proteins.
Statistical analysis
All data summarized in the bar graphs are the mean
of three independent experiments, and data were analyzed using the
SPSS software package for Windows (version 17.0; SPSS, Inc.,
Chicago, IL, USA). Furthermore, statistical analysis of the data
was performed using Student’s t-test and an analysis of variance.
P<0.05 was considered to indicate a statistically significant
difference.
Results
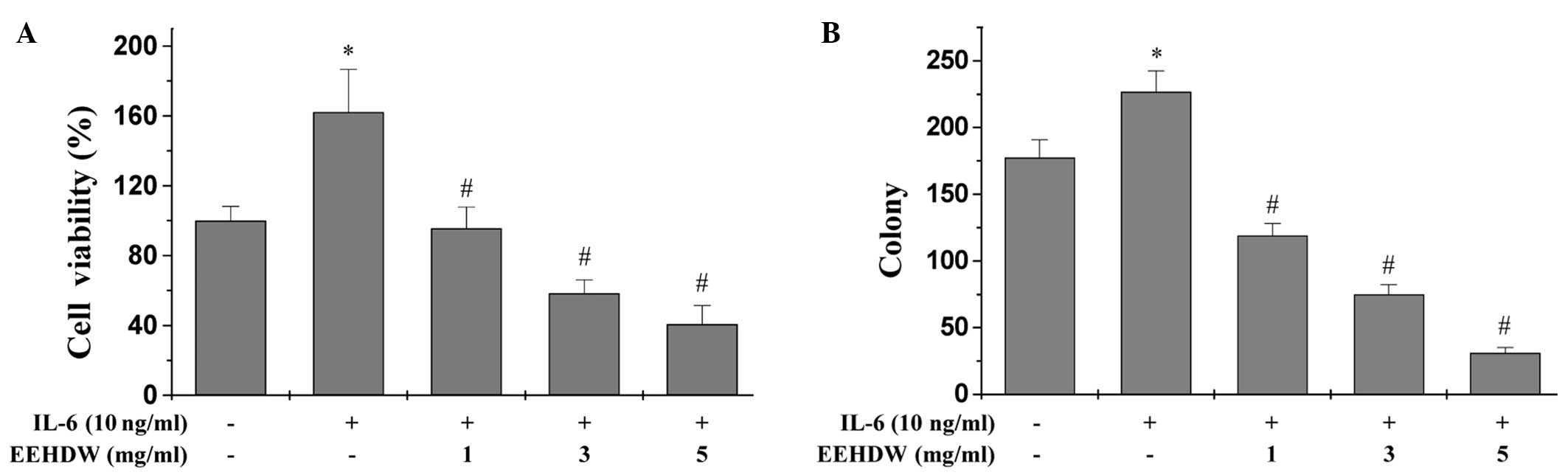
EEHDW inhibits the growth of HT-29
cells
Our previous study observed that EEHDW reduced the
viability and proliferation of HT-29 cells in the absence of IL-6
stimulation (17). To determine
whether the potency of EEHDW was maintained under inflammatory
conditions, the effect of EEHDW on HT-29 cell viability was
measured in the presence of IL-6 by performing an MTT assay
(Fig. 1A). IL-6 stimulation
appeared to significantly enhance the viability of HT-29 cells by
162.17% compared with the control cells (P<0.05). By contrast,
treatment with 1, 3 and 5 mg/ml EEHDW for 24 h reduced the cell
viability of the IL-6-stimulated cells in a dose-dependent manner
from 95.60 to 40.76% (P<0.05). To determine whether EEHDW was
effective at preventing multiple rounds of cell division, the
EEHDW-treated HT-29 cells were examined by performing a colony
formation assay (Fig. 1B).
Treatment with increasing doses (1, 3 and 5 mg/ml) of EEHDW for 24
h significantly reduced the survival rate of the IL-6-stimulated
cells by 32.8, 57.6 and 82.5%, respectively (P<0.05).
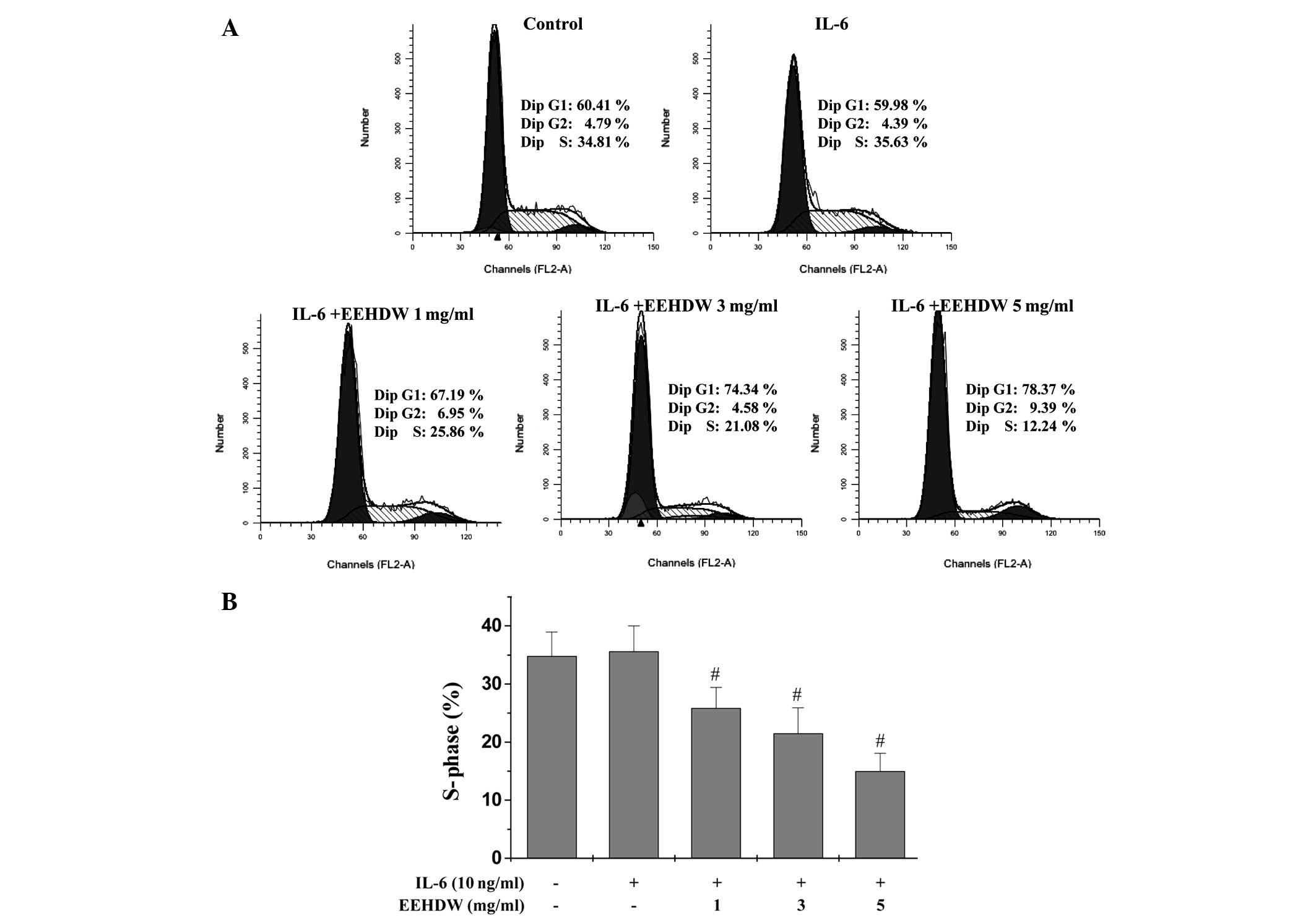
EEHDW blocks G1/S progression
of HT-29 cells
The G1/S transition is one of two major
checkpoints that regulate the cell cycle and cell proliferation.
Our previous study observed that EEHDW blocked G1/S cell
cycle progression in the absence of IL-6 stimulation (17). Thus, the present study aimed to
investigate whether the effect of EEHDW on IL-6 stimulated HT-29
cells would be similar (Fig. 2A and
B). Following staining with PI and FACS analysis, the
percentage of S-phase cells was not significantly different between
the untreated control and IL-6-stimulated HT-29 cells (34.81 vs.
35.63%, respectively; P>0.05). However, for the HT-29 cells
treated with increasing doses of EEHDW (1, 3 or 5 mg/ml), a
significant decrease in the percentage of S-phase cells was
observed (25.86, 21.08 and 12.24%, respectively; P<0.05). These
results indicate that EEHDW inhibits IL-6-stimulated HT-29
proliferation by blocking progression from the G1 phase
to the S phase of the cell cycle.
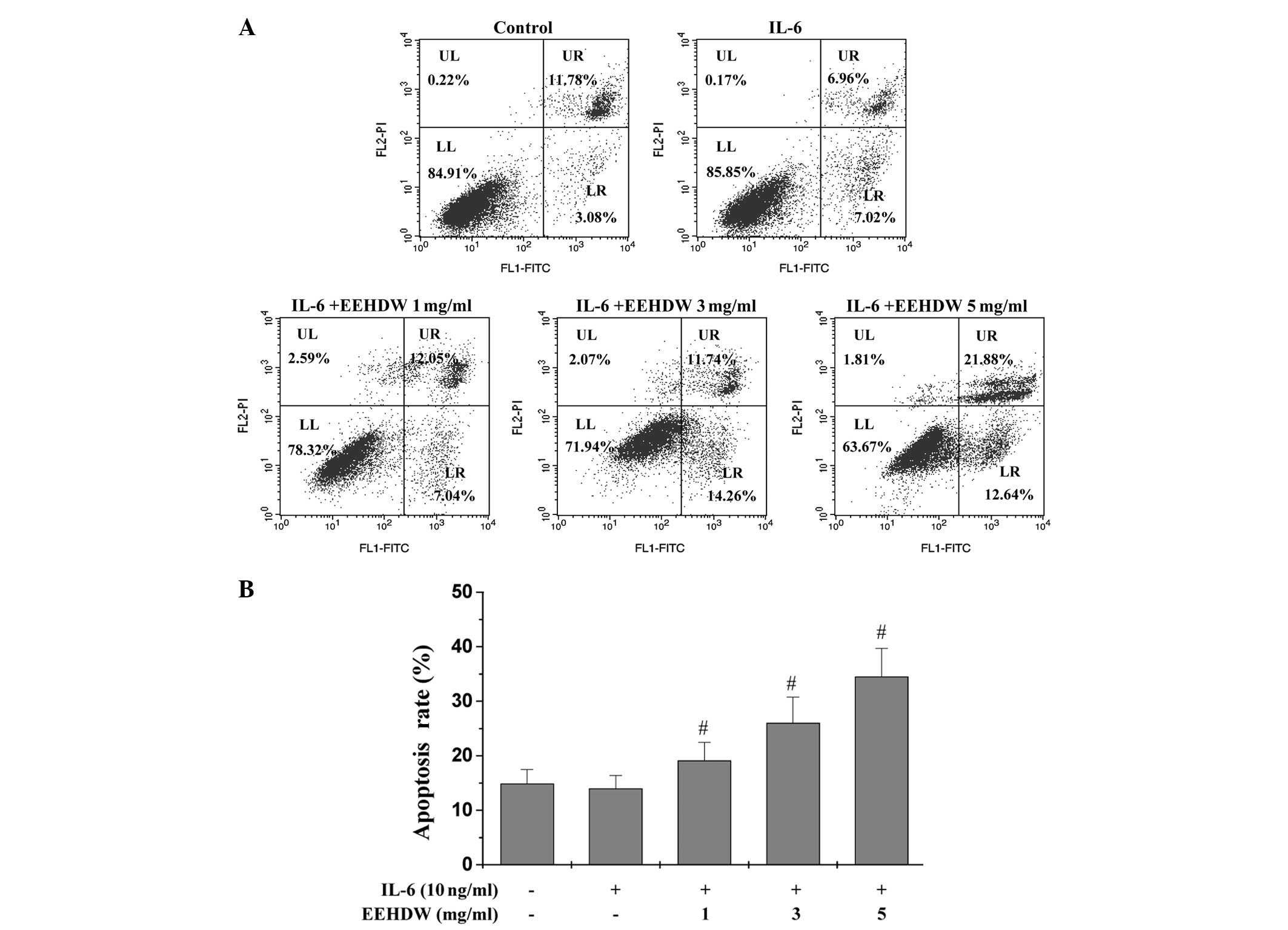
EEHDW induces HT-29 cell apoptosis
Our previous study observed that EEHDW induced
apoptosis in HT-29 cells in the absence of IL-6 stimulation
(20). To determine whether EEHDEW
induces cell apoptosis during cytokine-mediated activation and
proliferation, the induction of apoptosis in the IL-6-stimulated
HT-29 cells was determined by performing Annexin V/PI staining and
FACS analysis (Fig. 3A and B). In
comparison to the unstimulated cells, stimulation with 10 ng/ml
IL-6 did not significantly alter the proportion of the apoptotic
cells (P>0.05). By contrast, EEHDW treatment significantly
increased the percentage of cells undergoing early and late
apoptosis in a dose-dependent manner (P<0.05 vs. cells
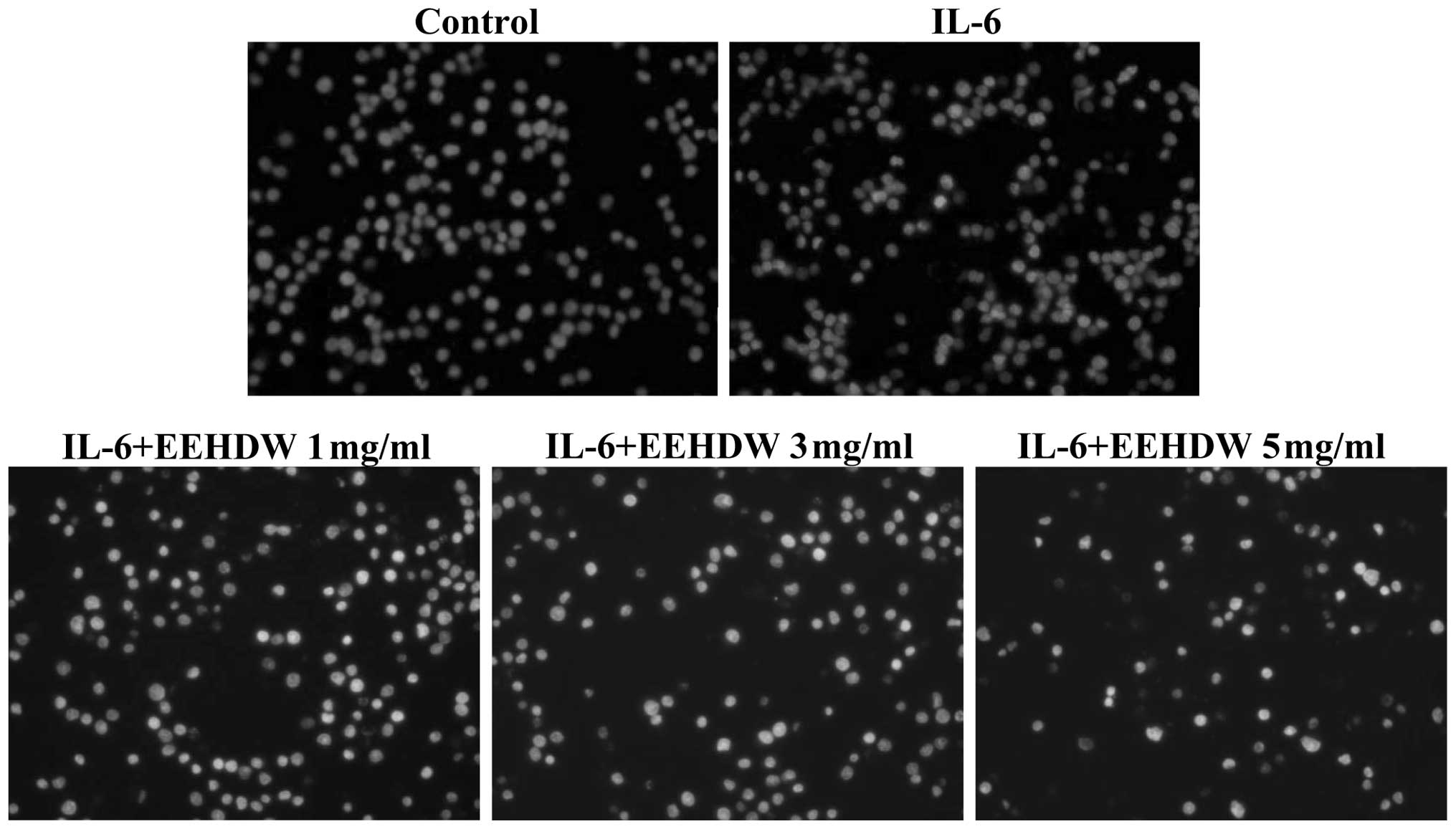
stimulated with IL-6 alone). In addition, the cellular morphology
and extent of DNA condensation of the apoptotic HT-29 cells were
examined by DAPI staining (Fig. 4).
Nuclei staining of the HT-29 cells treated with EEHDW was more
intense than the untreated cells, indicating that EEHDW promotes
HT-29 cell apoptosis in the presence of IL-6 stimulation.
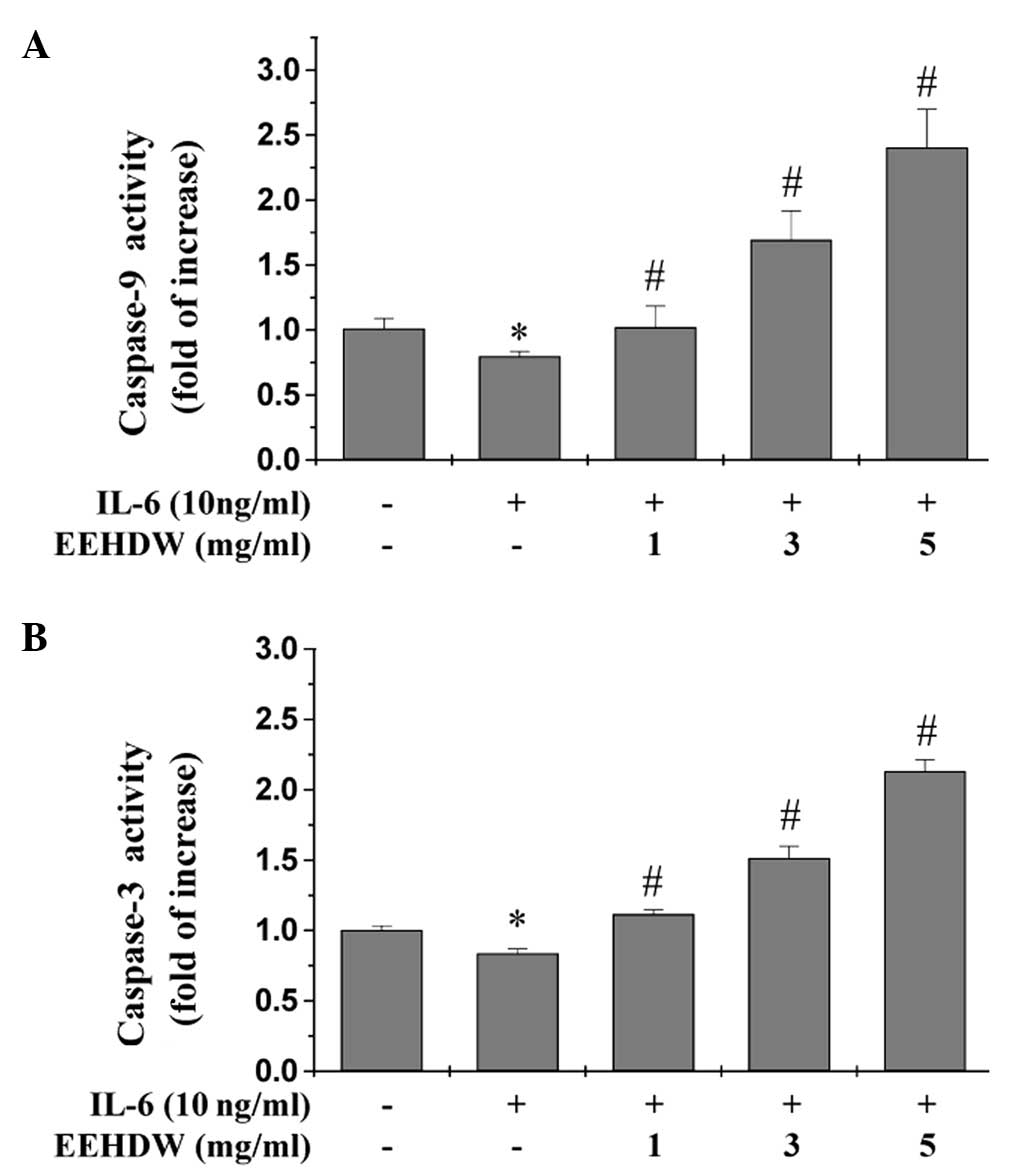
EEHDW induces the activation of
caspases-9 and -3 in HT-29 cells
Our previous study identified the activation of
caspases-9 and -3 in EEHDW-treated HT-29 cells in the absence of
IL-6 stimulation (20). Caspases
are cytoplasmic, aspartate-specific cysteine proteases whose
activation is required for apoptosis, and increased expression of
anti-apoptotic factors by the IL-6/STAT3 signaling pathway may
reduce caspase-mediated apoptosis in cancer cells (21). As expected, stimulation of the HT-29
cells with IL-6 alone significantly inhibited the activation of
caspases-9 and -3 (Fig. 5A and B).
By contrast, EEHDW treatment significantly and dose-dependently
induced activation of caspases-9 and -3 in the HT-29 cells
(P<0.05 vs. the cells stimulated with IL-6 alone; Fig. 5A and B).
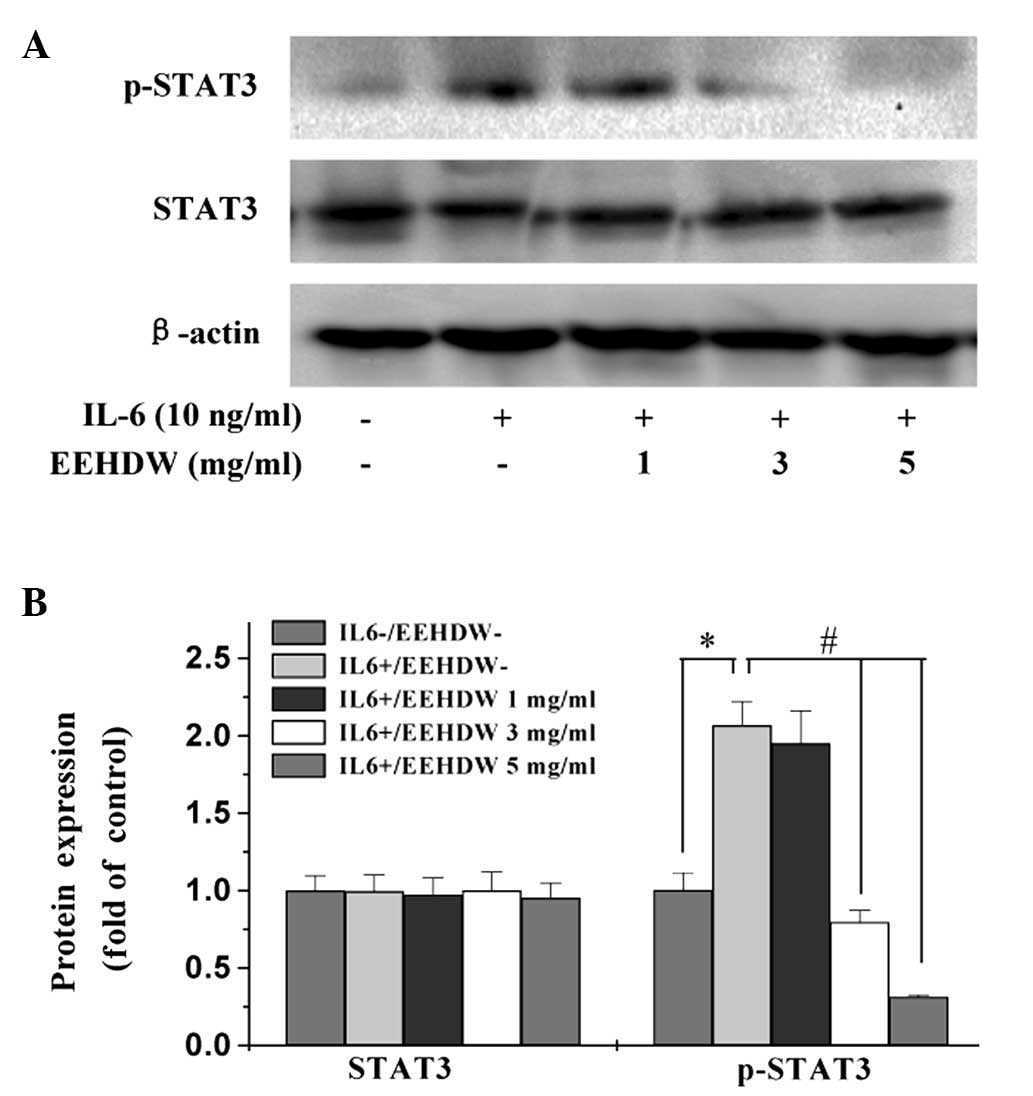
EEHDW inhibits IL-6-mediated STAT3
activation in HT-29 cells
Numerous human cancer cell lines, including HT-29,
do not constitutively express p-STAT3 in vitro, however,
previous studies have demonstrated that IL-6 can stimulate STAT3
activation in HT-29 cells (22).
Thus, the present study stimulated STAT3 activation by
administering IL-6 to the HT-29 cells, and western blot analysis of
the cell lysates was performed to determine the phosphorylation
levels of STAT3 at Tyr705. Stimulation of the HT-29
cells with IL-6 (10 ng/ml) significantly increased the protein
expression levels of p-STAT3, however, phosphorylation was
significantly inhibited by EEHDW in a dose-dependent manner
(P<0.05) (Fig. 6). By contrast,
the protein expression level of non-phosphorylated STAT3 remained
unchanged following treatment with IL-6 and/or EEHDW.
EEHDW significantly downregulates the
mRNA and protein expression levels of cyclin D1, CDK4, Bcl-1 and
Bax in HT-29 cells
To investigate the underlying mechanism of action in
EEHDW-treated HT-29 cells, RT-PCR and western blot analyses were
performed to examine the effect of EEHDW administration on the
expression levels of various important target genes of the
IL-6/STAT3 signaling pathway. These genes included
pro-proliferative cyclin D1 and CDK4, anti-apoptotic Bcl-1, and
pro-apoptotic Bax. Excluding CDK4 mRNA expression, the protein and
mRNA expression levels of cyclin D1, CDK4, and Bcl-1 were not
significantly altered following IL-6 stimulation (P>0.05;
Fig. 7). By contrast, EEHDW
treatment significantly reduced the IL-6-mediated expression of all
three genes at the transcriptional and translational levels
(P<0.05). Furthermore, although the mRNA and protein expression
levels of Bax were significantly decreased in the presence of IL-6
stimulation (P<0.05), a significant increase in the expression
levels of pro-apoptotic Bax were observed in the IL-6-stimulated
HT-29 cells treated with various concentrations of EEHDW
(P>0.05; Fig. 7).
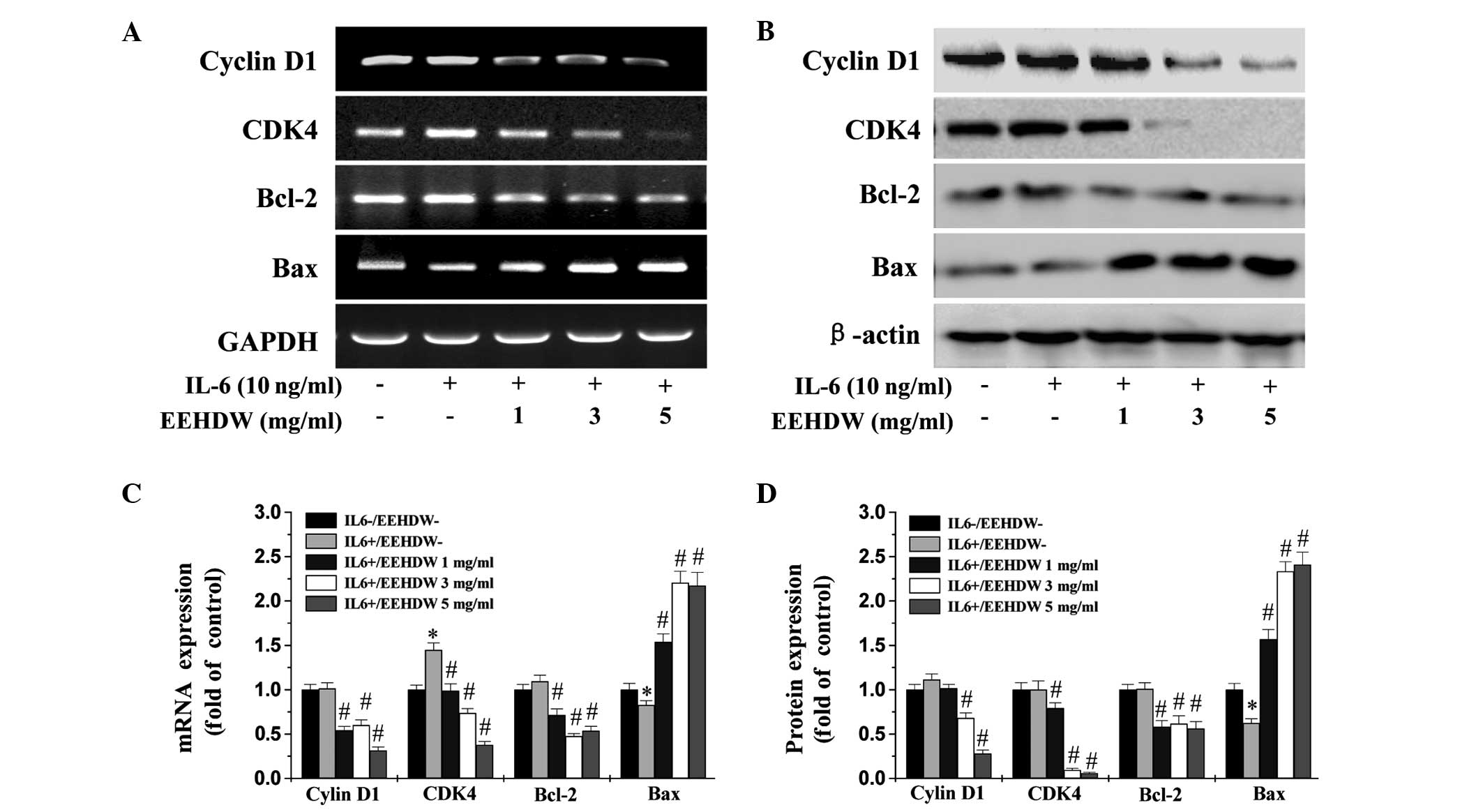 | Figure 7Effect of EEHDW treatment on the mRNA
and protein expression levels of Bcl-2, Bax, cyclin D1 and CDK4 in
HT-29 cells. The HT-29 cells were pretreated with the indicated
doses of EEHDW for 1 h prior to IL-6 stimulation for 24 h. Cell
lysates were prepared and assayed for expression of cyclin D1,
Bcl-2, Bax and CDK4 by (A) reverse transcription-polymerase chain
reaction or (B) western blot analysis. GAPDH and β-actin served as
the internal controls for each assay, respectively (n=3). (C-D)
Densitometric analysis. The data were normalized to the mean mRNA
or protein expression level of untreated control, respectively. The
columns represent the mean of three experiments, and the bars
represent the standard deviation from the mean.
*P<0.01 vs. controls; #P<0.05 vs. cells
treated with IL-6 alone. CDK4, cyclin-dependent kinase 4; Bcl-2,
B-cell lymphoma-2; Bax, Bcl-2-associated X protein; IL-6,
interleukin-6; EEHDW, ethanol extract obtained from Hedyotis
diffusa Willd. |
Discussion
Specific inhibitors that target a single signaling
pathway may be less effective for the treatment of complex tumor
systems compared with multi-targeted agents, and the long-term use
of multiple single-target-based agents may lead to drug resistance
and negative side-effects (23).
Although the use of Chinese herbal medicines as an adjunctive
therapy for CRC has been widespread in Asia, the efficacy of these
treatments has not been well defined. Specific herbal extracts or
mixtures within traditional Chinese medicines have demonstrated
anticancer properties with fewer side-effects compared with current
anticancer treatment strategies, such as chemotherapeutic compounds
and antibodies; therefore, recent studies have reexamined the
therapeutic potential of traditional herbal medicines (24–26).
Among the cytokines linked to
inflammation-associated cancer, IL-6 appears to drive oncogenesis
via downstream activation of the JAK/STAT3 signaling pathway.
Additionally, dysregulation of the IL-6-mediated JAK/STAT3
signaling pathway is closely associated with the development of a
diverse range of solid tumors in humans, including CRC (27,28).
Thus, modulation of the IL-6/JAK/STAT3 signaling pathway is
currently being analyzed with the aim of developing novel therapies
for CRC (29–31). IL-6 is key in promoting cellular
proliferation and the inhibition of apoptosis (32), and acts by binding to its receptor
(soluble IL-6R) and co-receptor [glycoprotein 130 (gp130)],
resulting in activation of the associated Janus kinases (JAKs).
Subsequently, the activated JAKs phosphorylate gp130, leading to
the recruitment and activation of STAT3 (27), an important transcription factor
that is essential in cell survival and proliferation (33,34).
Furthermore, overexpression of various genes, including cyclin D1
and Bcl-1, mediated by the abnormal activation of IL-6/STAT3, leads
to excessive cell proliferation and apoptotic resistance, which may
result in tumorigenesis (8–11).
HDW is a traditional Chinese herbal medicine that
exhibits anticancer activities (17,19–21).
In the present study, MTT and colony formation assays were used to
demonstrate that EEHDW reduces significantly cell viability
following IL-6 stimulation (Fig.
1). Although IL-6 stimulation increased the growth of the HT-29
cells, EEHDW treatment significantly increased the number of
apoptotic cells in a dose-dependent manner (Fig. 3). Furthermore, the percentage of
IL-6-stimulated HT-29 cells in the S-phase significantly decreased
compared with the controls cells following treatment with
increasing concentrations of EEHDW (Fig. 2). In addition, IL-6 stimulation
significantly increased the protein level of pSTAT3; however,
phosphorylation of STAT3 was significantly inhibited by the
administration of EEHDW in a dose-dependent manner (Fig. 6). Although IL-6 stimulation markedly
increased the expression levels of various important target genes
of the IL-6/STAT3 pathway, EEHDW treatment significantly reduced
IL-6-induced mRNA and protein expression levels of cyclin D1, CDK4,
and Bcl-1 (Fig. 7). These data
indicate that EEHDW may be a useful therapeutic agent for the
treatment of CRC.
In conclusion, HDW is composed of a number of
natural products, each of which targets different sites, resulting
in the regulation of multiple signaling pathways. The current study
provided evidence that the anticancer activity of EEHDW on HT-29
cells acts via the IL-6/STAT3 signaling pathway. However, it
remains unknown whether HDW is able to affect other cancer-related
signaling pathways, such as mitogen-activated protein kinase,
phosphoinositol 3 kinase/Akt and Notch. Therefore, clarification of
the molecular mechanisms associated with HDW treatment of cancer is
required to develop improved multi-target agents for cancer
therapy.
Acknowledgements
The present study was supported by the Research Fund
for the Doctoral Program of Higher Education of China (grant no.
20133519110003).
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Akira S, Nishio Y, Inoue M, Wang XJ, Wei
S, Matsusaka T, Yoshida K, Sudo T, Naruto M and Kishimoto T:
Molecular cloning of APRF, a novel IFN-stimulated gene factor 3
p91-related transcription factor involved in the gp130-mediated
signaling pathway. Cell. 77:63–71. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhong B, Liu Q and Liu Y, Xiong X and Liu
Y: Expressions of STAT3, p-STAT3 and E-cadherin in colorectal
cancer and clinical implications. Zhonghua Wei Chang Wai Ke Za Zhi.
17:594–597. 2014.(In Chinese). PubMed/NCBI
|
|
4
|
Bromberg J and Wang TC: Inflammation and
cancer: IL-6 and STAT3 complete the link. Cancer Cell. 15:79–80.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang C, Yang G, Jiang T, Huang K, Cao J
and Qiu Z: Effects of IL-6 and AG490 on regulation of Stat3
signaling pathway and invasion of human pancreatic cancer cells in
vitro. J Exp Clin Cancer Res. 29:512010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Adam N, Rabe B, Suthaus J, Grötzinger J,
Rose-John S and Scheller J: Unraveling viral interleukin-6 binding
to gp130 and activation of STAT-signaling pathways independently of
the interleukin-6 receptor. J Virol. 83:5117–5126. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li X, Wang Y, Han C, Li P and Zhang H:
Colorectal cancer progression is associated with accumulation of
Th17 lymphocytes in tumor tissues and increased serum levels of
interleukin-6. Tohoku J Exp Med. 233:175–182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aneknan P, Kukongviriyapan V, Prawan A,
Kongpetch S, Sripa B and Senggunprai L: Luteolin arrests cell
cycling, induces apoptosis and inhibits the JAK/STAT3 pathway in
human cholangiocarcinoma cells. Asian Pac J Cancer Prev.
15:5071–5076. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Darnell JE Jr: STATs and gene regulation.
Science. 277:1630–1635. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zushi S, Shinomura Y, Kiyohara T, Miyazaki
Y, Kondo S, Sugimachi M, Higashimoto Y, Kanayama S and Matsuzawa Y:
STAT3 mediates the survival signal in oncogenic ras-transfected
intestinal epithelial cells. Int J Cancer. 78:326–330. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Masuda M, Suzui M, Yasumatu R, Nakashima
T, Kuratomi Y, Azuma K, Tomita K, Komiyama S and Weinstein IB:
Constitutive activation of signal transducers and activators of
transcription 3 correlates with cyclin D1 overexpression and may
provide a novel prognostic marker in head and neck squamous cell
carcinoma. Cancer Res. 62:3351–3355. 2002.PubMed/NCBI
|
|
12
|
Van Cutsem E and Nordlinger B: Advanced
colorectal cancer: ESMO Clinical Practice Guidelines for treatment.
Ann Oncol. 21(Suppl 5): v93–v97. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meng QX, Roubin RH and Hanrahan JR:
Ethnopharmacological and bioactivity guided investigation of five
TCM anticancer herbs. J Ethnopharmacol. 148:229–238. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Niu Y and Meng QX: Chemical and
preclinical studies on Hedyotis diffusa with anticancer potential.
J Asian Nat Prod Res. 15:550–565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yeh YC, Chen HY, Yang SH, Lin YH, Chiu JH,
Lin YH and Chen JL: Hedyotis diffusa combined with Scutellaria
barbata are the core treatment of Chinese herbal medicine used for
breast cancer patients: a population-based study. Evid Based
Complement Alternat Med. 2014:2023782014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin J, Wei L, Shen A, Cai Q, Xu W, Li H,
Zhan Y, Hong Z and Peng J: Hedyotis diffusa Willd extract
suppresses Sonic hedgehog signaling leading to the inhibition of
colorectal cancer angiogenesis. Int J Oncol. 42:651–656. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin M, Lin J, Wei L, Xu W, Hong Z, Cai Q,
Peng J and Zhu D: Hedyotis diffusa Willd extract inhibits HT-29
cell proliferation via cell cycle arrest. Exp Ther Med. 4:307–310.
2012.PubMed/NCBI
|
|
18
|
Lin J, Wei L, Xu W, Hong Z, Liu X and Peng
J: Effect of Hedyotis diffusa Willd extract on tumor angiogenesis.
Mol Med Rep. 4:1283–1288. 2011.PubMed/NCBI
|
|
19
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa Willd
inhibits colorectal cancer growth in vivo via inhibition of STAT3
signaling pathway. Int J Mol Sci. 13:6117–6128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin J, Chen Y, Wei L, Chen X, Xu W, Hong
Z, Sferra TJ and Peng J: Hedyotis diffusa Willd extract induces
apoptosis via activation of the mitochondrion-dependent pathway in
human colon carcinoma cells. Int J Oncol. 37:1331–1338.
2010.PubMed/NCBI
|
|
21
|
Fiandalo MV and Kyprianou N: Caspase
control: protagonists of cancer cell apoptosis. Exp Oncol.
34:165–175. 2012.PubMed/NCBI
|
|
22
|
Lin J, Chen Y, Wei L, Shen A, Sferra TJ,
Hong Z and Peng J: Ursolic acid promotes colorectal cancer cell
apoptosis and inhibits cell proliferation via modulation of
multiple signaling pathways. Int J Oncol. 43:1235–1243.
2013.PubMed/NCBI
|
|
23
|
Wang S, Wu X, Tan M, Gong J, Tan W, Bian
B, Chen M and Wang Y: Fighting fire with fire: poisonous Chinese
herbal medicine for cancer therapy. J Ethnopharmacol. 140:33–45.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qi F, Li A, Inagaki Y, et al: Chinese
herbal medicines as adjuvant treatment during chemo- or
radio-therapy for cancer. Biosci Trends. 4:297–307. 2010.
|
|
25
|
Cheng HM, Li CC, Chen CY, Lo HY, Cheng WY,
Lee CH, Yang SZ, Wu SL, Hsiang CY and Ho TY: Application of
bioactivity database of Chinese herbal medicine on the therapeutic
prediction, drug development, and safety evaluation. J
Ethnopharmacol. 132:429–437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang SW and Sun YM: The IL-6/JAK/STAT3
pathway: potential therapeutic strategies in treating colorectal
cancer (Review). Int J Oncol. 44:1032–1040. 2014.PubMed/NCBI
|
|
27
|
He Z, Ke J, He X, Lian L, Sun L, Chen Z,
Wu X and Lan P: Inflammation promotes the development of
colitis-associated colorectal cancer. Zhonghua Wei Chang Wai Ke Za
Zhi. 17:706–710. 2014.(In Chinese). PubMed/NCBI
|
|
28
|
Dai Y, Jiao H, Teng G, Wang W, Zhang R,
Wang Y, Hebbard L, George J and Qiao L: Embelin reduces
colitis-associated tumorigenesis through limiting IL-6/STAT3
signaling. Mol Cancer Ther. 13:1206–1216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chang Q, Bournazou E, Sansone P, Berishaj
M, Gao SP, Daly L, Wels J, Theilen T, Granitto S, Zhang X, et al:
The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and
metastasis. Neoplasia. 15:848–862. 2013.PubMed/NCBI
|
|
30
|
Zhao Y, Yao J, Wu XP, Zhao L, Zhou YX,
Zhang Y, You QD, Guo QL and Lu N: Wogonin suppresses human alveolar
adenocarcinoma cell A549 migration in inflammatory microenvironment
by modulating the IL-6/STAT3 signaling pathway. Mol Carcinog. Jun
29–2014.(Epub ahead of print). View
Article : Google Scholar
|
|
31
|
Landskron G, De la Fuente M, Thuwajit P,
Thuwajit C and Hermoso MA: Chronic inflammation of cytokines in the
tumor microenvironment. J Immunol Res. 2014:1491852014. View Article : Google Scholar
|
|
32
|
Heinrich PC, Behrmann I, Haan S, Hermanns
HM, Müller-Newen G and Schaper F: Principles of interleukin
(IL)-6-type cytokine signalling and its regulation. Biochem J.
374:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bromberg J and Darnell JE Jr: The role of
STATs in transcriptional control and their impact on cellular
function. Oncogene. 19:2468–2473. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Aggarwal BB, Kunnumakkara AB, Harikumar
KB, Gupta SR, Tharakan ST, Koca C, Dey S and Sung B: Signal
transducer and activator of transcription-3, inflammation, and
cancer: how intimate is the relationship? Ann NY Acad Sci.
1171:59–76. 2009. View Article : Google Scholar : PubMed/NCBI
|