Introduction
Abnormalities in the control of the execution of
apoptosis, which contributes to enhancement of tumor progression
and treatment resistance, are observed in many malignancies,
including ovarian cancer (1). Tumor
cells often circumvent the apoptotic cascade and proliferate in the
face of apoptotic stimuli, which facilitates tumor progression and
metastasis (2). Thus, targeting key
apoptosis regulators with a goal to overcome apoptosis resistance
of tumor cells has become the focus of extensive pharmaceutical
research.
Two main pathways leading to apoptosis-associated
caspase activation have been identified: the extrinsic and the
intrinsic apoptotic pathway (3). In
the extrinsic death receptor pathway, apoptosis is initiated
through the binding of death receptor superfamily proteins, such as
TRAIL, to their respective cell surface receptors, leading to
caspase-8 activation. In the intrinsic mitochondrial pathway, a
variety of apoptotic stimuli, including anticancer agents, triggers
changes in mitochondrial membrane permeability and the release of
pro-apoptotic proteins including cytochrome c and Smac.
Cytochrome c triggers caspase-3 activation through the
apoptosome complex, whereas Smac promotes caspase-3 activation by
binding to and neutralizing the inhibitor of the apoptotic proteins
(IAPs) (4).
The IAPs are a family of key apoptosis regulators,
characterized by one or more baculovirus IAP repeats (BIR)
(5). Among them, X-linked IAP
(XIAP), cellular IAP1 (cIAP1), cIAP2 and survivin, function to
block death receptor and mitochondrial-mediated apoptotic pathways
mainly by preventing the activation of procaspases and inhibiting
the enzymatic activity of mature caspases. Evidence indicates that
XIAP, survivin, cIAP1 and cIAP2 are overexpressed in human tumor
tissues and their downregulation confers an apoptosis enhancement
effect in chemoresistant human cancer cells (6). Smac interacts with IAPs, such as XIAP,
CIAP-1, and CIAP-2, at the level of the BIR domains via its
N-terminal AVPI terapeptide binding motif, thereby eliminating the
inhibitory effects of IAPs on caspase-3, -7 and -9 (7–9). Smac
peptides as short as 4-residue (AVPI), derived from the Smac
protein bind to the recombinant XIAP BIR3 domain protein with the
same affinities as the mature Smac protein (9). Previous studies have demonstrated that
the first 4–8 N-terminal amino acids of Smac fused to the
Drosophila antennapedia penetration sequence, a carrier
peptide, were shown to enhance the induction of apoptosis of
diverse antineoplastic agents including paclitaxel, etoposide,
7-ethyl-10-hydroxycamptothecin (SN-38), TRAIL and doxorubicin
(10,11).
Epithelial ovarian cancer (EOC) is the fifth most
frequent cause of cancer-related death in women. The standard
first-line therapy for ovarian cancer includes tumor debulking
followed by chemotherapy treatment with paclitaxel, platinum-based
agents, or combinations of both (12–14).
Despite the initial response to treatment, the majority of patients
relapse and eventually progress to a chemotherapy-resistant state
(14). We previously demonstrated
decreased Smac and increased XIAP expression in EOC tissues, and
ectopic expression of Smac sensitized drug-resistant EOC cells to
undergo apoptosis by TRAIL or paclitaxel (15). In the present study, we synthesized
a cell permeable Smac peptide and evaluated its effect on the in
vitro and in vivo therapeutic efficacy of various
chemotherapy agents, in an effort to provide a more effective
antitumor strategy for EOC.
Materials and methods
Cell line and cell culture
The human ovarian epithelial cancer cell line A2780
was provided by Qilu Hospital Biotechnology Center of Shandong
University. Cells were cultured in RPMI-1640 medium (Gibco/BRL,
Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal
bovine serum (FBS; Gibco/BRL) and incubated in an atmosphere of 5%
CO2 at 37°C.
Design, synthesis and detection of
peptides
The Smac N7 peptide and the control peptide (R-Smac
N7) were designed by conjugating the N-terminal seven amino acids
of Smac, AVPIAQK, which are critical for its binding to IAPs or the
random arrangement sequence of Smac, KIPAQVA with the
Drosophila antennapaedia penetration sequence and
fluorescein isothiocyanate (FITC). All of the peptides were
custom-synthesized by Sangon Biotechnology (Shanghai). The purity
of the peptides as determined by high-performance liquid
chromatography was >95%.
Cell viability assay
Cell viability rate was quantitated using a modified
methylthiazol tetrazolium (MTT) colorimetric assay. Cells were
seeded into 96-well culture plates (4000/well), allowed to adhere
overnight, and then treated with different concentrations of TRAIL
(Chemicon, USA) or paclitaxel (Bristol-Myers Squibb Co., USA) in
the absence or presence of diverse doses of Smac N7 or R-Smac N7
for 24 h, respectively. At the end of each treatment, cells were
incubated with 5 mg/ml MTT (Sigma, USA) for 4 h and then mixed with
dimethyl sulfoxide after the supernatant was removed. Cell
viability was quantitated by reading the dye absorption (A) at 550
nm (A1) and 630 nm (A2) with an automatic multiwell
spectrophotometer (Coda; Bio-Rad, Richmond, CA, USA). All the
experiments were performed in triplicate and reproduced at least
three times.
Flow cytometric analysis
Flow cytometric analysis was performed to identify
the intracellular uptake rate of FITC-tagged Smac N7 peptide and
R-Smac N7 peptide. Briefly, A2780 cells (5×106 cells/ml)
were planted in culture flasks. After most of the cells became
adherent, the cells were treated with 10 μmol/l Smac N7 or R-Smac
N7 for 24 h, respectively. All cells were collected and washed
twice with phosphate-buffered saline after centrifugation, and
intracellular uptake rates were determined by a flow cytometry
assay (FACScan, Becton Dickinson, USA).
Animal tumor models and treatment
Female nude mice (BALB/c, 4–5 weeks old) were
purchased from the Shanghai Experimental Animal Center (Chinese
Academy of Sciences, Shanghai). The mice were housed and maintained
under specific pathogen-free conditions according to the
experimental animal guidelines. A2780 cells were harvested and
re-suspended in RPMI-1640 media. A2780 cells (200 μl)
(2×107/ml) were injected into the right flank of the
mice. Two weeks later, the mice were randomly assigned to six
groups with 5 mice in each group. A total of 0.1 ml PBS, Smac N7
(12.5 mg/kg), TRAIL (10 μg/kg), paclitaxel (20 mg/kg), Smac N7
(12.5 mg/kg) + TRAIL (10 μg/kg), Smac N7 (12.5 mg/kg) + paclitaxel
(20 mg/kg) were intratumorally injected seven times at an interval
of 3 days, respectively. Tumor sizes were measured by length (l)
and width (w) every 4 days, and the tumor volumes were calculated
according to the following formula: Tumor volume = l ×
w2/2. The mice were observed daily for evaluation of
their mental state, diet and stools. Twelve hours following the
last injection, the mice were sacrificed, and the tumors, hearts,
livers, kidneys and spleens were removed. Part of the tissues were
fixed in formalin, embedded in paraffin and stained with
hematoxylin and eosin (H&E), while others were frozen
immediately in liquid nitrogen at −80°C for the western blot assay.
All animal studies were approved and carried out in accordance with
the guidelines of the Experimental ethics review committee of
Shandong University.
Western blotting
Cells or xenograft tumor tissues were lysed in lysis
buffer. The whole cell extracts (30 μg/lane) were equally loaded on
10–12% SDS-PAGE gels and transferred to nitrocellulose membranes
(Millipore, Bedford, MA, USA) using the Bio-Rad electrotransfer
system. The membrane was blocked in Tris-buffered saline with 5%
(w/v) nonfat dry milk, and then incubated with a primary antibody
against XIAP, survivin, PARP (Cell Signaling Technology, Danvers,
MA, USA), or Bcl-2, caspase-3, GAPDH, anti-FITC (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) overnight at 4°C,
respectively. After being washed and incubated with horseradish
peroxidase conjugated second antibody, the proteins were visualized
using an enhanced chemiluminescence (ECL) detection system (Pierce,
Rockford, IL, USA). The bands were examined using a densitometer
analysis system Flurochem 9900–50 (Alpha Innotech, San Leandro, CA,
USA). Band density was analyzed using Bandscan software (Glyko,
USA) and the results were expressed as a ratio of the protein of
interest/GAPDH to correct for loading for each sample.
TUNEL detection of apoptotic tumor
cells
TUNEL was performed with a Fluorometric TUNEL system
(KeyGeen Biotech, Nanjing), following the manufacturer’s protocol.
Dissected tumors were rinsed with PBS for 5 min after incubation
with proteinase K (18 μg/ml) for 20 min, and then blocked with
fetal bovine serum for 15 min at room temperature. TdT reaction mix
(50 μl) was added to the section and incubated in a humidified
chamber for 1 h at 37°C. Then the sections were rinsed with PBS and
assessed using a fluorescence microscope. Cell nuclei with green
fluorescent staining were defined as TUNEL-positive nuclei. Cell
apoptosis was quantitated by counting the number of green
fluorescence-positive cells in five random fields of view in each
section at a magnification of ×200.
Data analysis and statistics
The values are presented as means ± SD, and were
statistically evaluated by analysis of variance (ANOVA).
Statistical analysis was performed with SPSS software (version
13.0; SPSS Inc., Chicago, IL). P<0.05 was considered to indicate
a statistically significant result in all cases.
Results
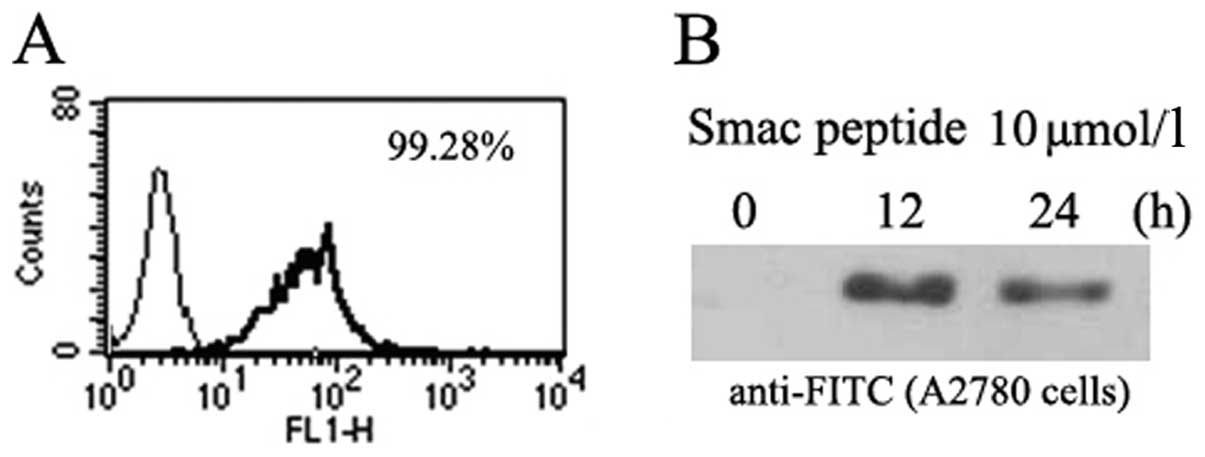
Cellular uptake of the Smac N7
peptide
Following exposure to 10 μmol/l of Smac N7 peptide
for 24 h, the intracellular uptake of FITC-tagged Smac N7 in A2780
cells was 99.28% (Fig. 1A), as
confirmed by flow cytometric analysis (FCM). We then assessed the
cellular uptake of Smac N7 using western blotting. After A2780
cells were treated with Smac N7 (10 μmol/l) for 12 or 24 h, cells
were harvested and subjected to western blotting with anti-FITC
antibodies. The synthetic Smac N7 peptide was successfully taken up
into the cytoplasm of A2780 cells, and the uptake was most
prominent after 12 h (Fig. 1B).
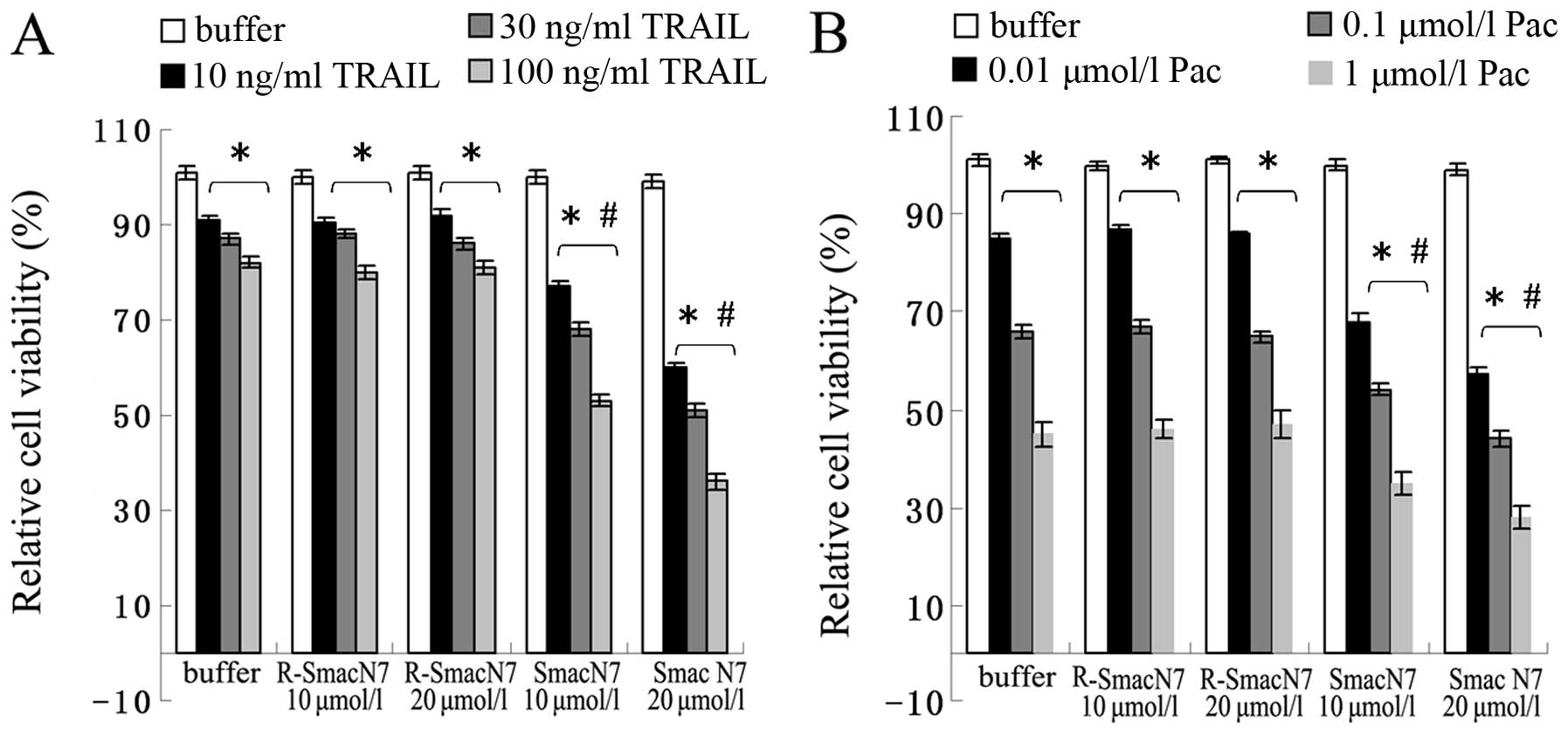
Smac N7 peptide sensitizes A2780 cells to
TRAIL- or paclitaxel-induced cell growth inhibition
Our previous study showed that ectopic expression of
the Smac gene enhanced the therapeutic potential of TRAIL or
paclitaxel in ovarian cancer cells (15). In this study, we explored whether
the Smac N7 peptide has a similar effect. As shown in Fig. 2, the results indicated that the
administration of TRAIL or paclitaxel, but not PBS or Smac N7,
resulted in cell growth inhibition in A2780 cells in a
dose-dependent manner. Moreover, compared with cells treated with
TRAIL or paclitaxel alone, significantly reduced cell viability was
observed in cells treated with TRAIL or paclitaxel accompanied with
Smac N7, respectively, and Smac N7 had a dose-dependent effect on
TRAIL or paclitaxel sensitization in A2780 cells.
Co-treatment with the Smac N7 peptide
potentiates TRAIL- or paclitaxel-induced tumor regression effect
and apoptosis in A2780 cell xenografts in nude mice
Given the TRAIL- or paclitaxel-sensitizing
properties of Smac N7 in vitro, we corroborated these
findings in BALB/c mice bearing A2780 cell xenografts. A2780 cells
were implanted into the right flank of nude mice. The rate of tumor
formation was 100%, and the average time was ~12–14 days. On day 38
after implantation, the A2780 cell tumors of mice treated with PBS
and Smac N7 reached 2021.74±123.19 and 1893.00±131.27
mm3 in volume, respectively (P>0.05). The volumes of
tumors treated with TRAIL, paclitaxel, Smac N7+TRAIL or Smac
N7+paclitaxel were 1476.50±188.90, 1188.25±141.24, 604.40±50.04 and
522.50±75.48 mm3, respectively, which were significantly
smaller than those treated with PBS or Smac N7 alone (P<0.05)
(Figs. 3 and 4). Noticeably, the combination of Smac N7
and TRAIL or paclitaxel resulted in a significant reduction in
tumor volume compared with the treatment of each drug alone
(P<0.05).
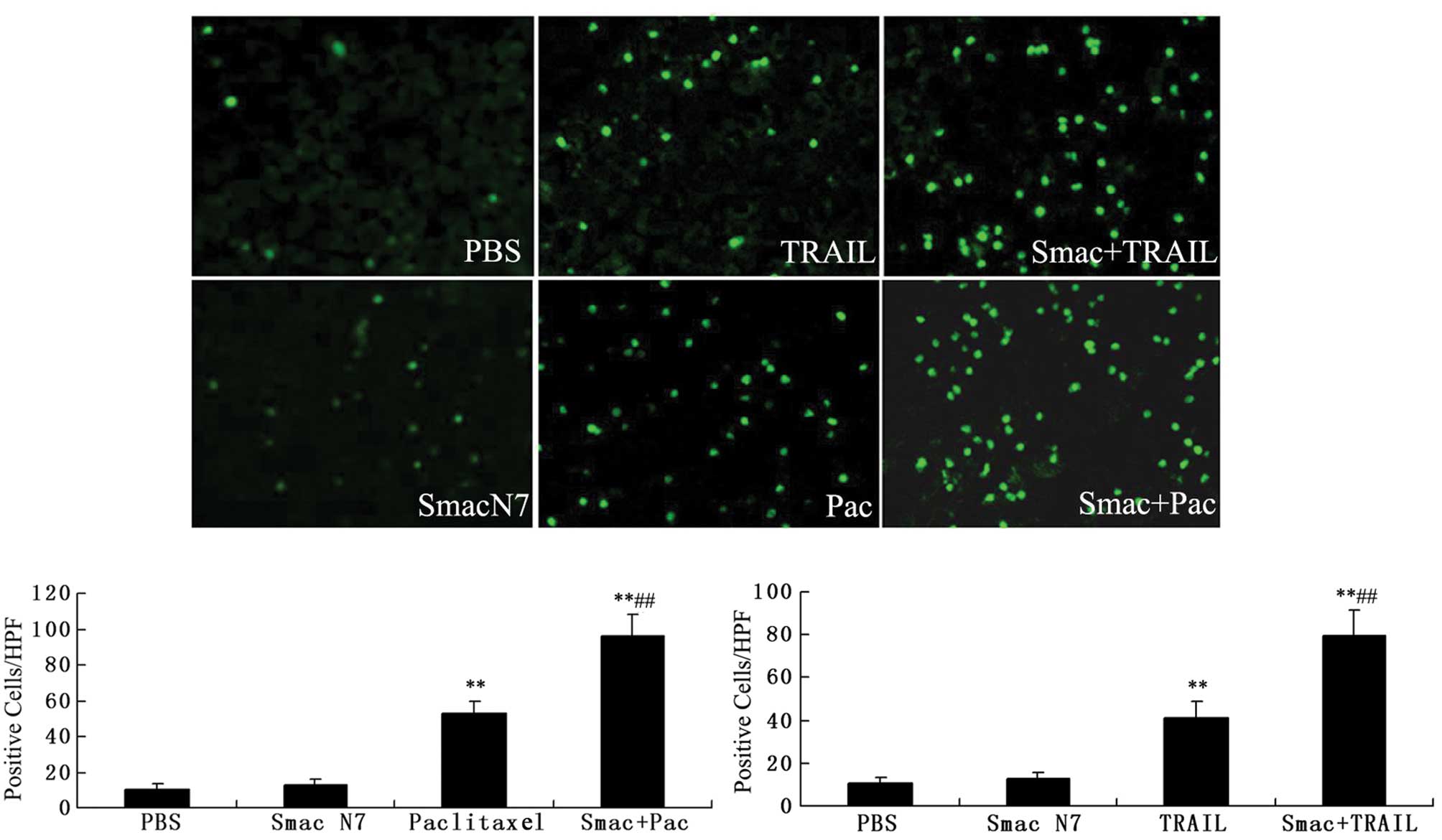
Fluorometric TUNEL assay was performed to detect the
apoptotic cells in the tumor tissues of nude mice. As shown in
Fig. 5, the apoptotic cells
accounted for 41±7.65 or 53±6.29 in the TRAIL or paclitaxel-treated
group versus 12.8±3.11 in the Smac N7-treated group and 10.6±3.0 in
PBS control group (P<0.05), and the apoptotic cells accounted
for 79.2±12.19 or 96±12.39 in the combination treatment groups
(Smac N7+TRAIL or Smac N7+paclitaxel, respectively). These results
suggest that Smac N7 alone had no significant apoptosis-inducing
effect on A2780 cell tumor xenografts, but further enhanced the
apoptosis induction by TRAIL or paclitaxel.
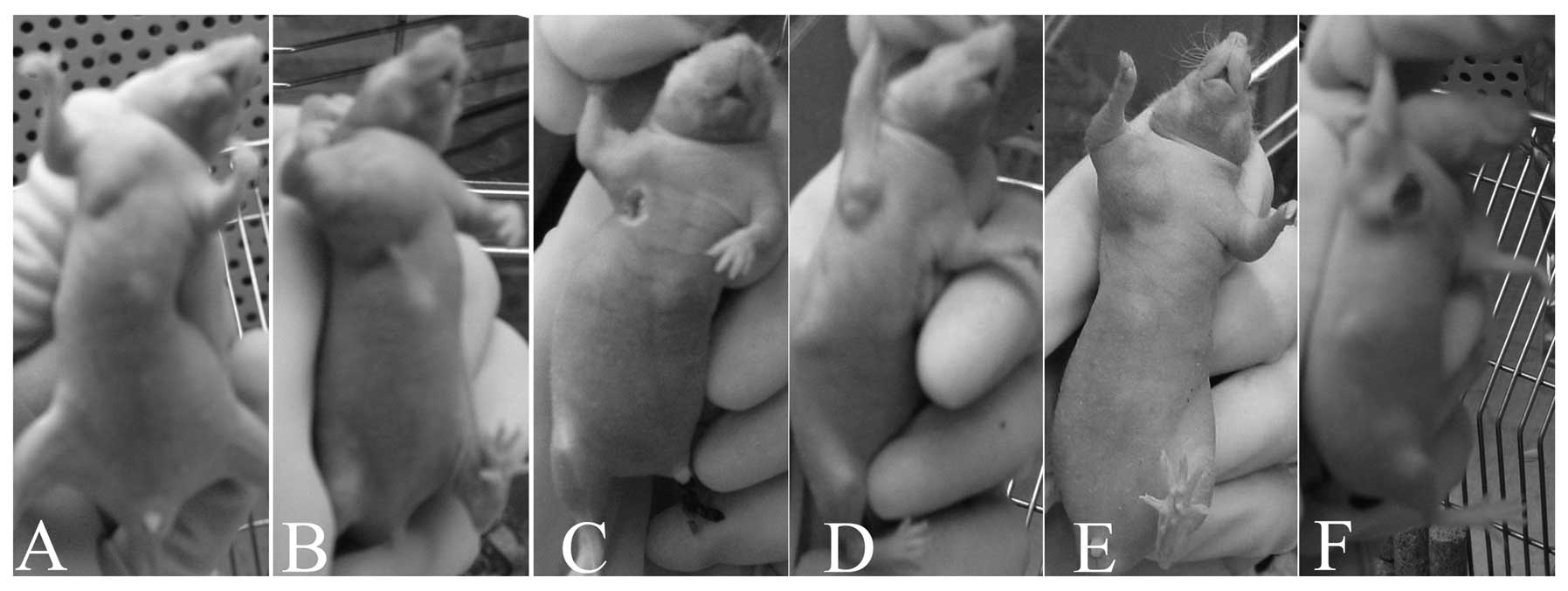
Smac N7 peptide reduces the cytotoxic
effect of TRAIL or paclitaxel on vital organs of nude mice
Although paclitaxel has demonstrated clinical
efficacy in most types of human cancers, adverse vital organ
pathologies and toxicities continue to be of major concern. In our
study, necrosis of mice skin around the injection point was
observed in both the paclitaxel and Smac N7+paclitaxel treatment
groups, but the range of skin necrosis seemed smaller in the latter
group (Fig. 3). We also carried out
H&E staining of the transplanted tumors and the heart, liver,
kidney and spleen of the nude mice. The karyomegaly, anachromasis,
and karyokinesis appearance observed in tumor sections confirmed
the formation of xenograft tumors. No obvious damage was found in
the examined organs of the mice treated with PBS or Smac N7.
Compared with TRAIL, paclitaxel showed severe cytotoxicity to the
liver and kidney of mice. In particular, the damage to the mice
treated with the combination therapy of Smac N7 and paclitaxel or
TRAIL was markedly less than that experienced by the mice treated
with paclitaxel or TRAIL alone. Notably, apart from the PBS-treated
mice, the spleen of all tested mice displayed an enlarged volume
with blood stasis and infiltrating mononuclear phagocytes (Table I).
 | Table IPathologic examination of nude mice
after treatment: the H&E staining of the heart, liver, kidney
and spleen. |
Table I
Pathologic examination of nude mice
after treatment: the H&E staining of the heart, liver, kidney
and spleen.
| Organ |
|---|
|
|
|---|
| Group | Heart | Liver | Spleen | Kidney |
|---|
| PBS | Normal | Normal | Normal | Normal |
| Smac N7 | Normal | Normal | Volume enlarged,
blood stasis, a large number of infiltrating mononuclear phagocytes
were observed (no. 1–5) | Normal |
| TRAIL | No obvious change in
myocardial cells | Normal (no. 1–3)
ballooning degeneration (no. 4, 5) | Volume enlarged,
blood stasis, a large number of infiltrating mononuclear phagocytes
were observed (no. 1–5) | Normal |
| Paclitaxel | No obvious change in
myocardial cells (no. 1–5), vasodilatation with excessive
accumulation of neutrophilic granulocytes in vessel lumen among
cardiac muscle was distinct (no. 2–4) | Significant
hepatocellular degeneration, necrosis in local liver cells, small
abscess, central vein dilation in central portal tract area (no.
1–5) | Volume enlarged,
blood stasis, a large number of infiltrating mononuclear phagocytes
was observed (no. 1–5), small abscess (no. 3, 4) | Glomerular capillary
plexus volume enlarged, glomerular cyst cavity became narrow,
leukocytes accumulated in capillary plexus. No cast in kidney
tubules, few neutrophilic granulocytes in renal interstitium (no.
1–5) |
| Smac N7 + TRAIL | No obvious change in
myocardial cells | Normal | Volume enlarged,
blood stasis, a large number of infiltrating mononuclear phagocytes
were observed (no. 1–5) | Normal |
| Smac N7+ Pac | No obvious change in
myocardial cells | Hepatocellular
degeneration, nidus for abscesses appeared in individual areas | Volume enlarged,
blood stasis, a large number of infiltrating mononuclear phagocytes
were observed (no. 1–5) | Almost normal,
individual kidney glomerulus had a similar appearance as the
paclitaxel treatment group |
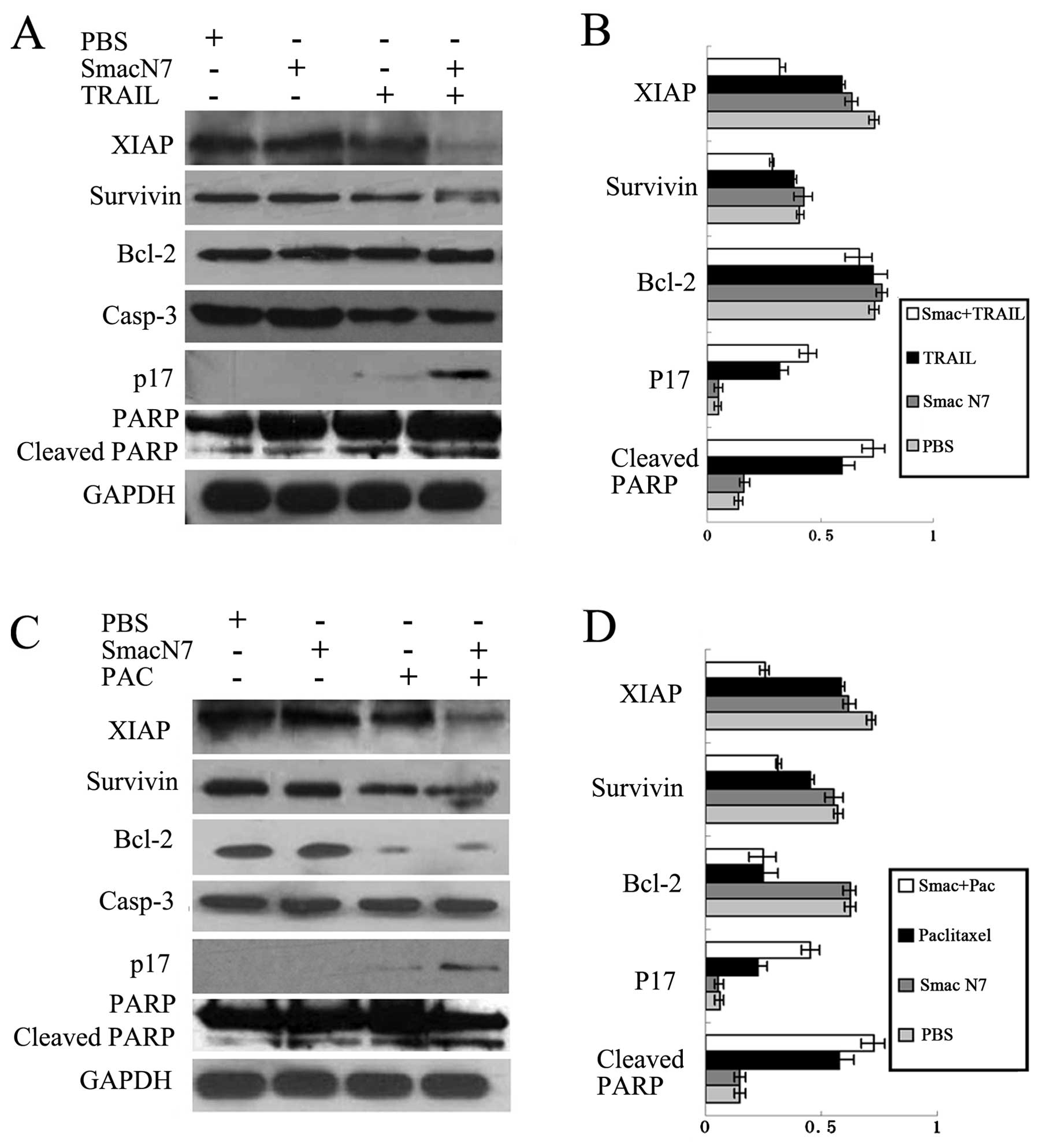
Smac N7 peptide increases TRAIL- or
paclitaxel-induced caspase-3 and PARP activation and downregulates
the protein levels of the IAP family
To gain insight into the mechanisms by which the
combination of Smac N7 peptide and TRAIL or paclitaxel exerts
enhanced anticancer activity, we analyzed the effects of the
combination therapy on caspase-3 activity and on the expression of
XIAP, survivin and Bcl-2 in comparison with either agent alone.
Evaluation of cell lysates revealed that the Smac N7 peptide had no
effect on caspase activation and only a slightly downregulatory
effect on XIAP protein expression, but potentiated TRAIL- or
paclitaxel-induced caspase-3 activation as represented by increased
levels of cleaved caspase-3 and PARP, as well as enhanced TRAIL- or
paclitaxel-induced downregulation of XIAP and survivin in the
xenograft tumor tissues (Fig. 6).
Notably, paclitaxel resulted in the downregulation of Bcl-2, but
paclitaxel plus the Smac N7 peptide did not have an additive
effect. Furthermore, no modification of Bcl-2 was observed in the
TRAIL-treated tumor model.
Discussion
Our previous study showed that the disrupted balance
of XIAP and Smac expression in EOC tissues, and ectopic Smac
expression reversed the resistance to paclitaxel and TRAIL of EOC
cell lines (15). According to
these observations, it was proposed that overexpression of Smac
might be a potent strategy for EOC treatment by sensitization of
tumor cells to chemotherapeutic drugs, yet this procedure has not
been developed at the clinical level.
Based on reports that the suppression of apoptosis
by the IAPs is primarily mediated either directly via their
baculoviral IAP repeat (BIR) domains or indirectly through their
ubiquitin E3 ligase RING domain, and that Smac through its
N-terminal AVPI tetrapeptide binding motif interacts with both BIR2
and BIR3 domains of multiple IAPs to release the binding of IAPs to
both initiator (caspase-9) and effector (caspase-3/7) caspases
(16–18), we synthesized a cell permeable Smac
peptide containing the N-terminal 7 amino acids of Smac and a
Drosophila antennaaedia penetration sequence. The Smac N7
peptide appeared to have excellent cellular uptake and the amount
of cytoplasmic accumulated Smac N7 in most cells peaked at 12 h
after addition, as demonstrated by FCM and western blot analysis
after 12 and 24 h of exposure.
TRAIL, a member of the TNF family, has been proposed
as a cancer-specific therapeutic agent, for its rapid
apoptosis-inducing effect and non-toxic effects on normal tissues
when systematically administered (19). Nevertheless, although TRAIL is well
tolerated in patients, its single-agent efficacy is very limited
and not all tumor cell lines respond well (20,21).
Paclitaxel has emerged as the first-line therapy for EOC or
recurrent EOC, however, its clinical application is usually limited
by the dose-dependent toxicity and development of drug resistance
(22,23). Therefore, combination chemotherapy
is usually required to achieve efficacy at tolerable dosages.
Herein, we demonstrated that even the A2780 cell line, which
revealed resistance to TRAIL as shown by our previous study
(15), could be strongly sensitized
by the addition of Smac N7 to TRAIL treatment in a dose-dependent
manner. Co-treatment with Smac N7 had the ability to potentiate the
cytotoxicity of paclitaxel, which offers the potential to broaden
the efficacy of previously established therapies in EOC.
However, we found that as a single agent, Smac N7
did not exhibit cytotoxicity. The non-cytotoxic effect induced by
Smac N7 alone may be explained by two reasons. First, as
demonstrated by western blot analysis, the expression levels of
XIAP and survivin protein in A2780 cells are extremely high, which
suggests that the Smac N7 peptide might not be sufficient to
inhibit their antiapoptotic function. Second, recent studies have
revealed that endogenous Smac protein is rapidly degraded after
being released from mitochondria by the proteasome, and XIAP
functions as an E3 ligase and promotes the degradation of Smac
during apoptosis (24,25). Thus, it could be considered that
XIAP promotes the degradation of synthetic Smac N7 peptide in A2780
cells.
Based on the response observed in vitro, we
corroborated these findings in vivo. Short Smac peptides
fused to a carrier peptide for intracellular delivery were shown to
overcome the resistance of cancer cells with high levels of IAPs
and to enhance the activity of anticancer drugs in a human glioma
xenograft model in vivo(26). Consistent with this study, our
results revealed that in the A2780 xenograft models, intratumoral
injection of the Smac N7 peptide combined with paclitaxel or TRAIL,
caused an additional decrease in tumor load compared to each agent
alone. The toxicity and side effects of antitumor drugs are always
the focus of great concern to oncologists. In our study, the Smac
N7 peptide exhibited no toxicity to vital organs, such as the
heart, kidney and liver of nude mice; TRAIL showed very slight
detectable damage, whereas the mice treated with paclitaxel showed
poor general conditions, such as the decrease in immune function,
and obvious damage to the kidney and liver. Notably, in the
combination treatment groups, administration of Smac N7 did not
reverse the lack of toxicity of TRAIL, and significantly reduced
the toxicity of paclitaxel. In addition, except for the PBS-treated
mice, the spleen of all tested mice displayed an enlarged volume
with blood stasis and infiltrating mononuclear phagocytes. It could
be hypothesized that the immune system is involved in the activity
of the Smac N7 peptide, which was supported by a recently published
report that a Smac mimetic elicited a proinflammatory cell death
that was sufficient to activate adaptive antitumor immune responses
in cancer (27).
We next explored whether the growth inhibition of
A2780 cell xenografts after combination therapy was in part due to
induction of apoptosis. Detection of apoptotic cells by
fluorometric TUNEL assay demonstrated that Smac N7 alone did not
induce apoptosis, while paclitaxel or TRAIL moderately induced
apoptosis while the combination of Smac N7 and paclitaxel or TRAIL
showed additive effects. The accumulation of cleaved PARP and
caspase-3 in the treated A2780 xenograft tumor cells further
confirmed the induction of the apoptotic process.
Concerning the mechanism of action of the Smac N7
peptide, several recent studies showed that this peptide enhanced
the apoptosis-inducing effect of cytotoxic drugs through the rapid
degradation of XIAP and survivin (28,29).
In the present study, although the level of XIAP protein in A2780
tumor xenografts was slightly downregulated and the survivin
protein level was not affected by Smac N7 alone, their expression
levels were significantly downregulated following the combination
treatment with paclitaxel or TRAIL. We hypothesized that this
result was probably associated with the ability to reduce the
protein level of XIAP by paclitaxel or TRAIL, which relieved the E3
ligase effect of XIAP on the degradation of Smac protein.
Additionally, it should be noted that Bcl-2 expression remained
unchanged in the Smac N7, TRAIL or the combination-treated A2780
tumor models. Paclitaxel reduced the expression of Bcl-2, but the
co-treatment with Smac N7 did not have an additive effect.
Collectively, these data provide strong evidence that XIAP and
survivin are the primary cellular targets for Smac N7 in its
synergistic interaction with paclitaxel or TRAIL.
In summary, our data showed that the combination
therapy of paclitaxel or TRAIL in conjugation with the Smac N7
peptide offers a promising strategy for the treatment of human
epithelial ovarian cancer. Administration of Smac N7 did not
reverse the lack of toxicity of TRAIL for normal tissues, and the
combination treatments of Smac N7 and paclitaxel allowed a
reduction in the therapeutically active dose of paclitaxel for EOC.
These effects are, at least in part, mediated by the reduction in
XIAP and survivin expression. Therefore, the combination of the
Smac N7 peptide with TRAIL or paclitaxel presents a promising
strategy for apoptosis-targeted therapies of EOC and warrants
further investigation.
References
|
1
|
Lee JY, Shin JY, Kim HS, et al: Effect of
combined treatment with progesterone and tamoxifen on the growth
and apoptosis of human ovarian cancer cells. Oncol Rep. 27:87–93.
2012.PubMed/NCBI
|
|
2
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev. 5:876–885. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sayers TJ: Targeting the extrinsic
apoptosis signaling pathway for cancer therapy. Cancer Immunol
Immunother. 60:1173–1180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qiu W, Liu H, Sebastini A, Sun Q, Wang H,
Zhang L and Yu J: An apoptosis-independent role of SMAC in tumor
suppression. Oncogene. Jul 2–2012.(Epub ahead of print).
|
|
5
|
Fulda S and Vucic D: Targeting IAP
proteins for therapeutic intervention in cancer. Nat Rev Drug
Discov. 11:109–124. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Almagro MC and Vucic D: The inhibitor
of apoptosis (IAP) proteins are critical regulators of signaling
pathways and targets for anti-cancer therapy. Exp Oncol.
34:200–211. 2012.PubMed/NCBI
|
|
7
|
Chai J, Du C, Wu JW, Kyin S, Wang X and
Shi Y: Structural and biochemical basis of apoptotic activation by
Smac/DIABLO. Nature. 406:855–862. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du C, Fang M, Li Y, Li L and Wang X: Smac,
a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Z, Sun C, Olejniczak ET, et al:
Structural basis for binding of Smac/DIABLO to the XIAP BIR3
domain. Nature. 408:1004–1008. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arnt CR, Chiorean MV, Heldebrant MP, Gores
GJ and Kaufmann SH: Synthetic Smac/DIABLO peptides enhance the
effects of chemotherapeutic agents by binding XIAP and cIAP1 in
situ. J Biol Chem. 277:44236–44243. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Giagkousiklidis S, Vogler M, Westhoff MA,
Kasperczyk H, Debatin KM and Fulda S: Sensitization for
gamma-irradiation-induced apoptosis by second mitochondria-derived
activator of caspase. Cancer Res. 65:10502–10513. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rein DT, Volkmer AK, Volkmer J, et al:
Systemic administration of bevacizumab prolongs survival in an
in vivo model of platinum pre-treated ovarian cancer. Oncol
Lett. 3:530–534. 2012.PubMed/NCBI
|
|
13
|
Krasner CN, Poveda A, Herzog TJ, et al:
Patient-reported outcomes in relapsed ovarian cancer: Results from
a randomized Phase III study of trabectedin with pegylated
liposomal doxorubicin (PLD) versus PLD Alone. Gynecol Oncol.
127:161–167. 2012. View Article : Google Scholar
|
|
14
|
Mulier S, Claes JP, Dierieck V, et al:
Survival benefit of adding Hyperthermic IntraPEritoneal
Chemotherapy (HIPEC) at the different time-points of treatment of
ovarian cancer: review of evidence. Curr Pharm Des. 18:3793–3803.
2012. View Article : Google Scholar
|
|
15
|
Mao HL, Liu PS, Zheng JF, Zhang PH, Zhou
LG, Xin G and Liu C: Transfection of Smac/DIABLO sensitizes
drug-resistant tumor cells to TRAIL or paclitaxel-induced apoptosis
in vitro. Pharmacol Res. 56:483–492. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oost TK, Sun C, Armstrong RC, et al:
Discovery of potent antagonists of the antiapoptotic protein XIAP
for the treatment of cancer. J Med Chem. 47:4417–4426. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bianchi A, Ugazzi M, Ferrante L, Lecis D,
Scavullo C, Mastrangelo E and Seneci P: Rational design, synthesis
and characterization of potent, drug-like monomeric Smac mimetics
as pro-apoptotic anticancer agents. Bioorg Med Chem Lett.
22:2204–2208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang D, Zhao Y, Li AY, Wang S, Wang G and
Sun Y: Smac-mimetic compound SM-164 induces radiosensitization in
breast cancer cells through activation of caspases and induction of
apoptosis. Breast Cancer Res Treat. 133:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Voelkel-Johnson C: TRAIL-mediated
signaling in prostate, bladder and renal cancer. Nat Rev Urol.
8:417–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kauntz H, Bousserouel S, Gossé F and Raul
F: The flavonolignan silibinin potentiates TRAIL-induced apoptosis
in human colon adenocarcinoma and in derived TRAIL-resistant
metastatic cells. Apoptosis. 17:797–809. 2012. View Article : Google Scholar
|
|
21
|
Fandy TE, Shankar S and Srivastava RK:
Smac/DIABLO enhances the therapeutic potential of chemotherapeutic
drugs and irradiation, and sensitizes TRAIL-resistant breast cancer
cells. Mol Cancer. 7:602008. View Article : Google Scholar
|
|
22
|
Boere IA and van der Burg ME: Review of
dose-intense platinum and/or paclitaxel containing chemotherapy in
advanced and recurrent epithelial ovarian cancer. Curr Pharm Des.
8:3741–3753. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lindemann K, Christensen RD and Vergote I:
First-line treatment of advanced ovarian cancer with
paclitaxel/carboplatin with or without epirubicin (TEC versus TC) -
a gynecologic cancer intergroup study of the NSGO, EORTC GCG and
NCIC CTG. Ann Oncol. 23:2613–2619. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MacFarlane M, Merrison W, Bratton SB and
Cohen GM: Proteasome-mediated degradation of Smac during apoptosis:
XIAP promotes Smac ubiquitination in vitro. J Biol Chem.
277:36611–36616. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ma L, Huang Y, Song Z, et al: Livin
promotes Smac/DIABLO degradation by ubiquitin-proteasome pathway.
Cell Death Differ. 13:2079–2088. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fulda S, Wick W, Weller M and Debatin KM:
Smac agonists sensitize for Apo2L/TRAIL- or anticancer drug-induced
apoptosis and induce regression of malignant glioma in vivo.
Nat Med. 8:808–815. 2002.PubMed/NCBI
|
|
27
|
Emeagi PU, Van Lint S, Goyvaerts C, et al:
Proinflammatory characteristics of SMAC/DIABLO-induced cell death
in antitumor therapy. Cancer Res. 72:1342–1352. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang G, Zhao J, Xiao X, et al: An
N-terminal Smac peptide sensitizes human prostate carcinoma cells
to methyl jasmonate-induced apoptosis. Cancer Lett. 302:37–46.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiao XY, Jiang GS, Wang L, Lv L and Zeng
FQ: Predominant enhancement of apoptosis induced by methyl
jasmonate in bladder cancer cells: therapeutic effect of the
Antp-conjugated Smac peptide. Anticancer Drugs. 22:853–863. 2011.
View Article : Google Scholar : PubMed/NCBI
|