Introduction
Breast cancer is a malignant tumor with a high
mortality rate (1), which can be
attributed primarily to invasion and metastasis. One of the primary
approaches to treating breast cancer metastasis has been the
development of effective anti-invasive agents (2,3). The
initial steps of metastasis include cellular invasion through the
degradation of the extracellular matrix (ECM), followed by the
migration of cancer cells to other organs through the surrounding
tissues (4,5). The ECM comprises collagens, laminins,
glycoproteins, proteoglycans/glycosaminoglycans and fibronectin
(6). It is degraded by
extracellular proteases, of which MMPs play an important role in
breast cancer (4,5).
Matriptases are members of the type II transmembrane
serine protease (TTSP) family and are expressed in the epithelial
compartments of all tissue types (7). Matriptase dysregulation is involved
in various epithelial carcinomas, such as breast, prostate, colon,
ovarian, uterine, cervical and skin cancers, where it is reportedly
upregulated (8,9). Matriptase was first discovered in
breast cancer cell lines, where it is highly expressed (10,11);
however, despite its importance in breast cancer (12,13),
the mechanism underlying its effects on breast cancer metastasis is
unclear.
Protease-activated receptor-2 (PAR-2), a G
protein-coupled receptor 11, induces various intracellular
signaling pathways by activating endogenous serine proteinases,
including matriptase (14–17). Previous studies have shown that
matriptases are important activators of PAR-2 (16,18).
When PAR-2 binds to a G protein, it produces diacylglycerol (DAG)
and activates canonical phospholipase C
(PLC)/Ca2+/protein kinase C (PKC) signaling or
extracellular signal-regulated kinase-1/2 (19–21).
Furthermore, PAR-2 levels are elevated in breast cancer, which
plays a key role in regulating cellular migration by MAPKs
(22,23).
MMPs are a family of zinc-dependent endopeptidases
that consist of six subclasses: Collagenases, stromelysins,
gelatinases, matrilysins, membrane-associated MMP and other MMPs
(24). MMP-9 is involved in cancer
cell infiltration and is directly associated with poor patient
prognosis and the metastasis of breast cancer (25,26).
Therefore, the regulation of signaling pathways to inhibit MMP-9
expression may play an important role in the treatment of various
malignancies, including breast cancer (27–31).
The expression of MMP-9 is induced by various stimuli, including
cytokines, growth factors and
12-O-tetradecanoylphorbol-13-acetate (TPA) [10] (32–36).
In particular, TPA is known to stimulate MMP expression by
activating PKC in breast cancer cells (27,28,33).
Furthermore, several studies have indicated that TPA activates PKC
by activating PLC. In breast cancer invasion, TPA-induced MMP-9
expression is known to be induced by activation of NF-κB and
activator protein-1 (AP-1) (37,38),
transcription factors whose expression is regulated by MAPKs
(39,40).
In the present study, the regulatory role of
matriptase in TPA-induced MMP-9 expression, as well as invasion and
migration, were investigated using MCF-7 breast cancer cells.
Furthermore, to confirm the signaling mechanism of matriptase, the
association between PAR2 and PLC/PKC was investigated. These
results may provide a potential strategy for the treatment of
breast cancer metastasis.
Materials and methods
Cell lines and culture
The human MCF-7 breast cancer cell line was
purchased from the American Type Culture Collection. The cells were
cultured in high-glucose Dulbecco's Modified Eagle's Medium (DMEM)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.) and 1% antibiotics (antibiotic-antimycotic, 100X; Gibco;
Thermo Fisher Scientific, Inc.), and maintained in a humidified
incubator at 37°C (5% CO2).
Reagents
TPA (cat. no. P1585) and DMSO were obtained from
Sigma-Aldrich (Merck KGaA). Matrigel was acquired from Corning,
Inc. The PAR-2 antagonist (GB83) was purchased from Axon Medchem
LLC, and BAPTA-AM was obtained from Invitrogen (Thermo Fisher
Scientific, Inc.).
Western blot analysis
MCF-7 cells (5×107) were transfected with
matriptase siRNA for 24 h. Additionally, cells (7×105)
were treated with BAPTA-AM or GB83 for 1 h, and then incubated with
TPA for 24 h at 37°C. Total protein was extracted from cells using
RIPA lysis buffer (Thermo Fisher Scientific, Inc.) containing
protease and phosphatase inhibitors (Calbiochem; Merck KGaA). The
lysates were centrifuged at 16,000 × g for 10 min at 4°C, and the
protein concentrations were evaluated using the BioSpec-nano
spectrophotometer (Shimadzu Corporation). The samples (20 µg) were
separated by 10% SDS-PAGE and then transferred to Hybond™
polyvinylidene fluoride membranes (Cytiva). The membranes were
blocked with 5% BSA (bovine serum albumin) or 5% skim milk buffers
for 2 h at 4°C, and then incubated with the following primary
antibodies (all 1:2,500) overnight at 4°C: Anti-β-actin (cat. no.
A5441; Sigma-Aldrich; Merck KGaA); JNK (cat. no. 9252), p38 (cat.
no. 9212), ERK (cat. no. 9102), IκB kinase α (IKKα; cat. no. 2682),
IKKβ (cat. no. 2678), phosphorylated forms of PLCγ2 (cat. no.
3874), JNK (cat. no. 9251), p38 (cat. no. 9211), ERK (cat. no.
9101), c-Jun, IκBα (cat. no. 2859) and IKKαβ (cat. no. 2697) (all
Cell Signaling Technology, Inc.). PLCγ2 (cat. no. SC-5283), p50
(cat. no. SC-7178), IκBα (cat. no. SC-371), MMP-9 (cat. no.
SC-12759) and proliferating cell nuclear antigen (cat. no. SC-7907)
(all Santa Cruz Biotechnology, Inc.). PKCα (cat. no. ab32376), PKCβ
(cat. no. ab32026), PKCδ (cat. no. ab182126) and anti-sodium ATPase
plasma membrane loading control (cat. no. ab76020) (all Abcam).
Matriptase-specific antibodies were obtained from R&D Systems
(cat. no. MAB3946). The blots were washed in TBS with 0.2% Tween-20
and then incubated with secondary HRP (horseradish
peroxidase)-conjugated anti-mouse (cat. no. SC-2005) or anti-rabbit
(cat. no. SC-2004) antibodies (1:2,500; both Santa Cruz
Biotechnology, Inc.) for 1 h at 4°C. Immunoreactive bands were
detected using Luminol HRP Substrate Reagent (EMD Millipore) with a
Mini HD6 Image Analyzer and Alliance 1D (UVItec Cambridge; Cleaver
Scientific Ltd.). Immunoreactive bands were quantified using ImageJ
software (Version 1.53k; National Institutes of Health).
RNA isolation and reverse
transcription-quantitative (RT-q) PCR
RT-qPCR was performed using the StepOnePlus™
Real-time PCR System and SYBR-Green PCR Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Total RNA was isolated
from MCF-7 cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. RNA concentration and purity were determined by
absorbance at 260/280 nm. Complementary DNA was synthesized from 1
µg total RNA using the PrimeScript™ RT Reagent Kit (Takara Bio,
Inc.) according to the manufacturer's instructions. The primers
were: MMP-9 forward, 5′-CCTGGAGACCTGAGAACCAATCT-3′ and reverse,
5′-CCACCCGAGTGTAACCATAGC-3′; and GAPDH forward,
5′-ATGGAAATCCCATCACCATCTT-3′ and reverse, 5′-CGCCCCACTTGATTTTGG-3′.
mRNA expression levels were normalized to those of GAPDH. The qPCR
cycling conditions were as follows: Initial denaturation at 95°C
for 10 min, 40 cycles of 95°C for 15 sec and 60°C for 1 min,
followed by a melting curve ranging from 95°C for 15 sec, 60°C for
1 min, to 95°C for 15 sec. Relative quantitation was performed
using the comparative 2−∆∆Cq method (41).
Small interfering RNA (siRNA)
transfection and preparation of cytosolic and nuclear protein
extracts
MCF-7 cells were transfected with 100 pmol
matriptase siRNA or negative control siRNA (Shanghai GenePharma
Co., Ltd.) using Lipofectamine® RNAiMAX reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) for 24 h at 37°C (5%
CO2), and then incubated with TPA for 3 h at 37°C. The
sequences of human siRNA were as follows: Matriptase siRNA,
5′-GUGUCCAGAAGGUCUUCAATT-3′ (sense) and 5′-UUGAAGACCUUCUGGACACTT
(antisense); control siRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′ (sense) and
5′-ACGUGACACGUUCGGAGAATT-3′ (antisense). Cytoplasmic and nuclear
extracts were prepared from the cells using NE-PER Nuclear and
Cytoplasmic Extraction Reagents (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol.
Dual-luciferase reporter assay
Cells transfected with matriptase siRNA were then
transfected with the NF-κB/AP-1 luciferase reporter plasmid
(Agilent Technologies, Inc.) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. At 24 h post transfection, the cells
were treated with 20 nM TPA for 4 h at 37°C. Whole-cell lysates
were prepared, and luciferase activity was measured using the
Dual-luciferase Reporter Assay Kit (Promega Corporation) and Lumat
LB 9507 Luminometer (Berthold Technologies GmbH & Co.KG).
Relative Firefly luciferase activity was normalized to
Renilla luciferase activity.
Membrane fractionation
MCF-7 cells (5×107) were transfected with
matriptase siRNA for 24 h. Additionally, 7×105 cells
were treated with BAPTA-AM or GB83 for 1 h and then incubated with
TPA for 1 h at 37°C. The cells were mixed with homogenization
buffer (20 mM Tris-HCl, 2 mM EDTA, 5 mM EGTA, 5 mM DTT and protease
inhibitor; pH 7.5) and homogenized using a sonicator (5 times for
10 sec, each at 10% amplitude) and incubated on ice for 30 min. To
separate the soluble (cytosolic) and pellet (membrane) fractions,
the cell lysate was centrifuged at 16,000 × g for 15 min at 4°C.
The pellet fraction was incubated in a solubilization buffer
(homogenization buffer containing 1% NP-40) for 30 min on ice, and
then centrifuged at 16,000 × g for 15 min at 4°C.
Cellular invasion and migration
assays
The invasion assay was carried out in 24-well
chambers (pore size, 8 µm) coated with 20 µl Matrigel (diluted in
DMEM) for 30 min at 37°C; Matrigel Basement Membrane Matrix
(Corning, Inc.) was rehydrated in 0.5 ml DMEM for 2 h immediately
prior to experimentation. The top chamber was seeded with medium
(0.5 ml; 10% FBS and 1% antibiotics) with 3×105
resuspended cells transfected with matriptase siRNA, while the
lower chamber was filled with medium containing TPA alone or
combined with GB83. A migration assay was performed using chambers
without Matrigel. Cells transfected with control and matriptase
siRNA were added to the upper chamber and medium with TPA alone or
with GB83 was added to the bottom chamber. Cells were allowed to
invade/migrate to the lower membrane for 24 h (37°C). After
incubation, the cells on the upper membrane surface were removed
with cotton swabs. The migrated/invasive cells were fixed with
formaldehyde solution (3.6%) for 10 min, stained with crystal
violet for 20 min (both at room temperature), and counted in five
random fields per chamber at ×10 magnification, using a Leica DM
ILLED inverted microscope (Leica Microsystems).
Statistical analysis
Data are presented as the mean ± SD of ≥3
independent experiments. Statistical analysis was performed using
ANOVA with Scheffe's post hoc test (SAS software, version 9.3: SAS
Institute Inc.), and P<0.05 was considered to indicate a
statistically significant difference.
Results
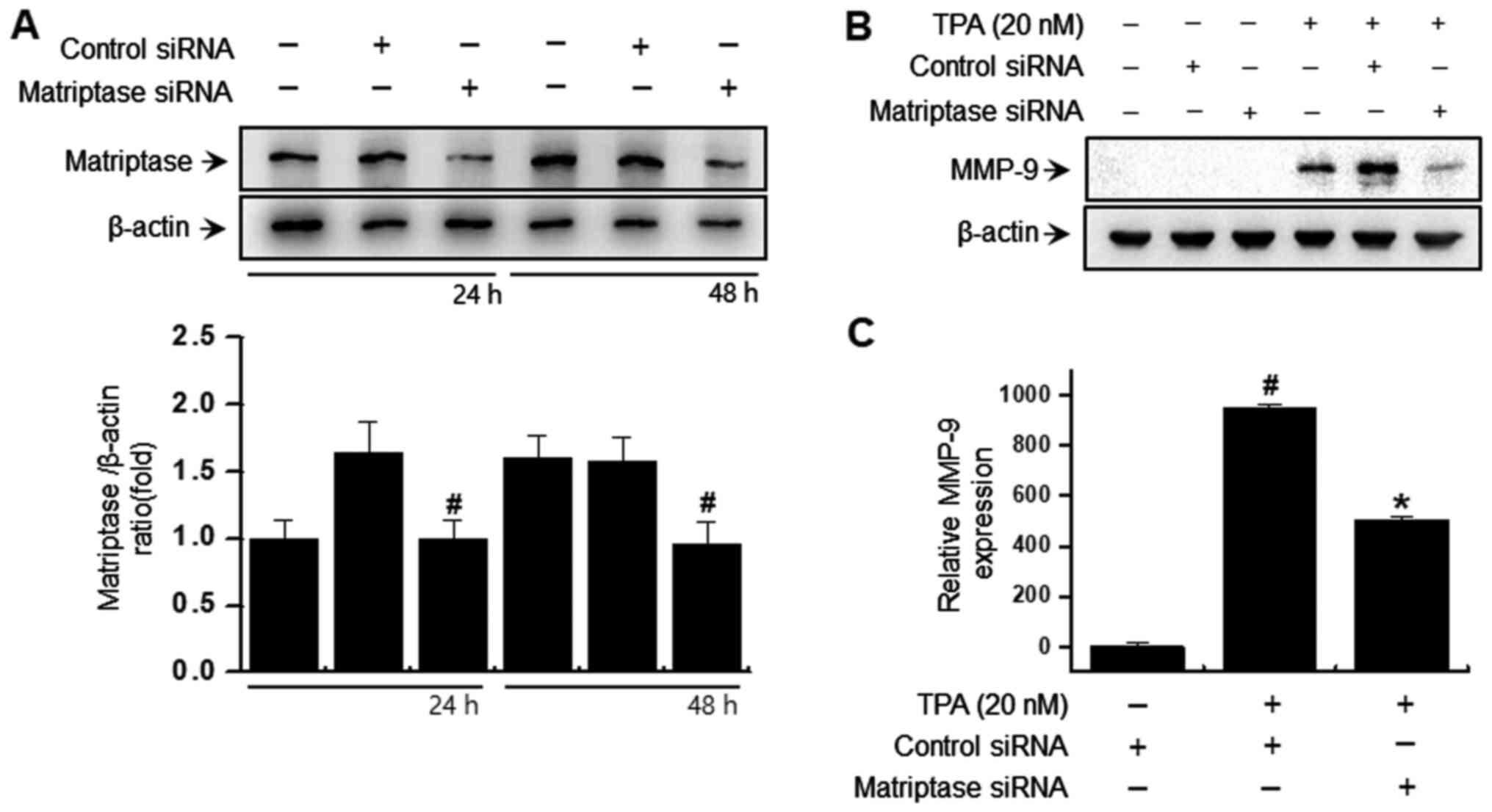
Downregulation of matriptase
suppresses TPA-induced MMP-9 expression in MCF-7 breast cancer
cells
Western blotting and RT-qPCR were used to determine
the effect of matriptase on TPA-induced MMP-9 expression.
Intracellular matriptase expression was suppressed by transfection
with matriptase siRNA (Fig. 1A),
which inhibited the protein/mRNA levels of TPA-induced MMP-9
(Fig. 1B and C). These results
suggested that matriptase was involved in TPA-induced MMP-9
expression.
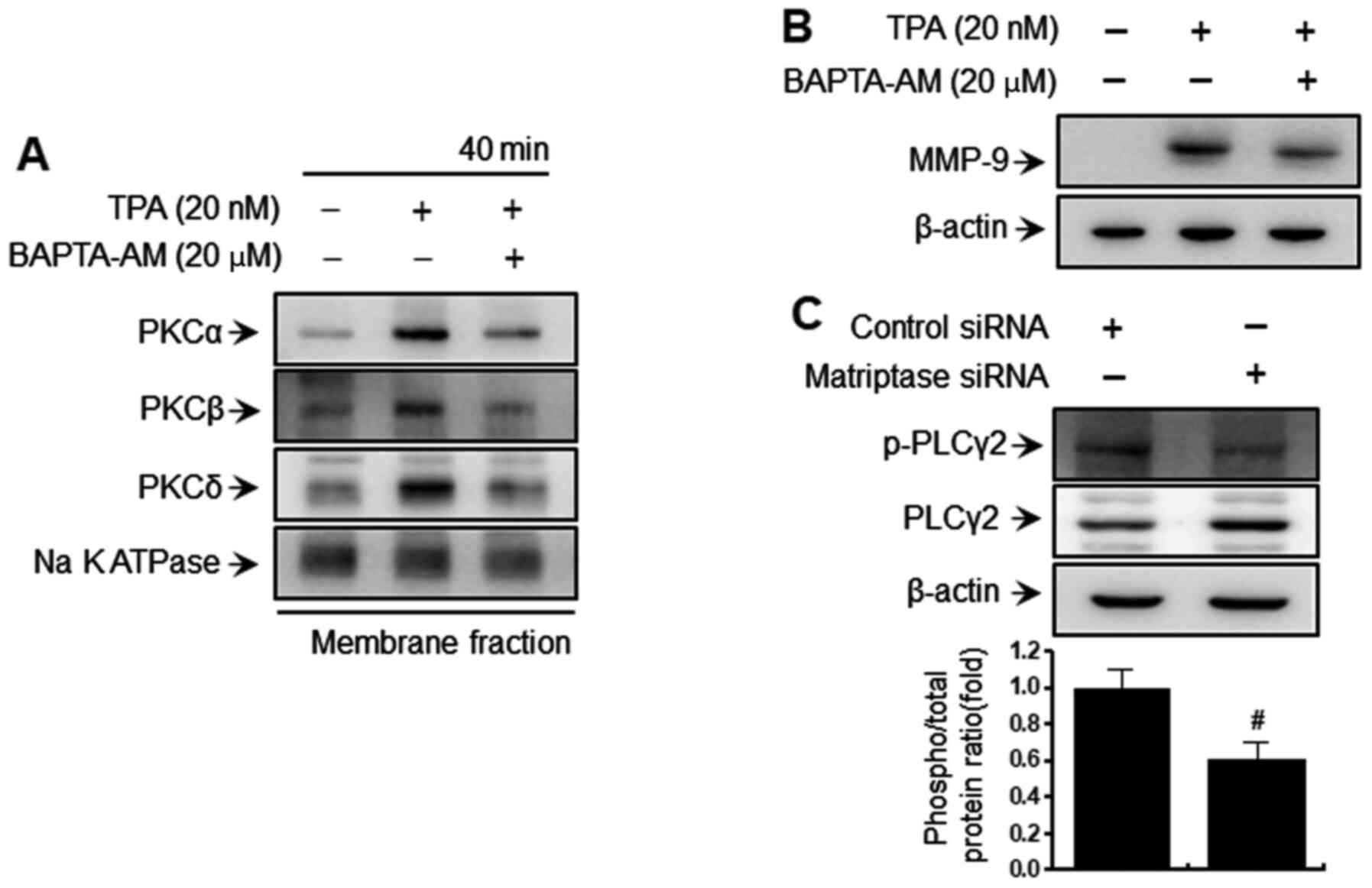
Downregulation of matriptase reduces
PLCγ2 phosphorylation in MCF-7 breast cancer cells
Activation of PKC isozymes is mediated by DAG and
Ca2+ (42). Therefore,
intracellular calcium levels are important for MMP-9 expression and
cellular metastasis through TPA-mediated PKC activation. In the
present study, it was confirmed that an intracellular calcium
chelator (BAPTA-AM) inhibited TPA-induced PKC activation (Fig. 2A) and MMP-9 expression (Fig. 2B). In addition,
matriptase-knockdown suppressed PLCγ2 phosphorylation (Fig. 2C) in MCF-7 cells. These findings
indicated that intracellular calcium levels are important for
PKC-mediated MMP-9 expression, and that matriptase downregulation
inhibited MMP expression and metastatic ability by regulating
PLCγ2-mediated intracellular calcium levels.
Matriptase regulates TPA-induced PKC
activation, as well as the MAPK and IKK signaling pathways in MCF-7
breast cancer cells
Previous studies have shown that PKC and the MAPK
and IKK signaling pathways are involved in the expression of MMP-9
induced by TPA (38). In the
present study, the effects of matriptase on PKC and the MAPK and
IKK signaling pathways were confirmed. To determine whether
matriptase affects PKC activation in TPA-induced MCF-7 breast
cancer cells, membrane translocation levels of PKCα, PKCβ and PKCδ
were evaluated. As shown in Fig.
3A, matriptase-knockdown attenuated TPA-mediated PKC membrane
translocation. Furthermore, matriptase-knockdown reduced MAPK
phosphorylation (p38, ERK and JNK) at 30 min post-TPA treatment
(Fig. 3B), confirming the effect
of matriptase on MAPK activation by TPA. In addition,
matriptase-knockdown suppressed p-IKKαβ and p-IκBα levels and the
degradation of IκBα in the cytoplasmic fraction, which confirms the
role of matriptase on the NF-κB signal transduction cascade
(Fig. 3C). These findings
suggested that matriptase is involved in the activation of PKC and
the MAPK and IKK signaling pathways through TPA-induced expression
of MMP-9 in MCF-7 breast cancer cells.
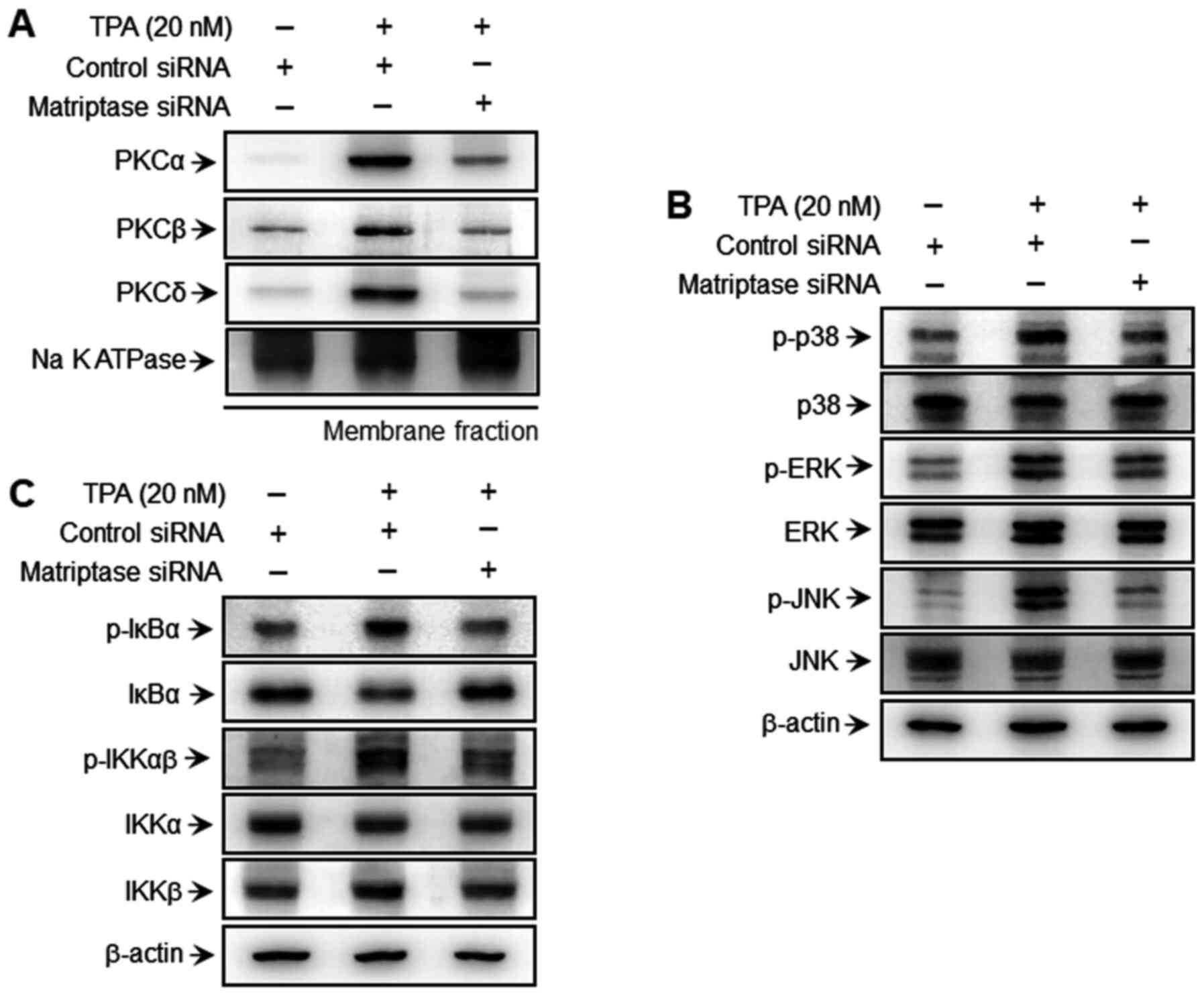 | Figure 3.Effect of matriptase on TPA-induced
PKC activation and MAPK signaling in MCF-7 cells. (A) MCF-7 cells
were transfected with control and matriptase siRNAs for 24 h,
followed by incubation with 20 nM TPA for 40 min. PKC isozyme
levels in cell membrane fractions were analyzed by western
blotting. (B) Transfected cells were treated with 20 nM TPA for 30
min and cell lysates were prepared for western blotting to evaluate
the MAPK signaling pathway. (C) Additionally, transfected cells
were treated with TPA, and after 4 h of incubation, cytoplasmic
lysates were prepared; expression of upstream signaling molecules
NF-κB, p-IκBα, IκBα, p-IKKαβ, IKKα and IKKβ were than analyzed via
western blotting. TPA, 12-O-tetradecanoylphorbol-13-acetate;
PKC, protein kinase C; siRNA, small interfering RNA; p-,
phosphorylated; Na K ATPase, sodium/potassium ATPase. |
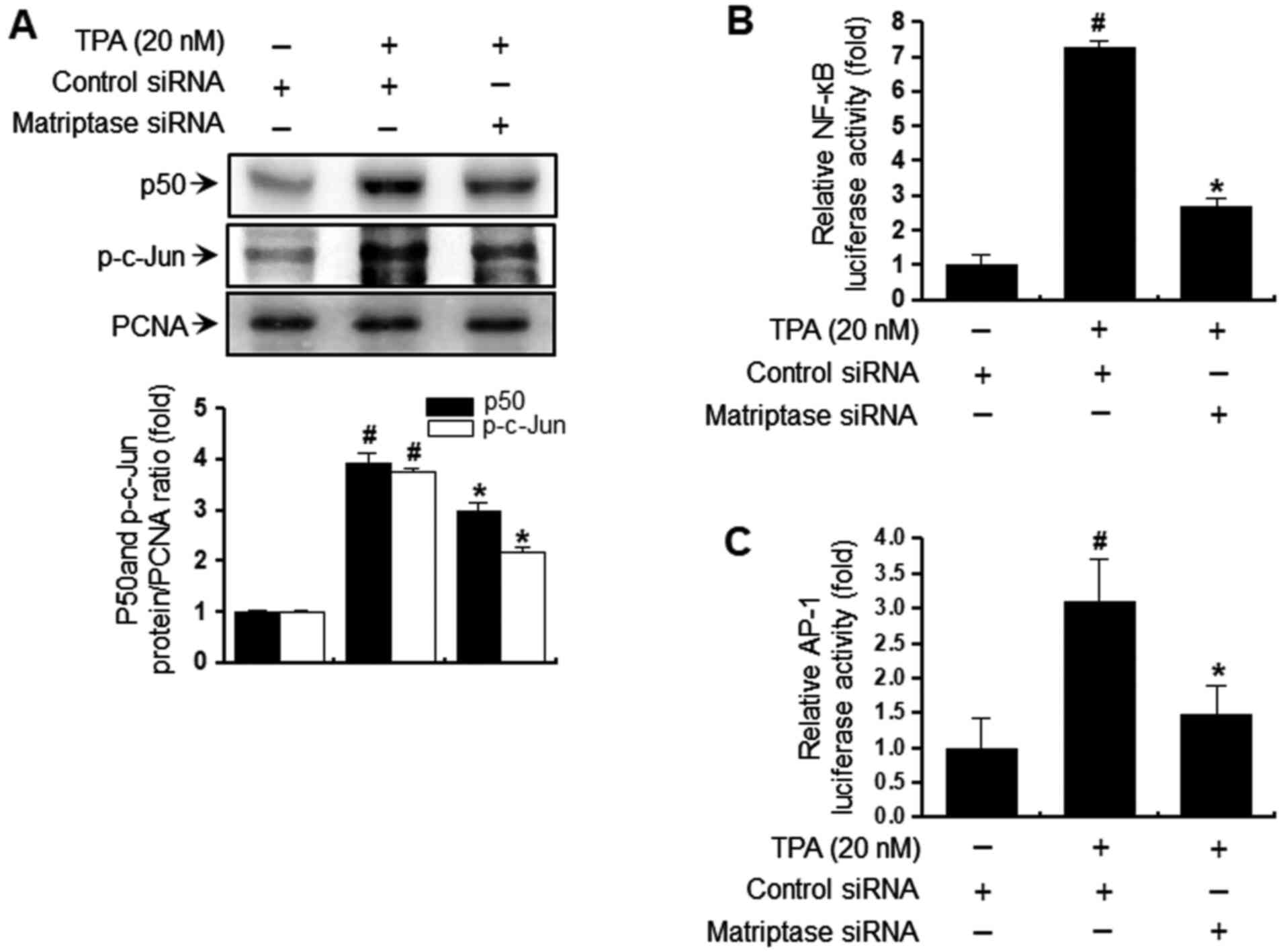
Matriptase-knockdown decreases
TPA-mediated activation of NF-κB and AP-1 in MCF-7 breast cancer
cells
To elucidate the mechanism by which matriptase
inhibits TPA-induced MMP-9 expression, the effect of matriptase
siRNA on TPA-induced NF-κB and AP-1 activation was evaluated using
a luciferase reporter assay. First, western blot analysis was used
to confirm that matriptase-knockdown suppressed p50 levels in the
nuclear fraction (Fig. 4A).
Furthermore, TPA induced the phosphorylation of c-Jun, a major
subunit of AP-1, and matriptase-knockdown inhibited the
phosphorylation of c-Jun (Fig.
4A). Also, matriptase siRNA treatment inhibited TPA-stimulated
NF-κB/AP-1 binding in a luciferase assay (Fig. 4B and C). These findings demonstrate
that matriptase regulated the expression of MMP-9 induced by TPA
through the NF-κB and AP-1 pathways in MCF-7 breast cancer
cells.
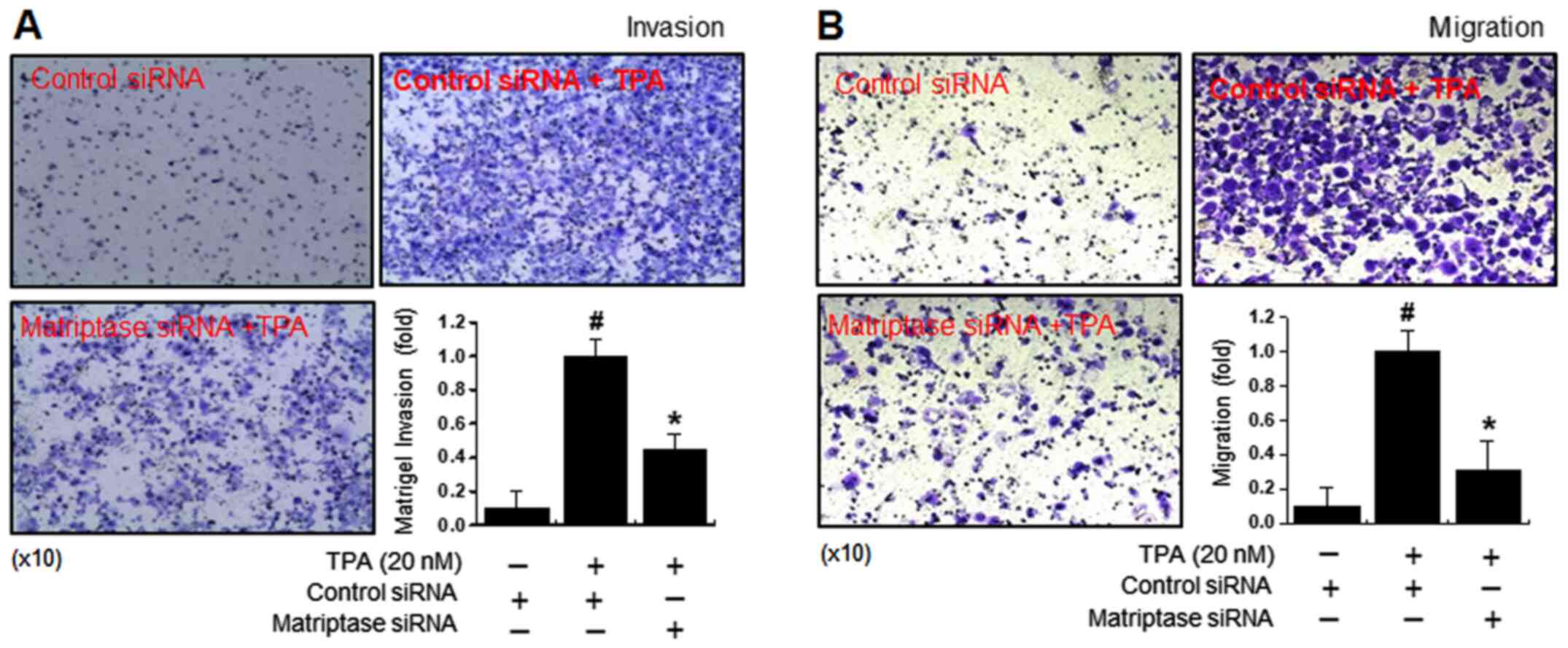
Matriptase-knockdown inhibits
TPA-mediated migration and invasiveness of MCF-7 breast cancer
cells
In previous study, upregulation of MMP-9 has been
associated with the induction of cancer cell metastasis, including
breast cancer (43). Therefore,
the inhibitory effect of matriptase siRNA on the metastatic
efficacy of MCF-7 cells was investigated using invasion (Fig. 5A) and migration (Fig. 5B) assays. TPA-induced invasiveness
and migration were significantly reduced in cells treated with
matriptase siRNA, compared with control siRNA- and TPA-treated
cells.
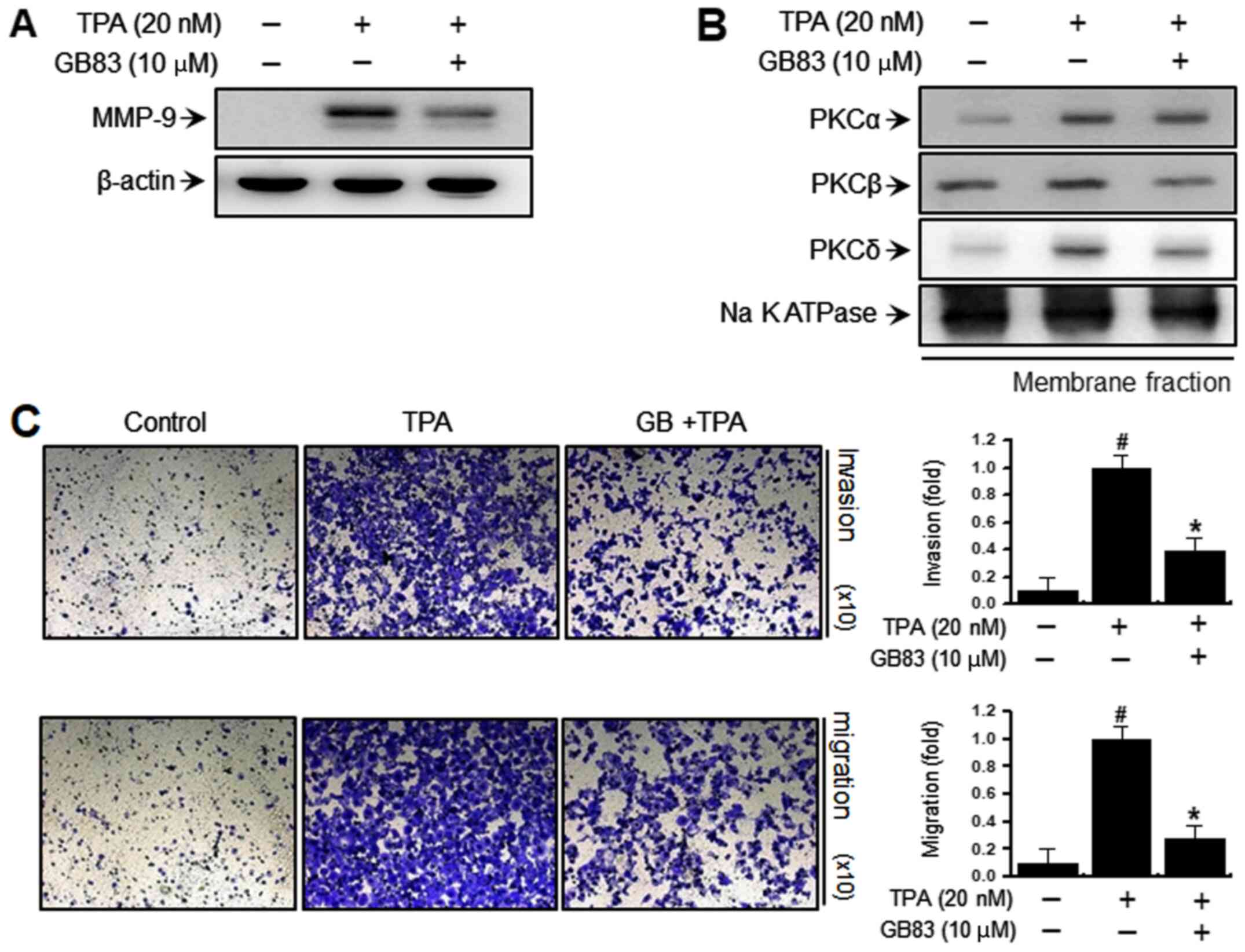
Inhibition of PAR-2 suppresses
TPA-induced PKC activation, MMP expression, as well as invasiveness
and migration, in MCF-7 breast cancer cells
Matriptase induces PAR-2 activation (15); therefore, to investigate the effect
of PAR-2-mediated breast cancer invasiveness, the effect of a PAR-2
inhibitor (GB83) on PKC activation and MMP-9 expression was
evaluated in TPA-treated MCF-7 cells. GB83 (10 µM) was found to
inhibit the expression of TPA-induced MMP-9 (Fig. 6A). It also attenuated TPA-mediated
translocation of PKC to the membrane (Fig. 6B). Furthermore, invasion and
migration assays revealed the inhibitory effect of GB83 on the
metastatic properties of MCF-7 cells (Fig. 6C). These results suggested that
PAR-2 was involved in PKC-mediated MMP-9 expression and metastatic
ability in MCF-7 breast cancer cells.
Discussion
Breast cancer is a malignant tumor and the leading
cause of mortality in women worldwide (2). The majority of breast cancer deaths
result from metastasis to the bone, lung, liver, brain and kidney
(1). The molecular mechanisms
underlying cancer cell invasiveness and migration are complex; the
initial event that provides biochemical and mechanical barriers to
cancer cell migration is the proteolytic degradation of the ECM
(4,44), which requires the activation and
expression of MMPs, known to play a major role in breast cancer
(43,44). Among the MMPs, MMP-9 activation is
associated with tumor progression and invasion (45,46).
Therefore, inhibition of the regulatory pathway involved in MMP-9
expression may be an important therapeutic strategy for preventing
breast cancer metastasis. In the present study, matriptase was
proposed as a signaling protein for inhibiting cellular metastasis
through the regulation of MMP-9. Matriptase was first reported in
1993 to have novel gelatinolytic activity in breast cancer cells
(47). Matriptase is one of the
most well studied members of the TTSP family and is expressed in
the epithelial compartments of all tissue types (7,48,49),
where its dysregulation is associated with numerous types of cancer
and poor patient outcomes therein (8,9).
Furthermore, several studies have demonstrated that matriptase is
highly expressed in MCF-7 breast cancer cells (10,11);
in particular, matriptase is upregulated in breast cancer and
increases the proliferation and invasiveness of breast cancer cells
(11,50). A previous study demonstrated that
inhibiting matriptase suppresses breast cancer progression using
in vivo, ex vivo and in vitro approaches (50). However, the role and signaling
mechanisms of matriptase in breast cancer metastasis were
previously unclear. Therefore, the aim of the present study was to
identify the regulatory role of matriptase in TPA-induced MMP-9
expression and invasion/migration in MCF-7 breast cancer cells. The
findings show that inhibition of matriptase expression inhibited
TPA-induced increases in MMP-9 expression, cellular invasiveness
and migration (Fig. 1 and 5).
The present study demonstrated the role of
matriptase in breast cancer metastasis by identifying its effects
on MCF-7 breast cancer cell invasiveness, as well as the underlying
mechanisms. Matriptase mediates multiple intracellular signaling
pathways by cleaving the activation site of PAR-2, a G
protein-coupled receptor (16,17).
PAR-2 signaling produces DAG and activates the PKC-mediated NF-κB
signaling pathway (19,22). Furthermore, the binding of PAR-2 to
G protein induces canonical PLC/Ca2+/PKC signaling (19). Moreover, activation of
PAR-2-induced MAPK signaling plays an important role in regulating
the migration of breast cancer cells (22,23).
These findings suggest that the PAR2-mediated signaling pathway is
important in breast cancer cell metastasis. The current study
results confirmed that that inhibition of PAR-2 in MCF-7 cells
suppressed PKC activation, MMP-9 expression and cellular
invasiveness (Fig. 6). In
addition, inhibition of matriptase regulated MMP-9 expression and
invasiveness mediated by Par-2/PLCγ2/PKC or Par-2/MAPK.
The activation of PKC is highly associated with
increased invasiveness in breast cancer (51). TPA increases the invasiveness of
breast cancer cells by activating MMP-9 through PKC (52,53).
TPA also activates novel (δ, ε, η and θ) and conventional (α, βI,
βII and γ) PKC isozymes by binding the C1 domains of these isoforms
(54). The effect of TPA is
similar to that of DAG, a natural activator of the PKC isoform.
TPA-mediated activation of PKC involves the translocation of PKC
isoforms to the plasma membrane, resulting in modulation of gene
expression, proliferation, apoptosis, differentiation and malignant
transformation of cancer cells (54,55).
PLCγ2 is a member of the phosphoinositide-specific
PLCs and enhances PKC activation by catalyzing the degradation of
phosphatidylinositol-4,5-bisphosphate in DAG and
inositol-3,4,5-trisphosphate (IP3). IP3
induces an increase in intracellular calcium levels (42,55,56).
A variety of cell signaling pathways act downstream of PKC
isozymes, such as those of Ras/Raf/MAPK, PI3K/Akt and the
transcription factors NF-κB, AP-1 and STAT-3 (57).
Our previous study demonstrated that the activation
of PKCα, PKCβ and PKCδ by TPA mediates the expression and secretion
of MMP-9 (58). Therefore, the
current study confirmed that intracellular calcium is required for
TPA-induced PKC activation and MMP-9 expression and invasiveness
(Fig. 2A and B). In addition,
inhibiting matriptase expression was found to reduce the expression
of p-PLCγ2 (Fig. 2C). Furthermore,
the study revealed that inhibition of matriptase expression reduced
the TPA-induced membrane localization of PKCα, PKCβ and PKCδ in
MCF-7 cells (Fig. 3A). These
findings indicate that inhibiting matriptase expression modulates
PKC-mediated MMP-9 expression and metastasis in MCF-7 breast cancer
cells by inhibiting PLCγ2 activation, and regulating PLCγ2-mediated
calcium levels.
To investigate the TPA-induced PKC downstream
signaling cascade for TPA-induced MMP-9 expression, the expression
of three MAPKs, and the DNA binding capacity of transcription
factors, were also investigated. These MAPKs (ERK, p38 and JNK) are
upstream modulators of NF-κB, and activate MMP-9 expression
(59). MAPKs are expressed in
MCF-7 breast cancer cells and their activation can be confirmed by
analyzing their phosphorylation (60). MAPKs are required for the
activation of NF-κB and AP-1, which requires IκB kinase, MAPKs and
PI3K/Akt, depending on the cell type in question (39,61,62).
Herein, matriptase-knockdown suppressed the phosphorylation of p38,
ERK, JNK and IKK following TPA treatment (Fig. 3B and C). NF-κB and AP-1 are
important for the expression of MMP-9 in MCF-7 cells, and the MMP9
gene promoter contains NF-κB and AP-1 binding sites (63). The present study revealed that
inhibition of matriptase expression inhibited TPA-induced MMP-9
expression by inhibiting NF-κB and AP-1 activation in MCF-7 breast
cancer cells (Fig. 4).
The primarily aim of the present study was to
identify the MMP-regulated signaling mechanism for the
matriptase-induced inhibition of cellular metastatic capacity.
Inhibition of matriptase expression was found to attenuate
TPA-induced MMP-9 expression and invasiveness by blocking NF-κB and
AP-1 activation through the PAR-2/PLCγ2 and PKC/MAPK signaling
pathways in MCF-7 breast cancer cells. Therefore, to the best of
our knowledge, the present study is the first to demonstrated that
MCF-7 breast cancer cell invasiveness is mediated by inhibiting
MMP-9 expression through modulation of the PAR-2/PLCγ2-mediated PKC
signaling pathway, induced by matriptase. These findings suggest
that inhibiting matriptase may have potential therapeutic value in
the treatment of breast cancer metastasis. Furthermore, these
findings are expected to pave the way for in vivo and
clinical studies to determine the efficacy of matriptase in
preventing breast cancer metastasis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Research Foundation of Korea funded by the Ministry of Education,
Science and Technology, Republic of Korea (grant nos.
2013R1A1A1059747 and 2013R1A1A2007181).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HJY, YRL and JP designed the study and confirmed the
authenticity of all the raw data. JMK, EMN and HKS performed the
experiments. SYK and JSK analyzed the data. SHJ contributed to data
analysis and interpretation, and critically revised the manuscript.
YRL drafted the manuscript. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Redig AJ and McAllister SS: Breast cancer
as a systemic disease: A view of metastasis. J Intern Med.
274:113–126. 2013. View Article : Google Scholar
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
3
|
Leber MF and Efferth T: Molecular
principles of cancer invasion and metastasis (review). Int J Oncol.
34:881–895. 2009.
|
|
4
|
Jiang WG, Sanders AJ, Katoh M, Ungefroren
H, Gieseler F, Prince M, Thompson SK, Zollo M, Spano D, Dhawan P,
et al: Tissue invasion and metastasis: Molecular, biological and
clinical perspectives. Semin Cancer Biol. 35 (Suppl 1):S244–S275.
2015. View Article : Google Scholar
|
|
5
|
van Zijl F, Krupitza G and Mikulits W:
Initial steps of metastasis: Cell invasion and endothelial
transmigration. Mutat Res. 728:23–34. 2011. View Article : Google Scholar
|
|
6
|
Theocharis AD, Skandalis SS, Gialeli C and
Karamanos NK: Extracellular matrix structure. Adv Drug Deliv Rev.
97:4–27. 2016. View Article : Google Scholar
|
|
7
|
Szabo R and Bugge TH: Type II
transmembrane serine proteases in development and disease. Int J
Biochem Cell Biol. 40:1297–1316. 2008. View Article : Google Scholar
|
|
8
|
Murray AS, Varela FA and List K: Type II
transmembrane serine proteases as potential targets for cancer
therapy. Biol Chem. 397:815–826. 2016. View Article : Google Scholar
|
|
9
|
List K, Bugge TH and Szabo R: Matriptase:
Potent proteolysis on the cell surface. Mol Med. 12:1–7. 2006.
View Article : Google Scholar
|
|
10
|
Oberst M, Anders J, Xie B, Singh B,
Ossandon M, Johnson M, Dickson RB and Lin CY: Matriptase and HAI-1
are expressed by normal and malignant epithelial cells in vitro and
in vivo. Am J Pathol. 158:1301–1311. 2001. View Article : Google Scholar
|
|
11
|
Bergum C, Zoratti G, Boerner J and List K:
Strong expression association between matriptase and its substrate
prostasin in breast cancer. J Cell Physiol. 227:1604–1609. 2012.
View Article : Google Scholar
|
|
12
|
Tuhkanen H, Hartikainen JM, Soini Y,
Velasco G, Sironen R, Nykopp TK, Kataja V, Eskelinen M, Kosma VM
and Mannermaa A: Matriptase-2 gene (TMPRSS6) variants associate
with breast cancer survival, and reduced expression is related to
triple-negative breast cancer. Int J Cancer. 133:2334–2340. 2013.
View Article : Google Scholar
|
|
13
|
Parr C, Sanders AJ, Davies G, Martin T,
Lane J, Mason MD, Mansel RE and Jiang WG: Matriptase-2 inhibits
breast tumor growth and invasion and correlates with favorable
prognosis for breast cancer patients. Clin Cancer Res.
13:3568–3576. 2007. View Article : Google Scholar
|
|
14
|
Rattenholl A and Steinhoff M:
Proteinase-activated receptor-2 in the skin: Receptor expression,
activation and function during health and disease. Drug News
Perspect. 21:369–381. 2008. View Article : Google Scholar
|
|
15
|
Bao Y, Hou W and Hua B: Protease-activated
receptor 2 signalling pathways: A role in pain processing. Expert
Opin Ther Targets. 18:15–27. 2014. View Article : Google Scholar
|
|
16
|
Sales KU, Friis S, Konkel JE, Godiksen S,
Hatakeyama M, Hansen KK, Rogatto SR, Szabo R, Vogel LK, Chen W, et
al: Non-hematopoietic PAR-2 is essential for matriptase-driven
pre-malignant progression and potentiation of ras-mediated squamous
cell carcinogenesis. Oncogene. 34:346–356. 2015. View Article : Google Scholar
|
|
17
|
Wojtukiewicz MZ, Hempel D, Sierko E,
Tucker SC and Honn KV: Protease-activated receptors (PARs)-biology
and role in cancer invasion and metastasis. Cancer Metastasis Rev.
34:775–796. 2015. View Article : Google Scholar
|
|
18
|
Bocheva G, Rattenholl A, Kempkes C, Goerge
T, Lin CY, D'Andrea MR, Ständer S and Steinhoff M: Role of
matriptase and proteinase-activated receptor-2 in nonmelanoma skin
cancer. J Invest Dermatol. 129:1816–1823. 2009.Rothmeier AS and Ruf
W: Protease-activated receptor 2 signaling in inflammation. Semin
Immunopathol 34: 133–149, 2012. View Article : Google Scholar
|
|
19
|
Rothmeier AS and Ruf W: Protease-activated
receptor 2 signaling in inflammation. Semin Immunopathol.
34:133–149. 2012. View Article : Google Scholar
|
|
20
|
Lidington EA, Steinberg R, Kinderlerer AR,
Landis RC, Ohba M, Samarel A, Haskard DO and Mason JC: A role for
proteinase-activated receptor 2 and PKC-epsilon in
thrombin-mediated induction of decay-accelerating factor on human
endothelial cells. Am J Physiol Cell Physiol. 289:C1437–C1447.
2005. View Article : Google Scholar
|
|
21
|
van der Merwe JQ, Moreau F and MacNaughton
WK: Protease-activated receptor-2 stimulates intestinal epithelial
chloride transport through activation of PLC and selective PKC
isoforms. Am J Physiol Gastrointest Liver Physiol. 296:G1258–G1266.
2009. View Article : Google Scholar
|
|
22
|
Su S, Li Y, Luo Y, Sheng Y, Su Y, Padia
RN, Pan ZK, Dong Z and Huang S: Proteinase-activated receptor 2
expression in breast cancer and its role in breast cancer cell
migration. Oncogene. 28:3047–3057. 2009. View Article : Google Scholar
|
|
23
|
Jiang Y, Yau MK, Lim J, Wu KC, Xu W, Suen
JY and Fairlie DP: A potent antagonist of protease-activated
receptor 2 that inhibits multiple signaling functions in human
cancer cells. J Pharmacol Exp Ther. 364:246–257. 2018. View Article : Google Scholar
|
|
24
|
Stetler-Stevenson WG, Hewitt R and
Corcoran M: Matrix metalloproteinases and tumor invasion: From
correlation and causality to the clinic. Semin Cancer Biol.
7:147–154. 1996. View Article : Google Scholar
|
|
25
|
Itoh Y and Nagase H: Matrix
metalloproteinases in cancer. Essays Biochem. 38:21–36. 2002.
View Article : Google Scholar
|
|
26
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: A tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar
|
|
27
|
Lin CW, Hou WC, Shen SC, Juan SH, Ko CH,
Wang LM and Chen YC: Quercetin inhibition of tumor invasion via
suppressing PKC delta/ERK/AP-1-dependent matrix metalloproteinase-9
activation in breast carcinoma cells. Carcinogenesis. 29:1807–1815.
2008. View Article : Google Scholar
|
|
28
|
Lee SO, Jeong YJ, Kim M, Kim CH and Lee
IS: Suppression of PMA-induced tumor cell invasion by capillarisin
via the inhibition of NF-kappaB-dependent MMP-9 expression. Biochem
Biophys Res Commun. 366:1019–1024. 2008. View Article : Google Scholar
|
|
29
|
Saito N, Hatori T, Murata N, Zhang ZA,
Ishikawa F, Nonaka H, Iwabuchi S and Samejima H: A double
three-step theory of brain metastasis in mice: The role of the pia
mater and matrix metalloproteinases. Neuropathol Appl Neurobiol.
33:288–298. 2007. View Article : Google Scholar
|
|
30
|
Castellano G, Malaponte G, Mazzarino MC,
Figini M, Marchese F, Gangemi P, Travali S, Stivala F, Canevari S
and Libra M: Activation of the osteopontin/matrix
metalloproteinase-9 pathway correlates with prostate cancer
progression. Clin Cancer Res. 14:7470–7480. 2008. View Article : Google Scholar
|
|
31
|
Kanayama H: Matrix metalloproteinases and
bladder cancer. J Med Invest. 48:31–43. 2001.
|
|
32
|
Gum R, Wang H, Lengyel E, Juarez J and
Boyd D: Regulation of 92 kDa type IV collagenase expression by the
jun aminoterminal kinase- and the extracellular signal-regulated
kinase-dependent signaling cascades. Oncogene. 14:1481–1493. 1997.
View Article : Google Scholar
|
|
33
|
Newton AC: Regulation of protein kinase C.
Curr Opin Cell Biol. 9:161–167. 1997. View Article : Google Scholar
|
|
34
|
Zeigler ME, Chi Y, Schmidt T and Varani J:
Role of ERK and JNK pathways in regulating cell motility and matrix
metalloproteinase 9 production in growth factor-stimulated human
epidermal keratinocytes. J Cell Physiol. 180:271–284. 1999.
View Article : Google Scholar
|
|
35
|
Hozumi A, Nishimura Y, Nishiuma T, Kotani
Y and Yokoyama M: Induction of MMP-9 in normal human bronchial
epithelial cells by TNF-alpha via NF-kappa B-mediated pathway. Am J
Physiol Lung Cell Mol Physiol. 281:L1444–L1452. 2001. View Article : Google Scholar
|
|
36
|
Weng CJ, Chau CF, Hsieh YS, Yang SF and
Yen GC: Lucidenic acid inhibits PMA-induced invasion of human
hepatoma cells through inactivating MAPK/ERK signal transduction
pathway and reducing binding activities of NF-kappaB and AP-1.
Carcinogenesis. 29:147–156. 2008. View Article : Google Scholar
|
|
37
|
Noh EM, Park YJ, Kim JM, Kim MS, Kim HR,
Song HK, Hong OY, So HS, Yang SH, Kim JS, et al: Fisetin regulates
TPA-induced breast cell invasion by suppressing matrix
metalloproteinase-9 activation via the PKC/ROS/MAPK pathways. Eur J
Pharmacol. 764:79–86. 2015. View Article : Google Scholar
|
|
38
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim BS, Lee SH, Lee SJ, et al: Curcumin
suppresses the TPA-induced invasion through inhibition of
PKCα-dependent MMP-expression in MCF-7 human breast cancer cells.
Phytomedicine. 19:1085–1092. 2012. View Article : Google Scholar
|
|
39
|
Karin M: The regulation of AP-1 activity
by mitogen-activated protein kinases. J Biol Chem. 270:16483–16486.
1995. View Article : Google Scholar
|
|
40
|
Madrid LV, Mayo MW, Reuther JY and Baldwin
AS Jr: Akt stimulates the transactivation potential of the RelA/p65
Subunit of NF-kappa B through utilization of the Ikappa B kinase
and activation of the mitogen-activated protein kinase p38. J Biol
Chem. 276:18934–18940. 2001. View Article : Google Scholar
|
|
41
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar
|
|
42
|
Isakov N: Protein kinase C (PKC) isoforms
in cancer, tumor promotion and tumor suppression. Semin Cancer
Biol. 48:36–52. 2018. View Article : Google Scholar
|
|
43
|
Duffy MJ, Maguire TM, Hill A, McDermott E
and O'Higgins N: Metalloproteinases: Role in breast carcinogenesis,
invasion and metastasis. Breast Cancer Res. 2:252–257. 2000.
View Article : Google Scholar
|
|
44
|
Woessner JF Jr: Matrix metalloproteinases
and their inhibitors in connective tissue remodeling. FASEB J.
5:2145–2154. 1991. View Article : Google Scholar
|
|
45
|
Scorilas A, Karameris A, Arnogiannaki N,
Ardavanis A, Bassilopoulos P, Trangas T and Talieri M:
Overexpression of matrix-metalloproteinase-9 in human breast
cancer: A potential favourable indicator in node-negative patients.
Br J Cancer. 84:1488–1496. 2001. View Article : Google Scholar
|
|
46
|
Yousef EM, Tahir MR, St-Pierre Y and
Gaboury LA: MMP-9 expression varies according to molecular subtypes
of breast cancer. BMC Cancer. 14:6092014. View Article : Google Scholar
|
|
47
|
Shi YE, Torri J, Yieh L, Wellstein A,
Lippman ME and Dickson RB: Identification and characterization of a
novel matrix-degrading protease from hormone-dependent human breast
cancer cells. Cancer Res. 53:1409–1415. 1993.
|
|
48
|
Fan B, Brennan J, Grant D, Peale F,
Rangell L and Kirchhofer D: Hepatocyte growth factor activator
inhibitor-1 (HAI-1) is essential for the integrity of basement
membranes in the developing placental labyrinth. Dev Biol.
303:222–230. 2007. View Article : Google Scholar
|
|
49
|
Uhland K: Matriptase and its putative role
in cancer. Cell Mol Life Sci. 63:2968–2978. 2006. View Article : Google Scholar
|
|
50
|
Zoratti GL, Tanabe LM, Varela FA, Murray
AS, Bergum C, Colombo É, Lang JE, Molinolo AA, Leduc R, Marsault E,
et al: Targeting matriptase in breast cancer abrogates tumour
progression via impairment of stromal-epithelial growth factor
signalling. Nat Commun. 6:67762015. View Article : Google Scholar
|
|
51
|
Zhang J, Anastasiadis PZ, Liu Y, Thompson
EA and Fields AP: Protein kinase C (PKC) betaII induces cell
invasion through a Ras/Mek-, PKC iota/Rac 1-dependent signaling
pathway. J Biol Chem. 279:22118–22123. 2004. View Article : Google Scholar
|
|
52
|
de Vente JE, Kukoly CA, Bryant WO,
Posekany KJ, Chen J, Fletcher DJ, Parker PJ, Pettit GJ, Lozano G
and Cook PP: Phorbol esters induce death in MCF-7 breast cancer
cells with altered expression of protein kinase C isoforms. Role
for p53-independent induction of gadd-45 in initiating death. J
Clin Invest. 96:1874–1886. 1995. View Article : Google Scholar
|
|
53
|
Kim S, Han J, Lee SK, Choi MY, Kim J, Lee
J, Jung SP, Kim JS, Kim JH, Choe JH, et al: Berberine suppresses
the TPA-induced MMP-1 and MMP-9 expressions through the inhibition
of PKC-α in breast cancer cells. J Surg Res. 176:e21–e29. 2012.
View Article : Google Scholar
|
|
54
|
Barry OP and Kazanietz MG: Protein kinase
C isozymes, novel phorbol ester receptors and cancer chemotherapy.
Curr Pharm Des. 7:1725–1744. 2001. View Article : Google Scholar
|
|
55
|
Newton AC: Protein kinase C as a tumor
suppressor. Semin Cancer Biol. 48:18–26. 2018. View Article : Google Scholar
|
|
56
|
Wilde JI and Watson SP: Regulation of
phospholipase C gamma isoforms in haematopoietic cells: Why one,
not the other? Cell Signal. 13:691–701. 2001. View Article : Google Scholar
|
|
57
|
Koivunen J, Aaltonen V and Peltonen J:
Protein kinase C (PKC) family in cancer progression. Cancer Lett.
235:1–10. 2006. View Article : Google Scholar
|
|
58
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim BS, Lee SH, Lee SJ, et al: Curcumin
suppresses the TPA-induced invasion through inhibition of
PKCα-dependent MMP-expression in MCF-7 human breast cancer cells.
Phytomedicine. 19:1085–1092. 2012. View Article : Google Scholar
|
|
59
|
Chakraborti S, Mandal M, Das S, Mandal A
and Chakraborti T: Regulation of matrix metalloproteinases: An
overview. Mol Cell Biochem. 253:269–285. 2003. View Article : Google Scholar
|
|
60
|
Qi M and Elion EA: MAP kinase pathways. J
Cell Sci. 118:3569–3572. 2005. View Article : Google Scholar
|
|
61
|
Yao J, Xiong S, Klos K, Nguyen N, Grijalva
R, Li P and Yu D: Multiple signaling pathways involved in
activation of matrix metalloproteinase-9 (MMP-9) by heregulin-beta1
in human breast cancer cells. Oncogene. 20:8066–8074. 2001.
View Article : Google Scholar
|
|
62
|
Whitmarsh AJ: Regulation of gene
transcription by mitogen-activated protein kinase signaling
pathways. Biochim Biophys Acta. 1773:1285–1298. 2007. View Article : Google Scholar
|
|
63
|
Eberhardt W, Huwiler A, Beck KF, Walpen S
and Pfeilschifter J: Amplification of IL-1 beta-induced matrix
metalloproteinase-9 expression by superoxide in rat glomerular
mesangial cells is mediated by increased activities of NF-kappa B
and activating protein-1 and involves activation of the
mitogen-activated protein kinase pathways. J Immunol.
165:5788–5797. 2000. View Article : Google Scholar
|