Esophageal cancer (EC) is the seventh most common
type of cancer in the world and the sixth leading cause of
cancer-related deaths (1). At
present, the morbidity and mortality rates of EC are exhibiting an
increasing trend annually (2), and
the overall 5-year survival rate is only 20% (3). There are two primary pathological
types of EC: Esophageal adenocarcinoma (EAC) and esophageal
squamous cell carcinoma (ESCC). ESCC is the most common
pathological type, accounting for ~84% of EC cases, whereas the
rates of EAC are increasing in high-income countries (4). Significant geographical differences
have been found in the incidence of EC; the incidence of EC is
highest in East Asia, followed by South Africa and East Africa
(1). It is worth mentioning that
gastroesophageal reflux disease (GERD) can cause Barrett esophagus
(BE), which is the precursor lesion to EAC (5). It is estimated that 10-15% of GERD
patients will develop BE, and the existence and grading of
dysplasia in BE are the most important predictors of EAC (6). There are also studies highlighting
the involvement of sex (4),
obesity (7), smoking (8), and alcohol consumption (9) in the development of EC. Due to the
massive amount of lymphatic drainage into the submucosa of the
esophagus, lymphatic metastasis occurs in the early stage of EC,
which leads to a lower survival rate (10). Recent findings indicated that
patients with EC exhibited metastasis to adjacent and distant
tissues, such as the liver (11),
lungs (12), and bones (13). Thus, the occurrence and metastasis
of EC appear to be related to several factors. To improve the
management of this life-threatening disease, the factors involved
in the development of EC as well as those involved in its malignant
transformation need to be determined.
With the development of molecular biology research,
a considerable body of evidence has indicated that several
molecular pathways are involved in the acquisition of malignancy in
EC, which may pave the way for effective EC therapies. The hedgehog
family is highly conserved in mammals and consists of three
proteins: Sonic hedgehog (Shh), Indian hedgehog, and Desert
hedgehog (14). Hedgehog signaling
is generally involved in the development and differentiation of
embryonic tissues, hemostasis of normal adult cells, and
carcinogenesis (15). Shh acts on
the activity of the GLI family, including GLI1, GLI2, and GLI3, for
regulating gene expression (16).
In EC, GLI1 was overexpressed and linked to epithelial-mesenchymal
transformation (EMT) and lymph node metastasis (17). Research has revealed that GLI1 can
activate cyclin D2 and FOXM1, serving an oncogenic role (18). The Wnt signaling pathway is also
confirmed to exert a carcinogenic effect in several types of
tissues when aberrantly activated (19). Wnt binds to the cysteine-rich
domain of the Frizzled (Fzd) family receptors, preventing β-catenin
phosphorylation and ubiquitination by activating Dishevelled.
β-Catenin then enters the nucleus, where it binds to the
transcriptional complex consisting of the T-cell factor (TCF)-1,
BCL-9, and Pygopus 2 to promote cancer cell development and
metastasis (20). Upregulation of
the Wnt signaling pathway has been demonstrated to enhance the
proliferation and motility of EC cells (21-23).
The NOTCH pathway is mediated through ligands binding to NOTCH
receptors and regulating squamous differentiation in the skin and
esophagus (24). In ESCC, the
NOTCH pathway exerts a carcinogenic effect when aberrantly
activated, which promotes EMT and tumorigenesis by mediating the
expression of transforming growth factor-β and thus increasing the
number of tumor stem cells (25).
Additionally, special attention should be paid to
one member of the signal transducer and activator of transcription
(STAT) family: STAT3. The STAT family was first discovered in 1994
when evaluating the molecular pathways involved in gene regulation
triggered by interferon (IFN) molecules, which are mediators that
transmit signals from the extracellular matrix to the nucleus
(26,27). A total of seven STAT proteins have
been identified to date: STAT1, 2, 3, 4, 5a, 5b, and 6, and they
have been shown to mediate multiple cellular functions (28). STAT1 and STAT2 primarily take part
in the inhibition of the development of cancer, whereas STAT3,
STAT4, and STAT5 are commonly regarded as cancer promoters
(29). Among these STAT subtypes,
STAT3 has been confirmed to be activated in various types of
cancer, promoting tumor cell proliferation, angiogenesis, and
metastasis, thus attracting the attention of researchers (30,31).
In recent years, an increasing number of researchers have begun to
investigate the relationship between STAT3 and EC. Therefore, a
comprehensive search of literature published until the end of March
2022 was conducted using PubMed. Various combinations of keywords
were used for searching: 'Signal transducer and activator of
transcription 3' or 'STAT3', 'phosphorylated signal transducer and
activator of transcription 3' or 'p-STAT3', 'esophageal' or
'esophagus', and 'cancer' or 'carcinoma'. In addition, studies
related to the epidemiology of EC, the molecular structure of
STAT3, the role of STAT3 and its related molecules in various
cancers, and other signaling pathways in EC were also searched for
to fully present the topic. Duplicate data, research not relevant
to the topic of the present review, and non-English studies were
excluded. As a result, a total of 211 studies were cited in the
present review to summarize the research progress regarding STAT3
in EC.
The STAT3 coding gene is located on chromosome 12
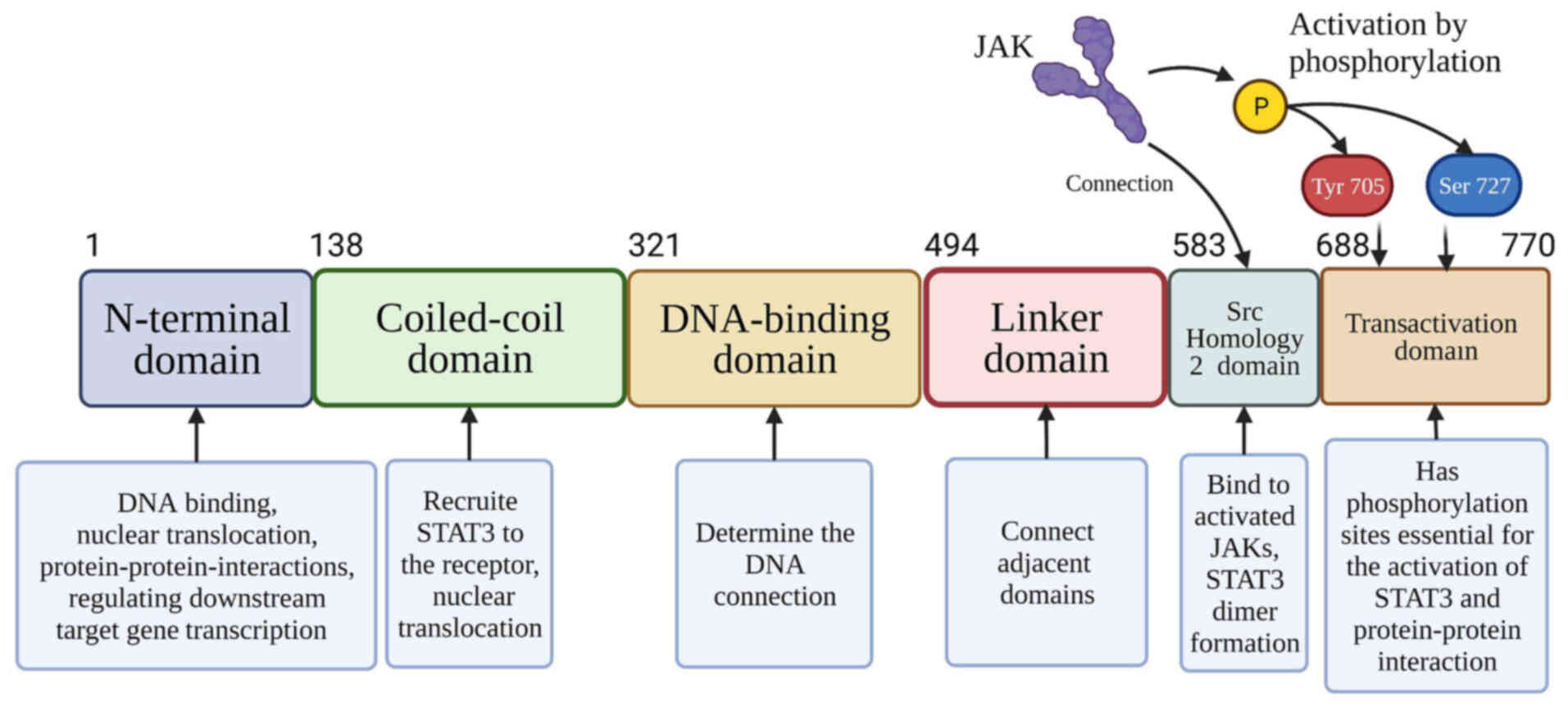
(q13-q14-1) in humans. The STAT3 protein has a unique secondary
structure consisting of six primary domains (Fig. 1): i) An N-terminal domain (NTD),
which has the functions of binding to DNA, nuclear translocation,
interactions between proteins, and regulating transcription of
downstream target genes (28); ii)
a coiled-coil domain that is critical in recruitment of STAT3 to
its receptor and in nuclear translocation (26,27).
iii) a DNA-binding domain that plays an essential role in
determining the DNA connection (32); iv) a linker domain (LD) that
connects the DNA-binding domain with the SH2 domain; v) an Src
Homology 2 (SH2) domain that can bind to and activate the JAK of
the cytokine-receptor cytoplasmic domain to induce STAT3 protein
recruitment to JAK, as STAT3 dimer formation relies on the
interactions of the SH2 domain (31); and vi) a transactivation domain
(TAD) which is a highly disordered and conserved domain that
possesses phosphorylation sites essential for the activation of
STAT3 and protein-protein interactions (33). Researchers have identified four
STAT3 isoforms: STAT3α, STAT3β, STAT3γ, and STAT3δ (34). STAT3α is a 92-kDa in length isoform
expressed in most cells, which has two crucial phosphorylation
sites (Yyr705 and Ser727) at the C-terminus (35). STAT3β has different splice isoforms
and only one phosphorylation site (Tyr705), whose TAD has only 7
residues, whereas the TAD in STAT3α has 50 residues (33). STAT3γ and STAT3δ are truncated
forms of STAT3α and play a role in granulocytic differentiation
(35).
A variety of signals can activate STAT3, including
cytokines such as interleukin (IL)-6 (36-38)
and leptin (39-41); growth factors including epidermal
growth factor (EGF) (42,43) and platelet-derived growth factor
(PDGF) (44); other molecules such
as cellular Src (c-Src) tyrosine kinase (45-47),
and thyroid-stimulating hormone (TSH) (48); and environmental factors including
nicotine (49), infections
(50), and stress (51), amongst others. These signals
activate STAT3 through phosphorylation of Ser727 or Tyr705 in the
TAD via several signaling pathways, form dimers via the SH2 domain,
then translocate into the nucleus to regulate gene transcription.
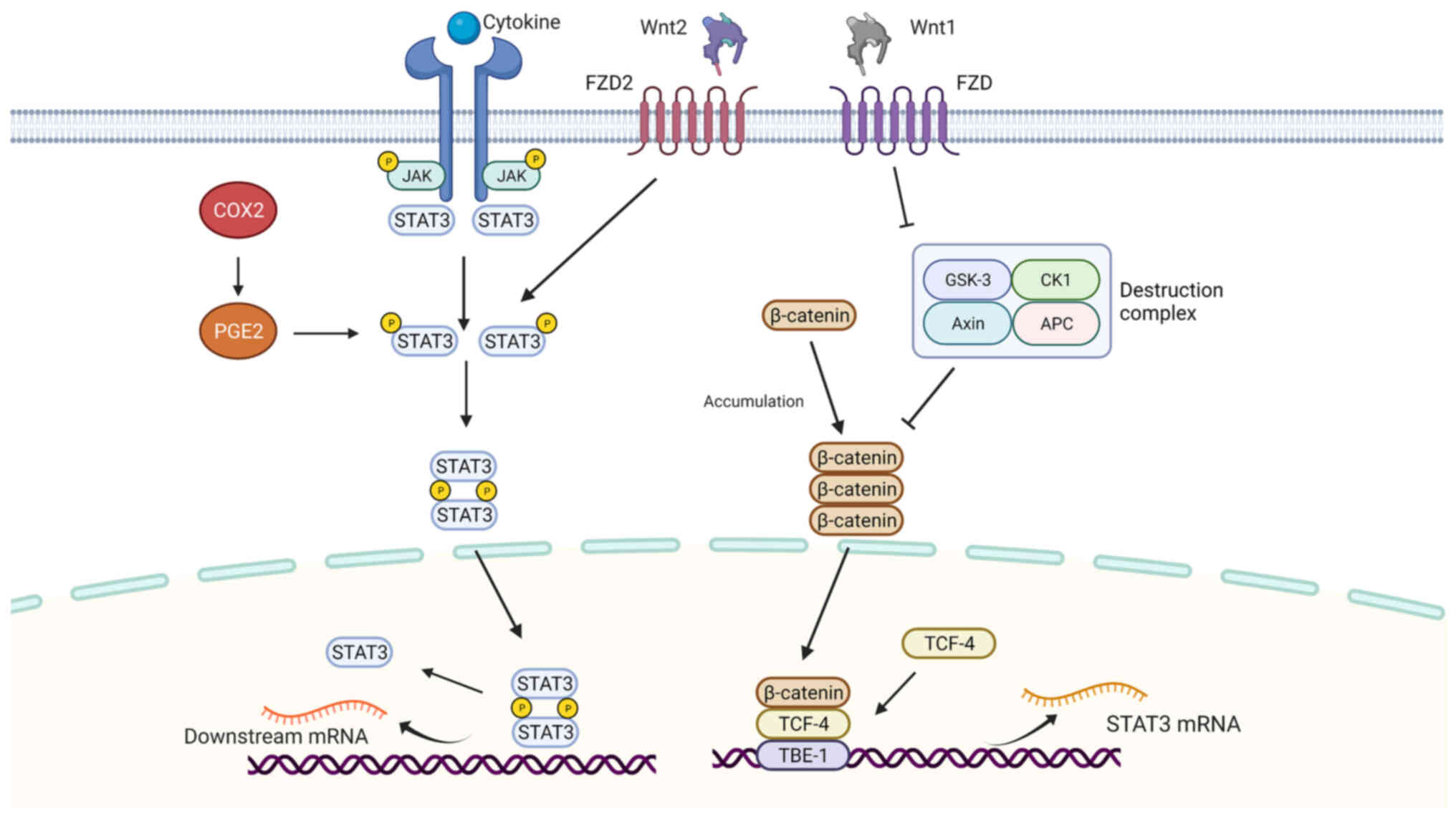
The JAK/STAT3 pathway is a canonical pathway of signal transduction
and transcriptional activation of STAT3 (Fig. 2). When activators bind to the
corresponding receptors on the cell membrane, the conformational
change in the receptors results in their dimerization or
oligomerization and thus activation (35,36).
Following activation, the cytoplasmic parts of the receptor bind to
each other, and the receptor-coupled tyrosine kinases are activated
by cross-phosphorylation. These tyrosine kinases are often Janus
kinase (JAK) family members, including JAK1, JAK2, JAK3, and TYK2.
Activated JAK kinase can phosphorylate a specific receptor in the
cytoplasm and activate the receptor (35,52).
The molecular configuration of the receptor changes to provide a
binding site for activating STAT3 monomers in the cytoplasm. STAT3
molecules recognize and bind to the receptor through the SH2
domain. JAK or Src, using the STAT3 molecule recruited by the
receptor as the substrate, phosphorylates the tyrosine (Tyr-705)
residue of the STAT3 molecule. Two phosphorylated STAT3 (p-STAT3)
monomers combine with the phosphorylated tyrosine residue of SH2 to
form a dimer and separate from the receptor, which is then
transferred into the nucleus where they bind to specific DNA
sequences, regulating downstream gene transcription (53). This transport process primarily
relies on the coiled-coil and DNA binding domains. Nuclear protein
tyrosine phosphatases such as TC45 are required for the
dephosphorylation of STAT3 in the nucleus. The inhibition of these
enzymes can lead to the downregulation of dephosphorylation of
STAT3, which results in continuous activation of STAT in the tumor
cell nucleus (54).
In the canonical STAT3 signaling pathway, STAT3 is
activated by JAK, but there are other STAT3 regulators that affect
the regulation of STAT3 (Fig.
2).
The cyclooxygenase (COX2)/prostaglandin E2
(PGE2)/STAT3 pathway is typical of non-canonical STAT3 signaling
pathways. COX is a rate-limiting enzyme that can catalyze
arachidonic acid conversion to prostaglandins to induce the
production of PGE2. COX2 expression is upregulated in ESCC. STAT3
has been indicated as the downstream target of COX2/PGE2 that is
involved in the induction of proliferation and EMT in EC tumors
(55,56).
The Wnt signal transduction pathway controls
numerous biological processes. The Wnt pathway can be divided into
canonical (β-catenin dependent) and non-canonical (β-catenin
independent) signaling; and both can modulate the STAT3 signaling
pathway.
In canonical Wnt/STAT3 signaling, Wnt-1 binds to its
cognate receptors, inhibiting the activity of destruction complexes
formed by Axin, APC, GSK3, and CK1, thus releasing and in turn
increasing the accumulation of β-catenin in the plasma (57). In ESCC, Wnt-1 induces β-catenin
accumulation and activates the TCF reporter gene (58). β-Catenin is transferred to the
nucleus, where it forms complexes with TCF4, and it has DNA binding
activity, regulating the transcription of genes. Further research
indicated that STAT3 promoters have five putative TCF4 binding
elements (TBEs). Among them, TBE1 specifically binds to TCF4
protein. Therefore, β-catenin/TCF4 increased STAT3 mRNA and protein
expression levels by binding to TBE1 (59).
In non-canonical Wnt/STAT3 signaling, one of the
Wnt2 transmembrane receptors from the frizzled (FZD) family, FZD2,
plays a vital role. FZD2 is upregulated in several types of cancer,
which is suggested to be a predictor of tumor recurrence (60). FZD2 was overexpressed in ESCC and
promoted the migration and invasion of ESCC cells and was
associated with a poor prognosis. FZD2 was revealed to directly
activate STAT3 at the Tyr705 site in ESCC, activating downstream
genes including TWIST1 and Slug (61).
By 2012, researchers determined that the activation
of STAT3 induces abnormal proliferation and migration of ESCC
(62). In subsequent studies, the
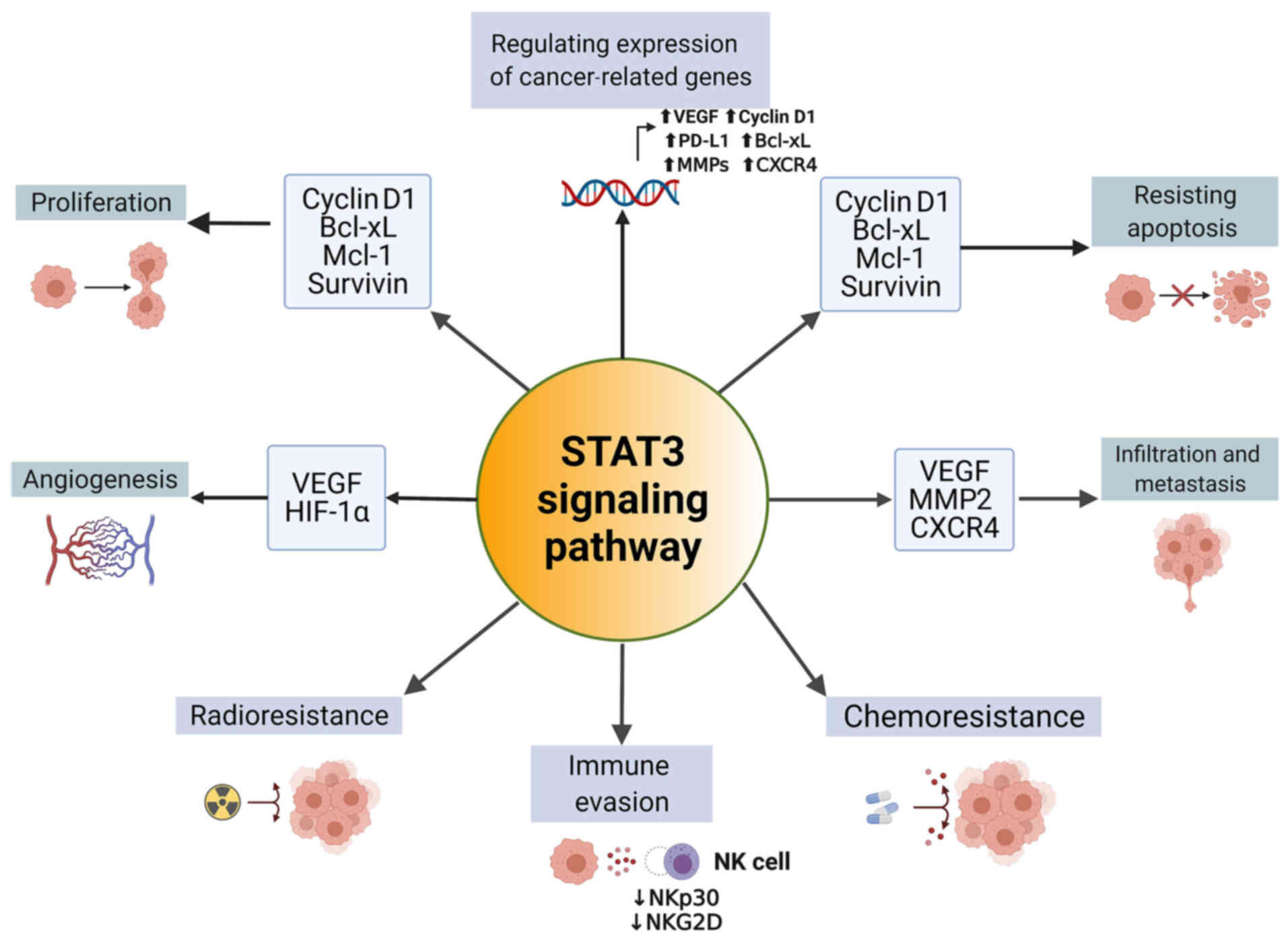
roles of STAT3 in EC have been gradually revealed (Fig. 3).
Activating STAT3 signaling promotes proliferation
and inhibits apoptosis in ESCC (63). The evidence from loss of function
studies has also confirmed that STAT3 acts as an oncogene in EC
cells. Inactivation of STAT3 in EC cells has been revealed to lead
to cell apoptosis, cell cycle arrest, and inhibition of
proliferation (64-67), confirming the crucial role in the
proliferation of EC from another perspective. The downstream genes
of STAT3 play an essential role in this process.
The Bcl-2 protein family is closely related to
apoptosis and consists of several antiapoptotic proteins including
Bcl-xL, Bcl-2, Bcl-w, and Mcl. These proteins are critical factors
in regulating apoptosis and the survival of cells. In EC, STAT3
acts as an upstream factor of Bcl-xL and enhances its expression,
inhibiting the programmed death of tumor cells (68). Additionally, suppressing STAT3
activity leads to the downregulation of Bcl-2 and thus the
induction of apoptosis (64,69).
Caspase-3 is a critical factor in apoptosis, and it
regulates tumor cell repopulation (70). Caspase-3 can be activated through
cleavage of upstream caspases, including caspase-8 and caspase-9,
which interact with promoters of various pro-angiogenic genes, such
as the vascular endothelial growth factor (VEGF)A gene (71). In mice trials, p-STAT3 increased
the levels of caspase-3, causing loss of muscle mass (72). In EC, after inhibition of the
activation of STAT3, cleaved caspase-3 expression was upregulated,
suggesting the correlation between p-STAT3 and cleaved caspase-3
(73).
Cyclins are cell-cycle-associated nuclear proteins.
Cyclin D1, a member of the cyclin family, typically binds with its
catalytic chaperone cyclin-dependent protein kinase (CDK)4 or CDK6
to form a complex, that controls the progression of the G1 phase of
the cell cycle (74).
Tumorigenesis is closely associated to the cell cycle. As an
essential cell cycle regulator, cyclin D1 has been demonstrated to
be a significant prognostic predictor of ESCC (75). Zhang et al (76) indicated that p-STAT3 upregulated
cyclin D1 expression in ESCC and reduced the survival of patients.
Additionally, Li et al (77) revealed that high levels of STAT3
and cyclin D1 were associated with a poorer prognosis after
curative resection of ESCC, showing the potential application of
STAT3 and cyclin D1 as molecular predictors of the curative effect
of treatment.
Angiogenesis provides the essential nutrition for
cancer cell growth and metastasis. Yang et al (78) demonstrated that metformin
suppressed the JAK/STAT3 signaling pathway and tumor angiogenesis
in vivo, highlighting the roles of STAT3 in angiogenesis and
in the formation of the cancer microenvironment in EC. In other
studies, AR-42, a pan-histone deacetylase (HDAC) inhibitor,
decreased the levels of p-STAT3 in ESCC cells, resulting in an
anti-angiogenic outcome both in vitro and in vivo
(79). Moreover, as a
gasotransmitter in ESCC cells, exogenous H2S promoted
angiogenesis via upregulating the activity of the STAT3 signaling
pathway (80).
Infiltration and distant metastasis are
manifestations of malignant esophageal tumors in which STAT3 is
also involved. Research has shown that in EC, STAT3 was associated
with infiltration degree (pT) and pTNM stage, and p-STAT3 was
associated with pT, lymphatic metastasis (pN), and pTNM stage
(76). The function of STAT3 in
promoting cancer invasion and metastasis can be regulated by
upstream proteins, and realized through the expression of
downstream genes. T-LAK cell-originated protein kinase (TOPK) is a
type of MAPKK-like kinase that can activate the Src/GSK3β/STAT3
signaling pathway to promote the invasion and migration of ESCC
cells (46).
VEGF has been revealed to stimulate endothelial cell
proliferation and migration, in addition to promoting the malignant
progression of tumors. p-STAT3 was demonstrated to increase VEGF
expression in EC and induce infiltration and metastasis of tumors
(76).
MMPs are proteolytic enzymes that degrade the
extracellular matrix (ECM), promoting migration and invasion of
tumors. MMP2 is overexpressed in ESCC samples and this is
significantly associated with tumor invasion depth, clinical stage,
and lymph node metastasis (81).
p-STAT3 can bind to the MMP2 promoter at 648-641 bp (TTCTCGAA) to
induce MMP2 expression in a dose-dependent manner (82). In addition, tumor necrosis factor
receptor-associated protein 1 (TRAP1), a member of the
mitochondrial heat shock protein 90 family acts as an upstream
protein of the STAT3/MMP2 pathway, where it is involved in
mediating migration and invasion via STAT3/MMP2 (83).
CXC chemokine receptor 4 (CXCR4) belongs to the
G-protein coupled receptor (GPCR), which is the co-receptor of
HIV-1 (84). In EC, CXCR4
expression was revealed to be significantly upregulated, and the
levels of CXCR4 were downregulated after using STAT3 inhibitors
(85), indicating that CXCR4 is a
downstream molecule regulated by STAT3, providing another piece of
evidence that STAT3 promotes infiltration and metastasis.
Although the primary treatment for EC is surgery;
immunity, radiotherapy, and chemotherapy also serve an essential
role in the treatment regimen and in improving the prognosis of
patients with EC.
Natural killer (NK) cells are crucially involved in
innate immunity. ESCC cells secrete IL-6 and IL-8 to activate the
STAT3 signaling pathway of NK cells and decrease the activation of
receptors (NKp30 and NKG2D) on NK cells (86), which may be the mechanism by which
EC cells escape from innate immune cell surveillance. Additionally,
STAT3 can induce chemoresistance in EC cells. Activating
transcription factor 4 transactivates STAT3 to mediate multidrug
resistance in ESCC (87),
highlighting the critical role of STAT3 in drug resistance. In
addition, suppressing the activation of STAT3 was demonstrated to
enhance the sensitivity to cisplatin (88). For commonly used chemotherapeutic
drugs such as 5-fluorouracil (5-FU), cisplatin, and paclitaxel, the
IC50 values in EC cells were reduced when these drugs
were combined with STAT3 inhibitors such as niclosamide (89). In addition to resistance to
chemotherapy, STAT3 is involved in radioresistance as well. Zang
et al (90) demonstrated
that STAT3 activation is a critical event involved in ionizing
radiation-induced EMT and radioresistance, and inhibiting the
activation of STAT3 prevents these processes. Additionally, after
being treated with the STAT3 inhibitor Stattic, the levels of
p-STAT3, HIF-1α, and VEGF in EC cells decreased, resulting in an
increase in the radiosensitivity of EC cells both in vitro
and in vivo (91),
indicating the potential for the combined use of STAT3 inhibitors
with radiotherapy in improving the curative effects in patients
with EC.
Therefore, it is not surprising that the high levels
of STAT3 and p-STAT3 are closely associated with reduced survival
rates in patients with EC. The 5-year survival rate of patients
with ESCC was revealed to be significantly associated with p-STAT3
expression, and p-STAT3 was a relevant independent factor of a poor
prognosis and an independent prognostic factor for progression-free
survival (PFS) in ESCC (76).
miRNAs, lncRNAs, circRNAs, small nucleolar RNAs
(snoRNAs), and transfer RNAs (tRNAs) are non-coding RNAs (ncRNAs)
that are transcribed from DNA but are not translated into proteins
(92). An increasing body of
literature supports the theory that ncRNAs play an important role
in cancer development (93,94).
STAT3 is an upstream mediator of multiple transcription factors,
but it can also be modulated by ncRNAs themselves (95). miRNAs are small molecules 18-25
nucleotides in length and are generally transcribed by type II RNA
polymerases (96,97), and they play a crucial role in
modulating the translation of downstream genes by binding the
3'untranslated regions (UTRs) of mRNAs (98). Certain miRNAs have been revealed to
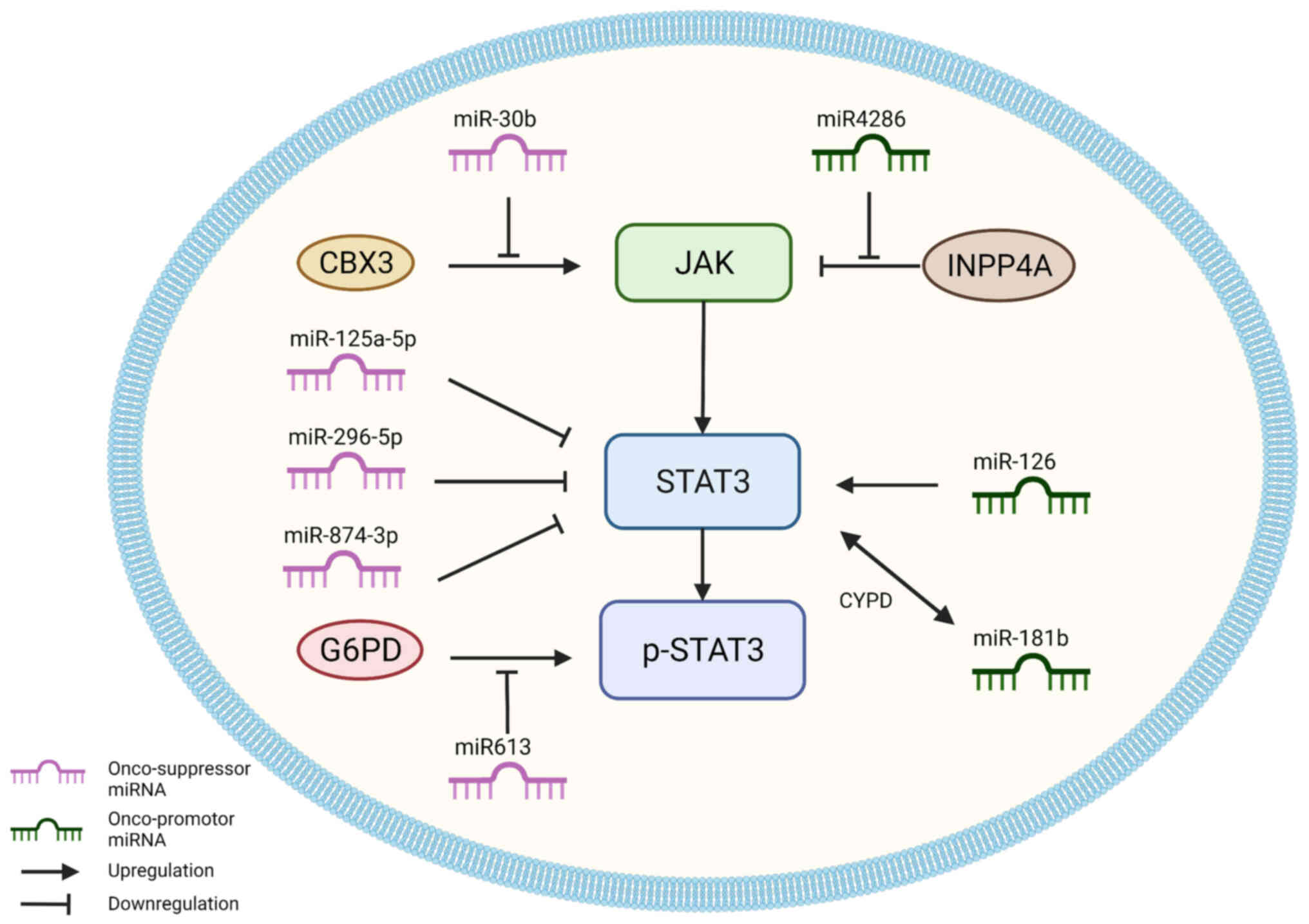
downregulate or upregulate STAT3 expression. Thus, these miRNAs can
be categorized into two types: Onco-suppressor miRNAs that suppress
STAT3 expression, and onco-promotor miRNAs that enhance the
expression of STAT3 (Fig. 4;
Table I).
Most miRNAs associated with STAT3 signaling play
tumor-suppressive roles, and some of them directly target STAT3 in
EC, including miR-125a-3p, miR-296-5p and miR-874-3p.
miR-125 is a highly conserved miRNA family that
consists of two sub-types, miR-125a and miR-125b. miR-125a is
further subdivided into miR-125a-3p, and miR-125a-5p, which are
derived from the 3′ (passenger strand) and 5′ (leading strand) ends
of pre-miR-125a, respectively. The miR-125 family is closely
associated with the STAT3 signaling pathway. For example, miR-125b
was demonstrated to regulate the levels of STAT3 in T cells
(99). In osteosarcoma, miR-125b
was revealed to be downregulated and negatively associated with
STAT3 expression (100). In ESCC
tissues and cells, the expression of miR-12a-5p was identified to
be low. Downregulation of miR125a-5p increased cell proliferation,
accelerated cell cycle progression, inhibited apoptosis, and
improved the migratory and invasive capacities of ESCC cells, which
were linked to EMT in ESCC, resulting in the progression of cancer
to more advanced tumor stages and a shorter survival duration as a
result. In ESCC cells, STAT3 was discovered to be a direct target
of miR-125a-5p, and overexpression of miR-125a-5p significantly
decreased STAT3, p-STAT3, and VEGF protein levels (88).
The miR-296 family consists of miR-296-5p and
miR-296-3p and is involved in tumorigenicity (101). In hepatocellular carcinoma,
miR-296 suppressed cell proliferation and increased apoptosis by
modulating fibroblast growth factor receptor 1 (FGFR1) (102). Additionally, miR-296 inhibited
colorectal cancer metastasis and EMT by targeting S100A (103). Wang et al (104) demonstrated that the levels of
miR-296-5p in ESCC are downregulated, and miR-296-5p directly
targets STAT3 to suppress STAT3 expression, thus inhibiting
migration and invasion of ESCC cells.
There are also antitumor miRNAs that indirectly
regulate the activity of JAK, including miR-30b and miR-613.
miR-30b plays essential roles in suppressing hepatocellular
carcinoma, colorectal cancer, gastric cancer, bladder cancer, and
breast cancer (117). In ESCC,
miR-30b acts as a JAK suppressor by downregulating chromobox 3
(CBX3) and inhibiting the migration and invasion of tumor cells
(118). CBX3 is a CBX protein,
which serves functions in stem cell self-renewal and cancer
development (119). Therefore,
miR-30b is associated with a better prognosis in ESCC (120).
Unlike other tumor suppressor miRNAs, miR-613
regulates STAT3 signaling pathways indirectly. miR-613 has been
shown to exert a tumor-suppressive role in colorectal cancer
(121), gastric cancer (122), lung cancer (123), hepatocellular carcinoma (124,125), and laryngeal carcinoma (126), by downregulating the expression
of oncogenes and reducing the malignant potential of tumors. In
ESCC, miR-613 targets glucose-6-phosphate dehydrogenase (G6PD).
G6PD can promote the phosphorylation of STAT3 and STAT5, which
miR-613 negatively regulates (127). Su et al (128) revealed that miR-613 expression
was reduced in ESCC tissues, whereas G6PD expression was increased
in ESCC tissues; miR-613 inhibited ESCC migration and invasion by
targeting G6PD and reducing MMP2, MMP9, and p-STAT3 expression.
Compared to tumor suppressor miRNAs in EC, there are
fewer tumor promoter miRNAs associated with STAT3 (Fig. 4). miR-4286 has been identified to
act as a tumor promoter of prostate cancer (129), gastric cancer (130), NSCLC (131), and glioblastomas (132). In EC, miR-4286 was demonstrated
to indirectly inhibit JAK, which is closely related to inositol
polyphosphate 4-phosphatase type I (INPP4A). INPP4A is a negative
regulator of Akt and is confirmed to function as a tumor suppressor
in breast cancer and prostate cancer (133). Zhang et al (134) found that miR-4286 was
overexpressed in and significantly promoted the viabilities of
TE-1, HCE-4, and HCE-7 cells, and it also activated the JAK2/STAT3
pathway by inhibiting the function of INPP4A.
miR-126 tumor regulation is dependent on the
specific tissue type and is essential to inflammation,
angiogenesis, and cell migration (135). In lung cancer, overexpression of
miR-126 resulted in decreased caspase-3 mRNA expression and
increased STAT3 protein expression, increasing tumor cell
proliferation and migration (136). However, in osteosarcoma and
cervical cancer, miR-126 played an opposite role. miR-126 inhibited
proliferation, migration, and invasion of cervical cancer cells
in vitro by reducing MMP2 and MMP9 production and
inactivating the JAK2/STAT3 signaling pathway by targeting ZEB1
(137). Additionally, miR-126
suppressed osteosarcoma proliferation, migration, invasion, and EMT
by targeting ZEB1 and inactivating the JNK and JAK1/STAT3 pathways
(138). The role of miR-126 in
ESCC was investigated in a study by Li et al (139), which revealed that miR-126 was
upregulated in TE13 cells, Eca109 cells, and ESCC tissues, and
targeted the 3'-UTR of STAT3, enhancing its transcription and
translation, and thus increasing tumor cell viability. Considering
its differing characteristics in other types of cancer, the
specific role of miR-126 under certain conditions of EC remains to
be determined.
Similarly, miR-181b has been shown to possess an
interesting association with STAT3. miR-181b is a member of the
miR-181 family, which affects a variety of tumor-related biological
processes including cell proliferation, apoptosis, autophagy,
mitochondrial function, and the immune response (140). In a previous study, STAT3 was
hypothesized to be the target of miR-181b in cutaneous melanoma
cells (141). In EC stem-like
cells, miR-181b increased the levels of STAT3, and overexpression
of STAT3 transactivated the levels of miR-181b. The reciprocal
activation of miR-181b and STAT3 was mediated by cylindromatosis
(CYLD) and enhanced EC stem-like cell viability and sphere
formation (142).
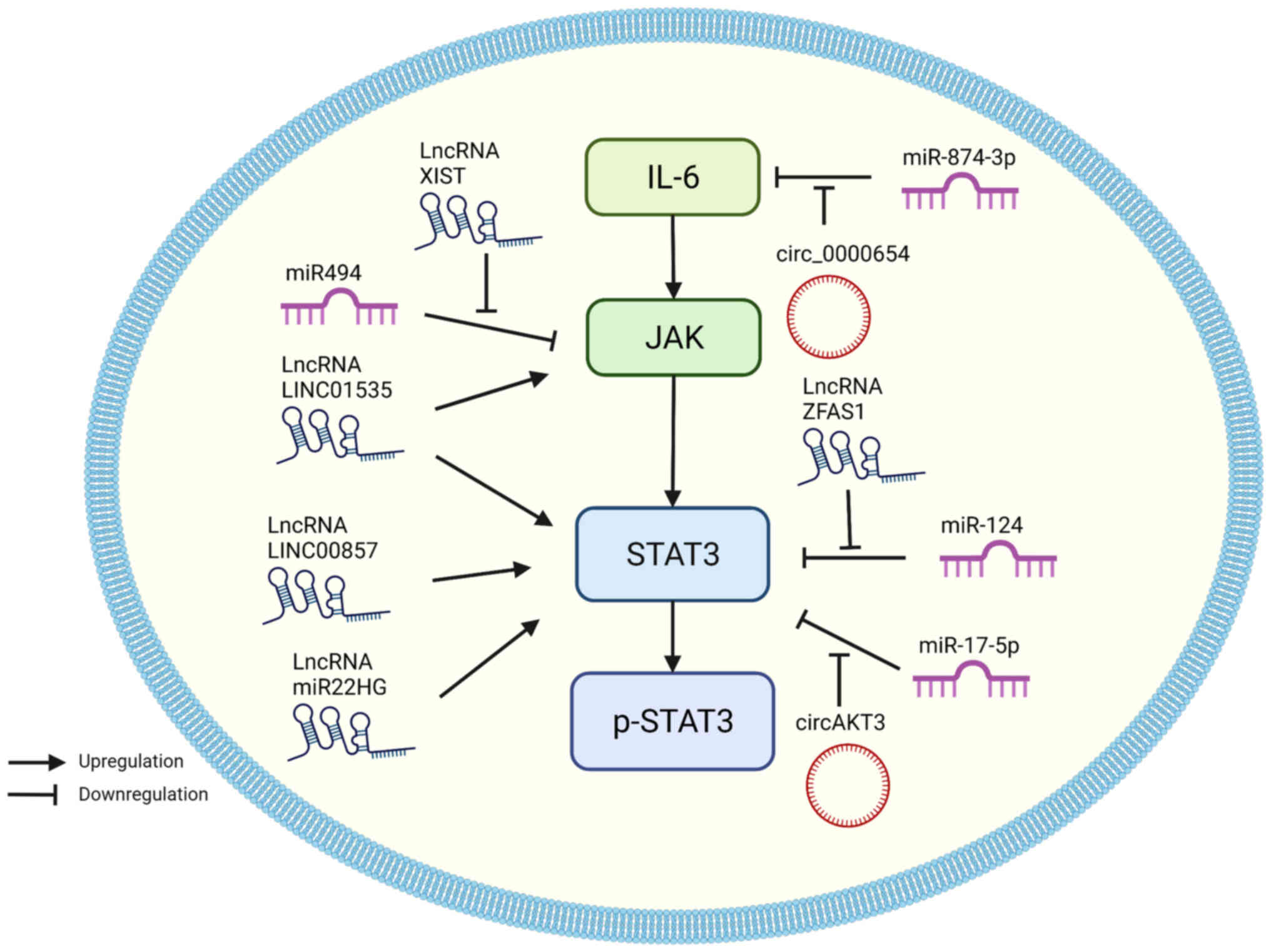
lncRNAs are a type of ncRNA that are >200
nucleotides in length and have been shown to play essential roles
in the epigenetic control and regulation of transcription,
translation, and RNA metabolism (143). The effects of lncRNAs on cancer
progression have attracted considerable research interest for
decades, and several advances have been made recently. In the
research on EC, the effects of lncRNA on STAT3 pathways have been
gradually recognized (Fig. 5;
Table II).
In EC, tumor promotor lncRNAs directly target JAK or
STAT3, including lncRNAs LINC01535, LINC00857, and miR22HG.
LINC01535 is a relatively recently discovered lncRNA of which
little is known regarding its characteristics. Current research has
shown that LINC01535 expression is correlated with the development
of osteosarcoma (144), cervical
cancer (145), and colorectal
cancer (146). In ESCC, LINC01535
expression was upregulated, and this increased proliferation and
prevented apoptosis by activating the JAK/STAT3 pathway (63). LINC00857 is another tumor promoter
lncRNA that can promote tumor progression in lung adenocarcinoma
(147), ovarian cancer (148), and hepatocellular carcinoma
(149). In EAC, LINC00857
expression was shown to be upregulated. After knocking down
LINC00857, STAT3 expression was decreased and the proliferation,
migration, and invasion of cells decreased, showing that LINC00857
affected EAC tumor cell progression by upregulating the expression
of oncoproteins including STAT3 (150).
The mechanism of tumor-suppressing lncRNAs differs
considerably from miRNAs. The expression of the 19-kb lncRNA X
inactivate-specific transcript (XIST) has been reported to be
dysregulated in NSCLC (151),
thyroid cancer (152,153), and colorectal cancer (154). XIST expression is upregulated and
sponges miR-494 in ESCC and EAC cell lines (155). miR-494 is the suppressor of the
JAK/STAT3 pathways (156).
Therefore, XIST can indirectly promote the proliferation, colony
formation, migration, and invasion of EC cells by upregulating
JAK/STAT3 activity (155).
Coincidentally, lncRNA ZFAS1 was shown to exhibit a similar
mechanism, which is closely related to miRNAs and exosomes.
Exosomes assist communication between cancerous cells by delivering
lncRNA ZFAS1 in gastric cancer to promote tumor progression
(157). In ESCC, ZFAS1 was
transmitted to surrounding cells via exosomes, which facilitated
ESCC cell proliferation and migration by downregulating miR-124 and
increasing STAT3 expression (158). Another study confirmed in ESCC
tissues, that ZFAS1 expression was increased and this was
associated with a poorer prognosis (159).
It is worth mentioning the unique role of lncRNA
miR22HG in EC. lncRNA miR22HG primarily functions as a tumor
suppressor in thyroid carcinoma, hepatocellular carcinoma,
endometrial carcinoma, cholangiocarcinoma, colorectal cancer,
gastric cancer, and NSCLC (160).
However, Su et al (161)
showed that silencing miR-22HG expression reduced STAT3 protein
expression and induced apoptosis in the OE33, OE19, and FLO-1 EAC
cell lines, which indicated that miR-22HG functioned as a tumor
promotor in EAC.
circRNAs have a closed-loop structure and play a
role in gene regulation by sponging miRNAs and interacting with RNA
binding proteins. Upregulated expression of circRNAs in tumor
tissues and cell lines is involved in modulating the tumor cell
malignant phenotypes (Fig. 5;
Table II).
circRNA AKT3 (circAKT3) originates from the AKT3
gene and promotes cell proliferation, survival, and drug resistance
(162,163). In EC, circAKT3 expression was
shown to be upregulated and it sponged miR-17-5p. miR-17-5p
directly targets STAT3 and Ras homolog gene family member C (RHOC).
Similar to STAT3, RHOC plays a key role in tumor invasion and
metastasis (164). circAKT3
increases EC cell proliferation, migration, and invasion by
decreasing miR-17-5p activity and indirectly facilitating STAT3 and
RHOC activity. circAKT3 knockdown decreased EC tumor growth in
vivo, suggesting that it could be a target of further study to
improve our understanding of the underlying processes in the
development and progression of EC (165).
Another circRNA, circ_0000654 acts as a tumor
promoter and was shown to be highly expressed in ESCC tissues and
cells. In EC cells, circ_0000654 was demonstrated as an inhibitor
of miR-145-5p, and miR-145-5p targeted IL-6 to reduce STAT3
expression. Therefore, by downregulating miR-145-5p activity,
circ_0000654 could indirectly affect the IL-6/STAT3 signaling
pathway and enhance cell proliferation, migration, and invasion
(166). The roles of various
circRNAs have been confirmed in several types of cancers; however,
the current body of literature regarding circRNAs in EC is still in
its infancy compared to the other types of ncRNAs. The application
of circRNAs in treating EC may have considerable potential and
should thus be further explored.
Studies have focused on regulating and targeting
STAT3 as a potential therapeutic approach, as STAT3 has been linked
to EC cell proliferation and malignancy, and considerable progress
has been made in this field (Fig.
6; Table III).
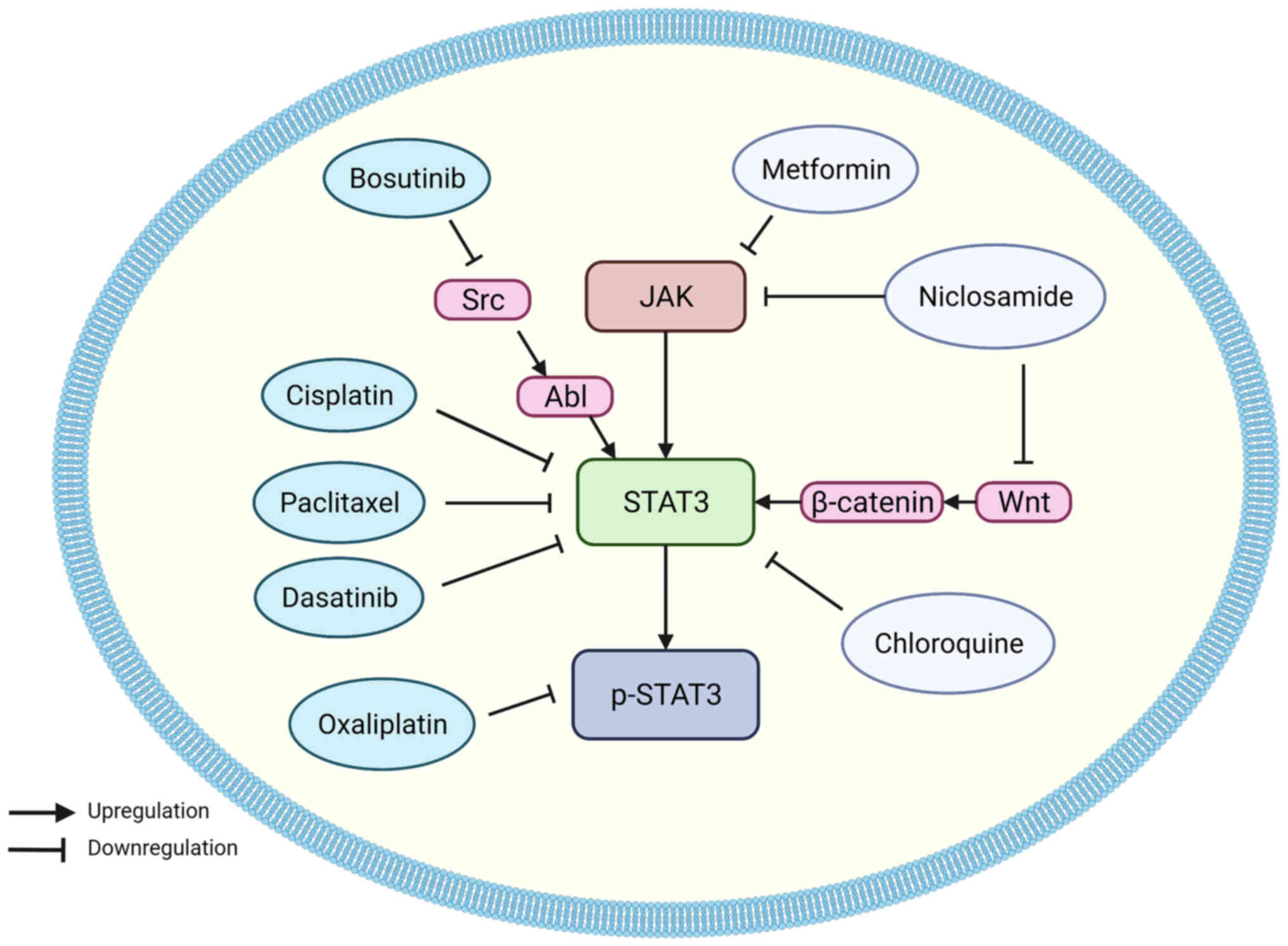
Several canonical anticancer drugs have been shown
to be closely related to STAT3 regulation. Platinum-based
chemotherapeutic drugs have been widely used for their broad
spectrum of antitumor activity (167). There are three generations of
platinum-based drugs, the first generation: Cisplatin, is effective
against a variety of solid tumors, including lung, ovarian, and
testicular cancer (168).
Cisplatin is most commonly combined with 5-FU for the treatment of
EC. However, the therapeutic effects are not satisfactory (169). In a study by Zhao et al
(88), cisplatin was shown to
inhibit proliferation, migration, invasion, and EMT, whilst
inducing apoptosis in ESCC cells by downregulating STAT3, p-STAT3,
and VEGF levels, promoting E-cadherin expression, and suppressing
N-cadherin and vimentin expression. miR-125a-5p increased the
cytotoxic effects of cisplatin, while IL-6 attenuated it.
Chemoresistance is the most common cause of chemotherapy failure,
and this study provides a novel viewpoint for explaining and
solving the problems of drug resistance in EC.
Oxaliplatin belongs to the third generation of
platinum-based antitumor agents. Ngan et al (170) found that oxaliplatin could
decrease the levels of p-STAT3 and reduce the viability of ESCC
cells, whilst also inducing apoptosis of tumor cells. Survivin
belongs to the inhibitor of apoptosis family of proteins; it plays
a significant role in cell division and inhibits apoptosis
(169), and its expression is
upregulated in EC (171). At the
transcriptional level, survivin is one of the downstream targets of
STAT3 in breast cancer cells (172). Oxaliplatin can reduce survivin
levels in EAC cells by altering the STAT3 signaling pathway,
causing EAC cell apoptosis.
Paclitaxel is a cytotoxic drug for treating a
variety of malignancies such as breast cancer, NSCLC, and ovarian
cancer (173). A recent study
revealed that neoadjuvant concurrent chemoradiotherapy with
paclitaxel and carboplatin is associated with improved survival
rates, higher surgical resection rates, and better safety profiles
than the combination of cisplatin and 5-FU for treating locally
advanced EC (174). Paclitaxel
reduced mitochondrial respiration in ESCC cells by downregulating
STAT3 and p-STAT3 expression, resulting in depolarization of the
mitochondrial membrane potential and significantly increasing the
reactive oxygen species (ROS) levels (175).
Using targeted therapies, tyrosine kinase
inhibitors have seen widespread adoption. Dasatinib, a short-acting
tyrosine kinase inhibitor, has become the first-line treatment for
chronic myeloid leukemia in Philadelphia chromosome-positive
patients (176). Several studies
have indicated that the antitumor effects of dasatinib in renal
cell carcinoma (177), pancreatic
cancer (178), and NSCLC
(179) are closely related to
STAT3. Chen et al (180)
found that dasatinib could improve ESCC cisplatin sensitivity by
inhibiting the PI3K/AKT and STAT3 pathways. Bosutinib (SKI-606) is
a second-generation tyrosine kinase inhibitor approved for the
treatment of chronic myeloid leukemia and it selectively inhibits
the kinase activity of Src (181). STAT3 was shown as a downstream
target of Src (46,182), thus it can be speculated that
bosutinib may also have an antitumor effect in EC. Ha et al
(183) showed that bosutinib
induced apoptosis of ESCC by inhibiting Src/Abl and its downstream
JAK/STAT3 signaling pathway, and this increased the cytotoxic
effects of doxorubicin on ESCC cells.
Recently, several non-chemotherapeutic drugs have
been found to possess anticancer activity, including metformin,
niclosamide, and chloroquine (CQ). Metformin is one of the most
commonly used drugs for treating type 2 diabetes (184). Recent studies have shown the
chemopreventative and antineoplastic effects of metformin in
various types of malignancies, including bone cancer, breast
cancer, melanoma, endometrial cancer, and colorectal cancer
(185). The mechanism underlying
the anticancer effects of metformin is also closely related to
STAT3. Metformin was demonstrated to suppress the COX2/PGE2/STAT3
axis and inhibit EMT in prostate cancer (56). Similar effects were confirmed in
ESCC; Feng et al (186)
showed that metformin promoted autophagy and apoptosis in ESCC, and
downregulated STAT3 signaling and its downstream protein Bcl-2. In
a further study, Yang et al (78) emphasized the anti-angiogenic
effects of metformin in ESCC. Metformin reduced ESCC tumor
angiogenesis in vitro and in vivo by suppressing the
JAK/STAT3/c-Myc pathway. Since the pharmacological effects of
metformin and the adverse reactions have been defined previously,
metformin may be a promising therapeutic option for the management
of EC.
Niclosamide is a well-tolerated anthelmintic drug
used to treat cestodes. Several studies have consistently
identified niclosamide as a possible antitumor drug (187,188). Liu et al (73) found that nimesulide decreased COX2
by suppressing JAK2 and STAT3 phosphorylation in ECA-109 cells,
thus mediating tumor cell apoptosis and growth inhibition in
vitro and in vivo. In EC cells, Wei et al
(189) discovered that
niclosamide inhibited the Wnt/β-catenin, STAT3, and mTORC1 pathways
and that it was more efficient in suppressing EC cell activity than
normal cells.
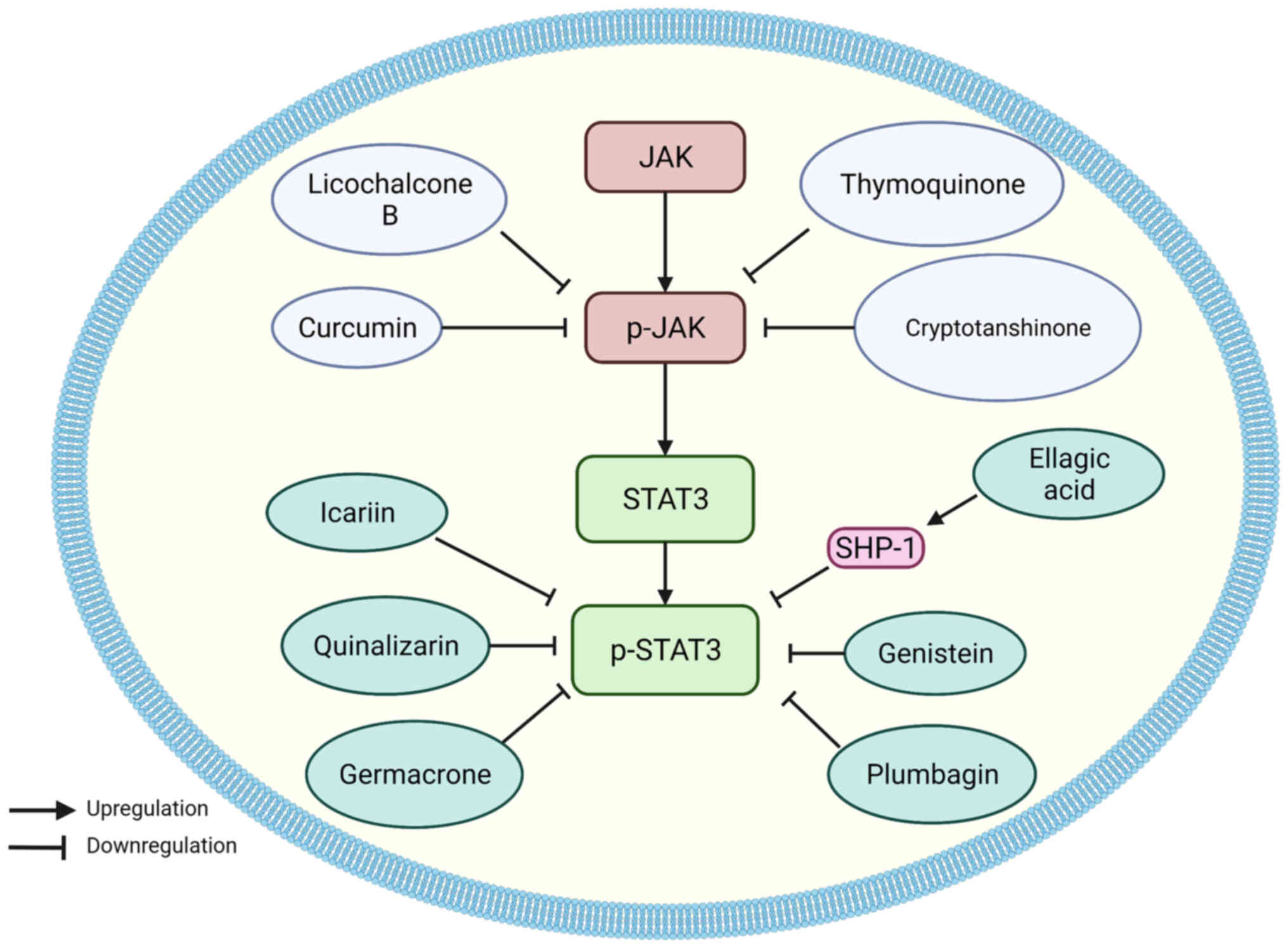
Numerous natural compounds extracted from herbal
medicines exhibit anticancer activity, and rapid progress
concerning the mechanisms of natural compounds has been made in
recent years. The natural compounds that regulate the STAT3
signaling pathway can be divided into two types: JAK inhibitors and
STAT3 inhibitors (Fig. 7; Table IV).
All the natural JAK inhibitors in EC play
anticancer roles by suppressing the activation of JAK. Curcumin
(diferuloylmethane), a naturally occurring substance found in
Curcuma longa, has been shown to reduce tumor proliferation
and suppress a variety of signaling pathways, including prostate
cancer, head and neck squamous cell carcinoma, lung cancer, breast
cancer, and brain tumors (191).
In EC, curcumin downregulated p-JAK, p-STAT3, and total STAT3
expression, and improved cell-cell and cell-matrix adhesion in
ECA-109 cells (192). According
to another study, curcumin was demonstrated to enhance cell cycle
arrest and induce cell death in ESCC in vitro and in
vivo (193). Additionally,
one of the curcumin analogs, 2-pyridyl cyclohexanone, also exhibits
similar functions. Through the JAK2/STAT3 pathway, 2-pyridyl
cyclohexanone suppressed the development of EC cells by triggering
apoptosis in a dose-dependent manner (194).
Cryptotanshinone is a fat-soluble diterpenoid
anthraquinone compound that primarily exists in the plants of the
genus Salvia (197). In
ESCC, cryptotanshinone decreased phosphorylation of JAK2 and STAT3,
inhibited proliferation and migration in vitro, and
inhibited tumor growth in vivo (198).
Some natural compounds can directly target STAT3.
Quinalizarin, also known as 1,2,5,8-tetrahydroquinone, is a dual
anthraquinone compound derived from the roots of the
Rubiaceae herb (201).
Quinalizarin is a compound found in several herbal remedies that
have anti-tumor properties. Zang et al (202) determined that quinalizarin
induced apoptosis and G0/G1 cell cycle arrest of HCE-4 cells via
inhibition of the NF-κB, MAPK, and STAT3 signaling pathways, whilst
also decreasing the levels of cyclin D1/E and CDK2/4, and
increasing the intracellular ROS levels.
Plumbagin, a natural naphthoquinone ingredient
derived from the roots of the medicinal plant Plumbago zeylanica
L., has been shown to possess anticancer effects in a range of
cancer cell lines, including leukemia, breast cancer, melanoma, and
liver cancer (203) In ESCC
cells, Cao et al (66,67)
confirmed that plumbagin downregulated STAT3 levels and arrested
cells in the G0/G1 phase.
Genistein is a simple isoflavone derived from
soybeans and is associated with a reduced risk of prostate, breast,
and lung cancer (204). It can
significantly decrease EGFR expression and STAT3 phosphorylation in
ESCC cells, thus inhibiting STAT3 nuclear translocation, thereby
suppressing the activity of the STAT3 signaling pathway (62). As a result, genistein was
demonstrated to inhibit tumor proliferation in vitro and
in vivo, halt the cell cycle in the G0/G1 phase, and
downregulate the expression of cell cycle-related genes: Cyclin D1,
Bcl-2, Bcl-xl, CDK4, and CDK6, induce apoptosis and upregulate the
expression of apoptosis-associated genes including Bax, Bid, PARP,
caspase-3, and p53.
Germacrone is a natural compound isolated from
Zingiberaceae that has been shown to possess anticancer activity in
breast, brain, liver, skin, prostate, and gastric cancer (205). In ESCC, germacrone was revealed
to inhibit the phosphorylation of STAT3 and increase the Bax/Bcl-2
ratio, thus triggering apoptosis and preventing cell migration
(65).
Ellagic acid is a natural phenol with antioxidant
and antiproliferative effects (208), which is an indirect STAT3
regulator in EC cells. In an in vitro study, Xu et al
(69) found that ellagic acid
suppressed the activation of STAT3 in ESCC cell lines via
upregulation of the levels of SHP-1. SHP-1 is a negative regulator
of STAT3. Therefore, ellagic acid could induce apoptosis in EC
cells.
In the present review, pertinent research on the
STAT3 signaling pathway in EC was summarized. The STAT3 signaling
pathway, which is functionally expressed in several types of cancer
including EC, is a well-known oncogene. STAT3 can be activated by
cytokines or growth factors via several routes, regulating the
transcription of downstream genes. Continuous activation of STAT3
plays a particularly important role in the proliferation,
infiltration, metastasis, angiogenesis, immune evasion,
chemoresistance, and radio-resistance of EC, and is associated with
a poor prognosis in patients, and may thus be considered a
diagnostic and prognostic factor in EC. A wide range of ncRNAs,
including miRNAs, lncRNAs, and circRNAs, can module STAT3 in EC;
some play a promoting role, while others are the inhibitors of
STAT3. Finally, the effects of antitumor drugs and natural
compounds for blocking the STAT3 signaling pathway in EC were
examined, which are worthy of further research in EC therapy.
Not applicable.
ZGS and NZ designed the review. RJM wrote the
manuscript. MMZ and CM prepared the figures and tables. KH revised
the manuscript. Data authentication is not applicable. All the
authors contributed to manuscript revision, as well as read and
approved the submitted version.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
The present review was supported by the Shandong Provincial
Natural Science Foundation (grant no. ZR2020MH204), the 19th Batch
of Science and Technology Innovation Development Plan of Jinan in
2020 (Clinical Medicine Science and Technology Innovation plan;
grant no. 202019032), and the Second Group of Science and
Technology Projects of Jinan Municipal Health Commission (grant no.
2020-3-15).
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Malhotra GK, Yanala U, Ravipati A, Follet
M, Vijayakumar M and Are C: Global trends in esophageal cancer. J
Surg Oncol. 115:564–579. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang FL and Yu SJ: Esophageal cancer:
Risk factors, genetic association, and treatment. Asian J Surg.
41:210–215. 2018. View Article : Google Scholar
|
|
4
|
Arnold M, Ferlay J, van Berge Henegouwen
MI and Soerjomataram I: Global burden of oesophageal and gastric
cancer by histology and subsite in 2018. Gut. 69:1564–1571. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maret-Ouda J, Markar SR and Lagergren J:
Gastroesophageal reflux disease: A review. JAMA. 324:2536–2547.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schlottmann F, Molena D and Patti MG:
Gastroesophageal reflux and Barrett's esophagus: A pathway to
esophageal adenocarcinoma. Updates Surg. 70:339–342. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thrift AP, Shaheen NJ, Gammon MD,
Bernstein L, Reid BJ, Onstad L, Risch HA, Liu G, Bird NC, Wu AH, et
al: Obesity and risk of esophageal adenocarcinoma and Barrett's
esophagus: A Mendelian randomization study. J Natl Cancer Inst.
106:dju2522014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oze I, Matsuo K, Ito H, Wakai K, Nagata C,
Mizoue T, Tanaka K, Tsuji I, Tamakoshi A, Sasazuki S, et al:
Cigarette smoking and esophageal cancer risk: An evaluation based
on a systematic review of epidemiologic evidence among the Japanese
population. Jpn J Clin Oncol. 42:63–73. 2012. View Article : Google Scholar
|
|
9
|
Katada C, Yokoyama T, Yano T, Kaneko K,
Oda I, Shimizu Y, Doyama H, Koike T, Takizawa K, Hirao M, et al:
Alcohol consumption and multiple dysplastic lesions increase risk
of squamous cell carcinoma in the esophagus, head, and neck.
Gastroenterology. 151:860–869.e7. 2016. View Article : Google Scholar
|
|
10
|
Mönig S, Chevallay M, Niclauss N, Zilli T,
Fang W, Bansal A and Hoeppner J: Early esophageal cancer: The
significance of surgery, endoscopy, and chemoradiation. Ann N Y
Acad Sci. 1434:115–123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Horn SR, Stoltzfus KC, Lehrer EJ, Dawson
LA, Tchelebi L, Gusani NJ, Sharma NK, Chen H, Trifiletti DM and
Zaorsky NG: Epidemiology of liver metastases. Cancer Epidemiol.
67:1017602020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi N, Kohno T, Haruta S, Fujimori
S, Shinohara H, Ueno M and Udagawa H: Pulmonary metastasectomy
secondary to esophageal carcinoma: Long-term survival and
prognostic factors. Ann Surg Oncol. 21(Suppl 3): S365–S369. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Ma W, Wu H, Wang J, Lin Y, Wang X
and Zhang C: Analysis of homogeneous and heterogeneous factors for
bone metastasis in esophageal cancer. Med Sci Monit. 25:9416–9425.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carballo GB, Honorato JR, de Lopes GPF and
Spohr TCLSE: A highlight on Sonic hedgehog pathway. Cell Commun
Signal. 16:112018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vaillant C and Monard D: SHH pathway and
cerebellar development. Cerebellum. 8:291–301. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Skoda AM, Simovic D, Karin V, Kardum V,
Vranic S and Serman L: The role of the hedgehog signaling pathway
in cancer: A comprehensive review. Bosn J Basic Med Sci. 18:8–20.
2018. View Article : Google Scholar :
|
|
17
|
Wang L, Jin JQ, Zhou Y, Tian Z, Jablons DM
and He B: Gli is activated and promotes epithelial-mesenchymal
transition in human esophageal adenocarcinoma. Oncotarget.
9:853–865. 2017. View Article : Google Scholar
|
|
18
|
Najafi M, Abbaszadegan MR, Rad A, Dastpak
M, Boroumand-Noughabi S and Forghanifard MM: Crosstalk between SHH
and stemness state signaling pathways in esophageal squamous cell
carcinoma. J Cell Commun Signal. 11:147–153. 2017. View Article : Google Scholar :
|
|
19
|
Zhan T, Rindtorff N and Boutros M: Wnt
signaling in cancer. Oncogene. 36:1461–1473. 2017. View Article : Google Scholar :
|
|
20
|
Moghbeli M, Abbaszadegan MR, Golmakani E
and Forghanifard MM: Correlation of Wnt and NOTCH pathways in
esophageal squamous cell carcinoma. J Cell Commun Signal.
10:129–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao D, Wang B and Chen H: RAB11A mediates
the proliferation and motility of esophageal cancer cells via WNT
signaling pathway. Acta Biochim Pol. 67:531–538. 2020.PubMed/NCBI
|
|
22
|
Feng AL, Han X, Meng X, Chen Z, Li Q, Shu
W, Dai H, Zhu J and Yang Z: PRDX2 plays an oncogenic role in
esophageal squamous cell carcinoma via Wnt/β-catenin and AKT
pathways. Clin Transl Oncol. 22:1838–1848. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang X and Wang X: Long non-coding RNA
colon cancer-associated transcript 2 may promote esophageal cancer
growth and metastasis by regulating the Wnt signaling pathway.
Oncol Lett. 18:1745–1754. 2019.PubMed/NCBI
|
|
24
|
Li Y, Li Y and Chen X: NOTCH and
esophageal squamous cell carcinoma. Adv Exp Med Biol. 1287:59–68.
2021. View Article : Google Scholar :
|
|
25
|
Natsuizaka M, Whelan KA, Kagawa S, Tanaka
K, Giroux V, Chandramouleeswaran PM, Long A, Sahu V, Darling DS,
Que J, et al: Interplay between Notch1 and Notch3 promotes EMT and
tumor initiation in squamous cell carcinoma. Nat Commun.
8:17582017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang T, Kee WH, Seow KT, Fung W and Cao
X: The coiled-coil domain of Stat3 is essential for its SH2
domain-mediated receptor binding and subsequent activation induced
by epidermal growth factor and interleukin-6. Mol Cell Biol.
20:7132–7139. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma J, Zhang T, Novotny-Diermayr V, Tan AL
and Cao X: A novel sequence in the coiled-coil domain of Stat3
essential for its nuclear translocation. J Biol Chem.
278:29252–29260. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu T, Yeh JE, Pinello L, Jacob J,
Chakravarthy S, Yuan GC, Chopra R and Frank DA: Impact of the
N-terminal domain of STAT3 in STAT3-dependent transcriptional
activity. Mol Cell Biol. 35:3284–3300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Verhoeven Y, Tilborghs S, Jacobs J, De
Waele J, Quatannens D, Deben C, Prenen H, Pauwels P, Trinh XB,
Wouters A, et al: The potential and controversy of targeting STAT
family members in cancer. Semin Cancer Biol. 60:41–56. 2020.
View Article : Google Scholar
|
|
30
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: New and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guanizo AC, Fernando CD, Garama DJ and
Gough DJ: STAT3: A multifaceted oncoprotein. Growth Factors.
36:1–14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lim CP and Cao X: Structure, function, and
regulation of STAT proteins. Mol Biosyst. 2:536–550. 2006.
View Article : Google Scholar
|
|
33
|
Sgrignani J, Garofalo M, Matkovic M,
Merulla J, Catapano CV and Cavalli A: Structural biology of STAT3
and Its implications for anticancer therapies development. Int J
Mol Sci. 19:15912018. View Article : Google Scholar :
|
|
34
|
Hevehan DL, Miller WM and Papoutsakis ET:
Differential expression and phosphorylation of distinct STAT3
proteins during granulocytic differentiation. Blood. 99:1627–1637.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kato T, Sakamoto E, Kutsuna H, Kimura-Eto
A, Hato F and Kitagawa S: Proteolytic conversion of STAT3alpha to
STAT3gamma in human neutrophils: role of granule-derived serine
proteases. J Biol Chem. 279:31076–31080. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Johnson DE, O'Keefe RA and Grandis JR:
Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev
Clin Oncol. 15:234–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen M, Chen P, Lu MS, Lin PY, Chen W and
Lee K: IL-6 expression predicts treatment response and outcome in
squamous cell carcinoma of the esophagus. Mol Cancer. 12:262013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Karakasheva TA, Lin EW, Tang Q, Qiao E,
Waldron TJ, Soni M, Klein-Szanto AJ, Sahu V, Basu D, Ohashi S, et
al: IL-6 mediates cross-talk between tumor cells and activated
fibroblasts in the tumor microenvironment. Cancer Res.
78:4957–4970. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Somasundar P, Yu AK, Vona-Davis L and
McFadden DW: Differential effects of leptin on cancer in vitro. J
Surg Res. 113:50–55. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Beales ILP, Garcia-Morales C, Ogunwobi OO
and Mutungi G: Adiponectin inhibits leptin-induced oncogenic
signalling in oesophageal cancer cells by activation of PTP1B. Mol
Cell Endocrinol. 382:150–158. 2014. View Article : Google Scholar
|
|
41
|
Murata T, Asanuma K, Ara N, Iijima K,
Hatta W, Hamada S, Asano N, Koike T, Imatani A, Masamune A and
Shimosegawa T: Leptin aggravates reflux esophagitis by increasing
tissue levels of macrophage migration inhibitory factor in rats.
Tohoku J Exp Med. 245:45–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Andl CD, Mizushima T, Oyama K, Bowser M,
Nakagawa H and Rustgi AK: EGFR-induced cell migration is mediated
predominantly by the JAK-STAT pathway in primary esophageal
keratinocytes. Am J Physiol Gastrointest Liver Physiol.
287:G1227–G1237. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bhat AA, Lu H, Soutto M, Capobianco A, Rai
P, Zaika A and El-Rifai W: Exposure of Barrett's and esophageal
adenocarcinoma cells to bile acids activates EGFR-STAT3 signaling
axis via induction of APE1. Oncogene. 37:6011–6024. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vij N, Sharma A, Thakkar M, Sinha S and
Mohan RR: PDGF-driven proliferation, migration, and IL8 chemokine
secretion in human corneal fibroblasts involve JAK2-STAT3 signaling
pathway. Mol Vis. 14:1020–1027. 2008.PubMed/NCBI
|
|
45
|
Wang Y, Liu S, Jiang G, Zhai W, Yang L, Li
M, Chang Z and Zhu B: NOK associates with c-Src and promotes
c-Src-induced STAT3 activation and cell proliferation. Cell Signal.
75:1097622020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jiang Y, Zhang J, Zhao J, Li Z, Chen H,
Qiao Y, Chen X, Liu K and Dong Z: TOPK promotes metastasis of
esophageal squamous cell carcinoma by activating the
Src/GSK3β/STAT3 signaling pathway via γ-catenin. BMC Cancer.
19:12642019. View Article : Google Scholar
|
|
47
|
Zhu J, Luo L, Tian L, Yin S, Ma X, Cheng
S, Tang W, Yu J, Ma W, Zhou X, et al: Aryl hydrocarbon receptor
promotes IL-10 expression in inflammatory macrophages through
Src-STAT3 signaling pathway. Front Immunol. 9:20332018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim H, Suh JM, Hwang ES, Kim DW, Chung HK,
Song JH, Hwang JH, Park KC, Ro HK and Jo EK: Thyrotropin-mediated
repression of class II trans-activator expression in thyroid cells:
Involvement of STAT3 and suppressor of cytokine signaling. J
Immunol. 171:616–627. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Y, Jia Y, Li P, Li H, Xiao D, Wang Y
and Ma X: Reciprocal activation of α5-nAChR and STAT3 in
nicotine-induced human lung cancer cell proliferation. J Genet
Genomics. 44:355–362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lu R, Zhang YG and Sun J: STAT3 activation
in infection and infection-associated cancer. Mol Cell Endocrinol.
451:80–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gao X, Cao Q, Cheng Y, Zhao D, Wang Z,
Yang H, Wu Q, You L, Wang Y and Lin Y: Chronic stress promotes
colitis by disturbing the gut microbiota and triggering immune
system response. Proc Natl Acad Sci USA. 115:E2960–E2969. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kisseleva T, Bhattacharya S, Braunstein J
and Schindler CW: Signaling through the JAK/STAT pathway, recent
advances and future challenges. Gene. 285:1–24. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bromberg JF, Wrzeszczynska MH, Devgan G,
Zhao Y, Pestell RG, Albanese C and Darnell JE Jr: Stat3 as an
oncogene. Cell. 98:295–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xu ZS, Zhang HX, Zhang YL, Liu TT, Ran Y,
Chen LT, Wang YY and Shu HB: PASD1 promotes STAT3 activity and
tumor growth by inhibiting TC45-mediated dephosphorylation of STAT3
in the nucleus. J Mol Cell Biol. 8:221–231. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhu P, Zhou K, Lu S, Bai Y, Qi R and Zhang
S: Modulation of aryl hydrocarbon receptor inhibits esophageal
squamous cell carcinoma progression by repressing COX2/PGE2/STAT3
axis. J Cell Commun Signal. 14:175–192. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tong D, Liu Q, Liu G, Xu J, Lan W, Jiang
Y, Xiao H, Zhang D and Jiang J: Metformin inhibits
castration-induced EMT in prostate cancer by repressing
COX2/PGE2/STAT3 axis. Cancer Lett. 389:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Colozza G and Koo BK: Wnt/β-catenin
signaling: Structure, assembly and endocytosis of the signalosome.
Dev Growth Differ. 63:199–218. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Mizushima T, Nakagawa H, Kamberov YG,
Wilder EL, Klein PS and Rustgi AK: Wnt-1 but not epidermal growth
factor induces beta-catenin/T-cell factor-dependent transcription
in esophageal cancer cells. Cancer Res. 62:277–282. 2002.PubMed/NCBI
|
|
59
|
Yan S, Zhou C, Zhang W, Zhang G, Zhao X,
Yang S, Wang Y, Lu N, Zhu H and Xu N: beta-Catenin/TCF pathway
upregulates STAT3 expression in human esophageal squamous cell
carcinoma. Cancer Lett. 271:85–97. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tuluhong D, Chen T, Wang J, Zeng H, Li H,
Dunzhu W, Li Q and Wang S: FZD2 promotes TGF-β-induced
epithelial-to-mesenchymal transition in breast cancer via
activating notch signaling pathway. Cancer Cell Int. 21:1992021.
View Article : Google Scholar
|
|
61
|
Fu Y, Zheng Q, Mao Y, Jiang X, Chen X, Liu
P, Lv B, Huang T, Yang J, Cheng Y, et al: WNT2-Mediated FZD2
stabilization regulates esophageal cancer metastasis via STAT3
signaling. Front Oncol. 10:11682020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhang Y, Du XL, Wang CJ, Lin DC, Ruan X,
Feng YB, Huo YQ, Peng H, Cui JL, Zhang TT, et al: Reciprocal
activation between PLK1 and Stat3 contributes to survival and
proliferation of esophageal cancer cells. Gastroenterology.
142:521–530.e3. 2012. View Article : Google Scholar
|
|
63
|
Fang Y, Zhang S, Yin J, Shen YX, Wang H,
Chen XS and Tang H: LINC01535 promotes proliferation and inhibits
apoptosis in esophageal squamous cell cancer by activating the
JAK/STAT3 pathway. Eur Rev Med Pharmacol Sci. 24:3694–3700.
2020.PubMed/NCBI
|
|
64
|
Gao J, Xia R, Chen J, Gao J, Luo X, Ke C,
Ren C, Li J and Mi Y: Inhibition of esophageal-carcinoma cell
proliferation by genistein via suppression of JAK1/2STAT3 and
AKT/MDM2/p53 signaling pathways. Aging (Albany NY). 12:6240–6259.
2020. View Article : Google Scholar
|
|
65
|
Zhang R, Hao J, Guo K, Liu W, Yao F, Wu Q,
Liu C, Wang Q and Yang X: Germacrone inhibits cell proliferation
and induces apoptosis in human esophageal squamous cell carcinoma
cells. Biomed Res Int. 2020:76432482020.PubMed/NCBI
|
|
66
|
Cao Y, Yin X, Jia Y, Liu B, Wu S and Shang
M: Plumbagin, a natural naphthoquinone, inhibits the growth of
esophageal squamous cell carcinoma cells through inactivation of
STAT3. Int J Mol Med. 42:1569–1576. 2018.PubMed/NCBI
|
|
67
|
Cao YY, Yu J, Liu TT, Yang KX, Yang LY,
Chen Q, Shi F, Hao JJ, Cai Y and Wang MR: Plumbagin inhibits the
proliferation and survival of esophageal cancer cells by blocking
STAT3-PLK1-AKT signaling. Cell Death Dis. 9:172018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen M, Ye A, Wei J, Wang R and Poon K:
Deoxycholic acid upregulates the reprogramming factors KFL4 and
OCT4 through the IL-6/STAT3 pathway in esophageal adenocarcinoma
cells. Technol Cancer Res Treat. 19:15330338209453022020.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Xu YY, Wang WW, Huang J and Zhu WG:
Ellagic acid induces esophageal squamous cell carcinoma cell
apoptosis by modulating SHP-1/STAT3 signaling. Kaohsiung J Med Sci.
36:699–704. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Huang Q, Li F, Liu X, Li W, Shi W, Liu FF,
O'Sullivan B, He Z, Peng Y, Tan AC, et al: Caspase 3-mediated
stimulation of tumor cell repopulation during cancer radiotherapy.
Nat Med. 17:860–866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bernard A, Chevrier S, Beltjens F, Dosset
M, Viltard E, Lagrange A, Derangère V, Oudot A, Ghiringhelli F,
Collin B, et al: Cleaved caspase-3 transcriptionally regulates
angiogenesis-promoting chemotherapy resistance. Cancer Res.
79:5958–5970. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Silva KA, Dong J, Dong Y, Dong Y, Schor N,
Tweardy DJ, Zhang L and Mitch WE: Inhibition of Stat3 activation
suppresses caspase-3 and the ubiquitin-proteasome system, leading
to preservation of muscle mass in cancer cachexia. J Biol Chem.
290:11177–11187. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu JR, Wu WJ, Liu SX, Zuo LF, Wang Y,
Yang JZ and Nan YM: Nimesulide inhibits the growth of human
esophageal carcinoma cells by inactivating the JAK2/STAT3 pathway.
Pathol Res Pract. 211:426–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mills JN, Rutkovsky AC and Giordano A:
Mechanisms of resistance in estrogen receptor positive breast
cancer: Overcoming resistance to tamoxifen/aromatase inhibitors.
Curr Opin Pharmacol. 41:59–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Itami A, Shimada Y, Watanabe G and Imamura
M: Prognostic value of p27(Kip1) and CyclinD1 expression in
esophageal cancer. Oncology. 57:311–317. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhang N, Zhang M, Wang Z, Gao W and Sun Z:
Activated STAT3 could reduce survival in patients with esophageal
squamous cell carcinoma by up-regulating VEGF and cyclin D1
expression. J Cancer. 11:1859–1868. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Li H, Xiao W, Ma J, Zhang Y, Li R, Ye J,
Wang X, Zhong X and Wang S: Dual high expression of STAT3 and
cyclinD1 is associated with poor prognosis after curative resection
of esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
7:7989–7998. 2014.
|
|
78
|
Yang Y, Jin G, Liu H, Liu K, Zhao J, Chen
X, Wang D, Bai R, Li X, Jang Y, et al: Metformin inhibits
esophageal squamous cell carcinoma-induced angiogenesis by
suppressing JAK/STAT3 signaling pathway. Oncotarget. 8:74673–74687.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhu Y, Yuan T, Zhang Y, Shi J, Bai L, Duan
X, Tong R and Zhong L: AR-42: A Pan-HDAC inhibitor with antitumor
and antiangiogenic activities in esophageal squamous cell
carcinoma. Drug Des Devel Ther. 13:4321–4330. 2019. View Article : Google Scholar
|
|
80
|
Lei YY, Feng YF, Zeng B, Zhang W, Xu Q,
Cheng F, Lan J, Luo HH, Zou JY, Chen ZG, et al: Exogenous
H2S promotes cancer progression by activating JAK2/STAT3
signaling pathway in esophageal EC109 cells. Int J Clin Exp Pathol.
11:3247–3256. 2018.
|
|
81
|
Li Y, Ma J, Guo Q, Duan F, Tang F, Zheng
P, Zhao Z and Lu G: Overexpression of MMP-2 and MMP-9 in esophageal
squamous cell carcinoma. Dis Esophagus. 22:664–667. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Xuan X, Li S, Lou X, Zheng X, Li Y, Wang
F, Gao Y, Zhang H, He H and Zeng Q: Stat3 promotes invasion of
esophageal squamous cell carcinoma through up-regulation of MMP2.
Mol Biol Rep. 42:907–915. 2015. View Article : Google Scholar
|
|
83
|
Ou Y, Liu L, Xue L, Zhou W, Zhao Z, Xu B,
Song Y and Zhan Q: TRAP1 shows clinical significance and promotes
cellular migration and invasion through STAT3/MMP2 pathway in human
esophageal squamous cell cancer. J Genet Genomics. 41:529–537.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Huang LSM, Snyder EY and Schooley RT:
Strategies and progress in CXCR4-targeted anti-human
immunodeficiency virus (HIV) therapeutic development. Clin Infect
Dis. 73:919–924. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yue D, Zhang D, Shi X, Liu S, Li A, Wang
D, Qin G, Ping Y, Qiao Y and Chen X: Chloroquine inhibits stemness
of esophageal squamous cell carcinoma cells through targeting
CXCR4-STAT3 pathway. Front Oncol. 10:3112020. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Wu J, Gao FX, Wang C, Qin M, Han F, Xu T,
Hu Z, Long Y, He XM and Deng X: IL-6 and IL-8 secreted by tumour
cells impair the function of NK cells via the STAT3 pathway in
oesophageal squamous cell carcinoma. J Exp Clin Cancer Res.
38:3212019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhu H, Chen X, Chen B, Chen B, Fan J, Song
W, Xie Z, Jiang D, Li Q and Zhou M: Activating transcription factor
4 mediates a multidrug resistance phenotype of esophageal squamous
cell carcinoma cells through transactivation of STAT3 expression.
Cancer Lett. 354:142–152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhao Y, Ma K, Yang S, Zhang X, Wang F,
Zhang X, Liu H and Fan Q: MicroRNA-125a-5p enhances the sensitivity
of esophageal squamous cell carcinoma cells to cisplatin by
suppressing the activation of the STAT3 signaling pathway. Int J
Oncol. 53:644–658. 2018.PubMed/NCBI
|
|
89
|
Lee MC, Chen YK, Hsu YJ and Lin BR:
Niclosamide inhibits the cell proliferation and enhances the
responsiveness of esophageal cancer cells to chemotherapeutic
agents. Oncol Rep. 43:549–561. 2020.PubMed/NCBI
|
|
90
|
Zang C, Liu X, Li B, He Y, Jing S, He Y,
Wu W, Zhang B, Ma S and Dai W: IL-6/STAT3/TWIST inhibition reverses
ionizing radiation-induced EMT and radioresistance in esophageal
squamous carcinoma. Oncotarget. 8:11228–11238. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhang Q, Zhang C, He J, Guo Q, Hu D, Yang
X, Wang J, Kang Y, She R, Wang Z, et al: STAT3 inhibitor stattic
enhances radiosensitivity in esophageal squamous cell carcinoma.
Tumour Biol. 36:2135–2142. 2015. View Article : Google Scholar
|
|
92
|
Liu Z, Wang Y, Shu S, Cai J, Tang C and
Dong Z: Non-coding RNAs in kidney injury and repair. Am J Physiol
Cell Physiol. 317:C177–C188. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Anastasiadou E, Jacob LS and Slack FJ:
Non-coding RNA networks in cancer. Nat Rev Cancer. 18:5–18. 2018.
View Article : Google Scholar
|
|
94
|
Romano G, Veneziano D, Acunzo M and Croce
CM: Small non-coding RNA and cancer. Carcinogenesis. 38:485–491.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Klingenberg M, Matsuda A, Diederichs S and
Patel T: Non-coding RNA in hepatocellular carcinoma: Mechanisms,
biomarkers and therapeutic targets. J Hepatol. 67:603–618. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Simonson B and Das S: MicroRNA
therapeutics: The next magic bullet? Mini Rev Med Chem. 15:467–474.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Luo X, Zhang L, Li M, Zhang W, Leng X,
Zhang F, Zhao Y and Zeng X: The role of miR-125b in T lymphocytes
in the pathogenesis of systemic lupus erythematosus. Clin Exp
Rheumatol. 31:263–271. 2013.PubMed/NCBI
|
|
100
|
Liu LH, Li H, Li JP, Zhong H, Zhang HC,
Chen J and Xiao T: miR-125b suppresses the proliferation and
migration of osteosarcoma cells through down-regulation of STAT3.
Biochem Biophys Res Commun. 416:31–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zhu L, Deng H, Hu J, Huang S, Xiong J and
Deng J: The promising role of miR-296 in human cancer. Pathol Res
Pract. 214:1915–1922. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Wang L, Bo X, Zheng Q, Xiao X, Wu L and Li
B: miR-296 inhibits proliferation and induces apoptosis by
targeting FGFR1 in human hepatocellular carcinoma. FEBS Lett.
590:4252–4262. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
He Z, Yu L, Luo S, Li M, Li J, Li Q, Sun Y
and Wang C: miR-296 inhibits the metastasis and
epithelial-mesenchymal transition of colorectal cancer by targeting
S100A4. BMC Cancer. 17:1402017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang ZZ, Luo YR, Du J, Yu Y, Yang XZ, Cui
YJ and Jin XF: MiR-296-5p inhibits cell invasion and migration of
esophageal squamous cell carcinoma by downregulating STAT3
signaling. Eur Rev Med Pharmacol Sci. 23:5206–5214. 2019.PubMed/NCBI
|
|
105
|
Lui WO, Pourmand N, Patterson BK and Fire
A: Patterns of known and novel small RNAs in human cervical cancer.
Cancer Res. 67:6031–6043. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Sun QH, Yin ZX, Li Z, Tian SB, Wang HC,
Zhang FX, Li LP, Zheng CN and Kong S: miR-874 inhibits gastric
cancer cell proliferation by targeting SPAG9. BMC Cancer.
20:5222020. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Liu WG, Zhuo L, Lu Y, Wang L, Ji YX and
Guo Q: miR-8743p inhibits cell migration through targeting RGS4 in
osteosarcoma. J Gene Med. 22:e32132020. View Article : Google Scholar
|
|
108
|
Dong D, Gong Y, Zhang D, Bao H and Gu G:
miR-874 suppresses the proliferation and metastasis of osteosarcoma
by targeting E2F3. Tumour Biol. 37:6447–6455. 2016. View Article : Google Scholar
|
|
109
|
Li Y, Chen X, Xue W, Liang J and Wang L:
MiR-874 inhibits cell proliferation, migration, and invasion of
glioma cells and correlates with prognosis of glioma patients.
Neuromolecular Med. 23:247–255. 2021. View Article : Google Scholar
|
|
110
|
Xia B, Lin M, Dong W, Chen H, Li B, Zhang
X, Hou Y and Lou G: Upregulation of miR-874-3p and miR-874-5p
inhibits epithelial ovarian cancer malignancy via SIK2. J Biochem
Mol Toxicol. 32:e221682018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Feng X, Xue H, Guo S, Chen Y, Zhang X and
Tang X: MiR-874-3p suppresses cell proliferation and invasion by
targeting ADAM19 in nasopharyngeal carcinoma. Panminerva Med.
63:238–239. 2021. View Article : Google Scholar
|
|
112
|
Jiang B, Li Z, Zhang W, Wang H, Zhi X,
Feng J, Chen Z, Zhu Y, Yang L, Xu H and Xu Z: miR-874 Inhibits cell
proliferation, migration and invasion through targeting aquaporin-3
in gastric cancer. J Gastroenterol. 49:1011–1025. 2014. View Article : Google Scholar
|
|
113
|
Zhao B and Dong AS: MiR-874 inhibits cell
growth and induces apoptosis by targeting STAT3 in human colorectal
cancer cells. Eur Rev Med Pharmacol Sci. 20:269–277.
2016.PubMed/NCBI
|
|
114
|
Bu L, Zhang L, Tian M, Zheng Z, Tang H and
Yang Q: LncRNA MIR210HG facilitates non-small cell lung cancer
progression through directly regulation of miR-874/STAT3 axis. Dose
Response. 18:15593258209180522020. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhang X, Tang J, Zhi X, Xie K, Wang W, Li
Z, Zhu Y, Yang L, Xu H and Xu Z: miR-874 functions as a tumor
suppressor by inhibiting angiogenesis through STAT3/VEGF-A pathway
in gastric cancer. Oncotarget. 6:1605–1617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Yuan RB, Zhang SH, He Y, Zhang XY and
Zhang YB: MiR-874-3p is an independent prognostic factor and
functions as an anti-oncomir in esophageal squamous cell carcinoma
via targeting STAT3. Eur Rev Med Pharmacol Sci. 22:7265–7273.
2018.PubMed/NCBI
|
|
117
|
Zhang Q, Liu S, Zhang J, Ma X, Dong M, Sun
B and Xin Y: Roles and regulatory mechanisms of miR-30b in cancer,
cardiovascular disease, and metabolic disorders (Review). Exp Ther
Med. 21:442021. View Article : Google Scholar
|
|
118
|
Meng L, Wang F, Sun S, Zheng Y, Ding Z,
Sun Y, Li B, Meng Q and Xu M: MicroRNA-30b targets CBX3 and
regulates cell proliferation, apoptosis, and migration in
esophageal squamous cell carcinoma via the JAK2/STAT3 signaling
pathway. Int J Clin Exp Pathol. 10:11828–11837. 2017.PubMed/NCBI
|
|
119
|
van Wijnen AJ, Bagheri L, Badreldin AA,
Larson AN, Dudakovic A, Thaler R, Paradise CR and Wu Z: Biological
functions of chromobox (CBX) proteins in stem cell self-renewal,
lineage-commitment, cancer and development. Bone. 143:1156592021.
View Article : Google Scholar
|
|
120
|
Xu J, Lv H, Zhang B, Xu F, Zhu H, Chen B,
Zhu C and Shen J: miR-30b-5p acts as a tumor suppressor microRNA in
esophageal squamous cell carcinoma. J Thorac Dis. 11:3015–3029.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Li B, Xie Z, Li Z, Chen S and Li B:
MicroRNA-613 targets FMNL2 and suppresses progression of colorectal
cancer. Am J Transl Res. 8:5475–5484. 2016.
|
|
122
|
Lu Y, Tang L, Zhang Q, Zhang Z and Wei W:
MicroRNA-613 inhibits the progression of gastric cancer by
targeting CDK9. Artif Cells Nanomed Biotechnol. 46:980–984. 2018.
View Article : Google Scholar
|
|
123
|
Luo J, Jin Y, Li M and Dong L: Tumor
suppressor miR-613 induces cisplatin sensitivity in non-small cell
lung cancer cells by targeting GJA1. Mol Med Rep. 23:3852021.
|
|
124
|
Jiang X, Wu J, Zhang Y, Wang S, Yu X, Li R
and Huang X: MiR-613 functions as tumor suppressor in
hepatocellular carcinoma by targeting YWHAZ. Gene. 659:168–174.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wang W, Zhang H, Wang L, Zhang S and Tang
M: miR-613 inhibits the growth and invasiveness of human
hepatocellular carcinoma via targeting DCLK1. Biochem Biophys Res
Commun. 473:987–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wang J, Yang S, Ge W, Wang Y, Han C and Li
M: MiR-613 suppressed the laryngeal squamous cell carcinoma
progression through regulating PDK1. J Cell Biochem. 119:5118–5125.
2018. View Article : Google Scholar
|
|
127
|
Yang HC, Wu YH, Yen WC, Liu HY, Hwang TL,
Stern A and Chiu DT: The redox role of G6PD in cell growth, cell
death, and cancer. Cells. 8:10552019. View Article : Google Scholar :
|
|
128
|
Su X, Gao C, Feng X and Jiang M: miR-613
suppresses migration and invasion in esophageal squamous cell
carcinoma via the targeting of G6PD. Exp Ther Med. 19:3081–3089.
2020.PubMed/NCBI
|
|
129
|
Li Z, Zhao S, Wang H, Zhang B and Zhang P:
miR-4286 promotes prostate cancer progression via targeting the
expression of SALL1. J Gene Med. e31272019.Epub ahead of print.
PubMed/NCBI
|
|
130
|
Tsai CC, Chen TY, Tsai KJ, Lin MW, Hsu CY,
Wu DC, Tsai EM and Hsieh TH: NF-κB/miR-18a-3p and miR-4286/BZRAP1
axis may mediate carcinogenesis in Helicobacter pylori-Associated
gastric cancer. Biomed Pharmacother. 132:1108692020. View Article : Google Scholar
|
|
131
|
An X, Ge J, Guo H, Mi H, Zhou J, Liu Y,
Weiyue and Wu Z: Overexpression of miR-4286 is an unfavorable
prognostic marker in individuals with non-small cell lung cancer. J
Cell Biochem. 120:17573–17583. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ho KH, Chen PH, Shih CM, Lee YT, Cheng CH,
Liu AJ, Lee CC and Chen KC: miR-4286 is involved in connections
between IGF-1 and TGF-β signaling for the mesenchymal transition
and invasion by glioblastomas. Cell Mol Neurobiol. 42:791–806.
2020. View Article : Google Scholar
|
|
133
|
Rynkiewicz NK, Liu HJ, Balamatsias D and
Mitchell CA: INPP4A/INPP4B and P-Rex proteins: Related but
different? Adv Biol Regul. 52:265–279. 2012. View Article : Google Scholar
|
|
134
|
Zhang M, Tian H, Li R, Yan W and Xu R:
MicroRNA-4286 promotes esophageal carcinoma development by
targeting INPP4A to evoke the JAK2/STAT3 pathway activation.
Pharmazie. 73:342–348. 2018.PubMed/NCBI
|
|
135
|
Ebrahimi F, Gopalan V, Smith RA and Lam
AK: miR-126 in human cancers: Clinical roles and current
perspectives. Exp Mol Pathol. 96:98–107. 2014. View Article : Google Scholar
|
|
136
|
Zhang Z, Wang J, Cheng J and Yu X: Effects
of miR-126 on the STAT3 signaling pathway and the regulation of
malignant behavior in lung cancer cells. Oncol Lett. 15:8412–8416.
2018.PubMed/NCBI
|
|
137
|
Xu J, Wang H, Wang H, Chen Q, Zhang L,
Song C, Zhou Q and Hong Y: The inhibition of miR-126 in cell
migration and invasion of cervical cancer through regulating ZEB1.
Hereditas. 156:112019. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Jiang R, Zhang C, Liu G, Gu R and Wu H:
MicroRNA-126 inhibits proliferation, migration, invasion, and EMT
in osteosarcoma by targeting ZEB1. J Cell Biochem. 118:3765–3774.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Li M, Meng X and Li M: MiR-126 promotes
esophageal squamous cell carcinoma via inhibition of apoptosis and
autophagy. Aging (Albany NY). 12:12107–12118. 2020. View Article : Google Scholar
|
|
140
|
Indrieri A, Carrella S, Carotenuto P,
Banfi S and Franco B: The pervasive role of the miR-181 family in
development, neurodegeneration, and cancer. Int J Mol Sci.
21:20922020. View Article : Google Scholar :
|
|
141
|
He Y, Yang Y, Xu J, Liao Y, Liu L, Deng L
and Xiong X: IL22 drives cutaneous melanoma cell proliferation,
migration and invasion through activation of miR-181/STAT3/AKT
axis. J Cancer. 11:2679–2687. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Xu DD, Zhou PJ, Wang Y, Zhang L, Fu WY,
Ruan BB, Xu HP, Hu CZ, Tian L, Qin JH, et al: Reciprocal activation
between STAT3 and miR-181b regulates the proliferation of
esophageal cancer stem-like cells via the CYLD pathway. Mol Cancer.
15:402016. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Qian X, Zhao J, Yeung PY, Zhang QC and
Kwok CK: Revealing lncRNA structures and interactions by
sequencing-based approaches. Trends Biochem Sci. 44:33–52. 2019.
View Article : Google Scholar
|
|
144
|
Yao X, Wu L, Gu Z and Li J: LINC01535
promotes the development of osteosarcoma through modulating
miR-214-3p/KCNC4 axis. Cancer Manag Res. 12:5575–5585. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Song H, Liu Y, Jin X, Liu Y, Yang Y, Li L,
Wang X and Li G: Long non-coding RNA LINC01535 promotes cervical
cancer progression via targeting the miR-214/EZH2 feedback loop. J
Cell Mol Med. 23:6098–6111. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Zhao C, Jiang Q, Chen L and Chen W: LncRNA
LINC01535 promotes colorectal cancer development and
chemoresistance by sponging miR-761. Exp Ther Med. 22:6852021.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Wang L, Cao L, Wen C, Li J, Yu G and Liu
C: LncRNA LINC00857 regulates lung adenocarcinoma progression,
apoptosis and glycolysis by targeting miR-1179/SPAG5 axis. Hum
Cell. 33:195–204. 2020. View Article : Google Scholar
|
|
148
|
Lin X, Feng D, Li P and Lv Y: LncRNA
LINC00857 regulates the progression and glycolysis in ovarian
cancer by modulating the Hippo signaling pathway. Cancer Med.
9:8122–8132. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Xia C, Zhang XY, Liu W, Ju M, Ju Y, Bu YZ,
Wang W and Shao H: LINC00857 contributes to hepatocellular
carcinoma malignancy via enhancing epithelial-mesenchymal
transition. J Cell Biochem. Dec 3–2018.Epub ahead of print.
|
|
150
|
Su W, Wang L, Niu F, Zou L, Guo C, Wang Z,
Yang X, Wu J, Lu Y and Zhang J: LINC00857 knockdown inhibits cell
proliferation and induces apoptosis via involving STAT3 and MET
oncogenic proteins in esophageal adenocarcinoma. Aging (Albany NY).
11:2812–2821. 2019. View Article : Google Scholar
|
|
151
|
Liu J, Yao L, Zhang M, Jiang J, Yang M and
Wang Y: Downregulation of LncRNA-XIST inhibited development of
non-small cell lung cancer by activating miR-335/SOD2/ROS signal
pathway mediated pyroptotic cell death. Aging (Albany NY).
11:7830–7846. 2019. View Article : Google Scholar
|
|
152
|
Liu H, Deng H, Zhao Y, Li C and Liang Y:
LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor
growth of thyroid cancer through MET-PI3K-AKT signaling. J Exp Clin
Cancer Res. 37:2792018. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Du YL, Liang Y, Cao Y, Liu L, Li J and Shi
GQ: LncRNA XIST promotes migration and invasion of papillary
thyroid cancer cell by modulating MiR-101-3p/CLDN1 axis. Biochem
Genet. 59:437–452. 2021. View Article : Google Scholar
|
|
154
|
Zheng H, Zhang M, Ke X, Deng X, Li D and
Wang Q, Yan S, Xue Y and Wang Q: LncRNA XIST/miR-137 axis
strengthens chemo-resistance and glycolysis of colorectal cancer
cells by hindering transformation from PKM2 to PKM1. Cancer
Biomark. 30:395–406. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Chen Z, Hu X, Wu Y, Cong L, He X, Lu J,
Feng J and Liu D: Long non-coding RNA XIST promotes the development
of esophageal cancer by sponging miR-494 to regulate CDK6
expression. Biomed Pharmacother. 109:2228–2236. 2019. View Article : Google Scholar
|
|
156
|
Jiang H, Deng W, Zhu K, Zeng Z, Hu B, Zhou
Z, Xie A, Zhang C, Fu B, Zhou X and Wang G: LINC00467 promotes
prostate cancer progression via M2 macrophage polarization and the
miR-494-3p/STAT3 axis. Front Oncol. 11:6614312021. View Article : Google Scholar :
|
|
157
|
Pan L, Liang W, Fu M, Huang ZH, Li X,
Zhang W, Zhang P, Qian H, Jiang PC, Xu WR and Zhang X:
Exosomes-mediated transfer of long noncoding RNA ZFAS1 promotes
gastric cancer progression. J Cancer Res Clin Oncol. 143:991–1004.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Li Z, Qin X, Bian W, Li Y, Shan B, Yao Z
and Li S: Exosomal lncRNA ZFAS1 regulates esophageal squamous cell
carcinoma cell proliferation, invasion, migration and apoptosis via
microRNA-124/STAT3 axis. J Exp Clin Canc Res. 38:4772019.
View Article : Google Scholar
|
|
159
|
Shi H, Liu Z, Pei D, Jiang Y, Zhu H and
Chen B: Development and validation of nomogram based on lncRNA
ZFAS1 for predicting survival in lymph node-negative esophageal
squamous cell carcinoma patients. Oncotarget. 8:59048–59057. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Zhang L, Li C and Su X: Emerging impact of
the long noncoding RNA MIR22HG on proliferation and apoptosis in
multiple human cancers. J Exp Clin Cancer Res. 39:2712020.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Su W, Guo C, Wang L, Wang Z, Yang X, Niu
F, Tzou D, Yang X, Huang X, Wu J, et al: LncRNA MIR22HG abrogation
inhibits proliferation and induces apoptosis in esophageal
adenocarcinoma cells via activation of the STAT3/c-Myc/FAK
signaling. Aging (Albany NY). 11:4587–4596. 2019. View Article : Google Scholar
|
|
162
|
Li F, Zhang L and Sun Q: CircAKT3 promotes
cell proliferation, survival and glutamine metabolism of gastric
cancer by activating SLC1A5 expression via targeting miR-515-5p.
Histol Histopathol. 37:227–241. 2022.
|
|
163
|
Huang X, Li Z, Zhang Q, Wang W, Li B, Wang
L, Xu Z, Zeng A, Zhang X, Zhang X, et al: Circular RNA AKT3
upregulates PIK3R1 to enhance cisplatin resistance in gastric
cancer via miR-198 suppression. Mol Cancer. 18:712019. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Guan X, Chen S and Zhao Y: The role of
RhoC in malignant tumor invasion, metastasis and targeted therapy.
Histol Histopathol. 33:255–260. 2018.
|
|
165
|
Zang HL, Ji FJ, Ju HY and Tian XF:
Circular RNA AKT3 governs malignant behaviors of esophageal cancer
cells by sponging miR-17-5p. World J Gastroentero. 27:240–254.
2021. View Article : Google Scholar
|
|
166
|
Xu Z, Tie X, Li N, Yi Z, Shen F and Zhang
Y: Circular RNA hsa_circ_0000654 promotes esophageal squamous cell
carcinoma progression by regulating the miR-149-5p/IL-6/STAT3
pathway. IUBMB Life. 72:426–439. 2020. View Article : Google Scholar
|
|
167
|
Dilruba S and Kalayda GV: Platinum-based
drugs: Past, present and future. Cancer Chemother Pharmacol.
77:1103–1124. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Ghosh S: Cisplatin: The first metal based
anticancer drug. Bioorg Chem. 88:1029252019. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Kato H and Nakajima M: Treatments for
esophageal cancer: A review. Gen Thorac Cardiovasc Surg.
61:330–335. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Ngan CY, Yamamoto H, Takagi A, Fujie Y,
Takemasa I, Ikeda M, Takahashi-Yanaga F, Sasaguri T, Sekimoto M,
Matsuura N and Monden M: Oxaliplatin induces mitotic catastrophe
and apoptosis in esophageal cancer cells. Cancer Sci. 99:129–139.
2008.
|
|
171
|
Kato J, Kuwabara Y, Mitani M, Shinoda N,
Sato A, Toyama T, Mitsui A, Nishiwaki T, Moriyama S, Kudo J and
Fujii Y: Expression of survivin in esophageal cancer: Correlation
with the prognosis and response to chemotherapy. Int J Cancer.
95:92–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Gritsko T, Williams A, Turkson J, Kaneko
S, Bowman T, Huang M, Nam S, Eweis I, Diaz N, Sullivan D, et al:
Persistent activation of stat3 signaling induces survivin gene
expression and confers resistance to apoptosis in human breast
cancer cells. Clin Cancer Res. 12:11–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Zhu L and Chen L: Progress in research on
paclitaxel and tumor immunotherapy. Cell Mol Biol Lett. 24:402019.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Mayanagi S, Irino T, Kawakubo H and
Kitagawa Y: Neoadjuvant treatment strategy for locally advanced
thoracic esophageal cancer. Ann Gastroenterol Surg. 3:269–275.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Zhang X, Wu X, Zhang F, Mo S, Lu Y, Wei W,
Chen X, Lan L, Lu B and Liu Y: Paclitaxel induces apoptosis of
esophageal squamous cell carcinoma cells by downregulating STAT3
phosphorylation at Ser727. Oncol Rep. 37:2237–2244. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Miura M: Therapeutic drug monitoring of
imatinib, nilotinib, and dasatinib for patients with chronic
myeloid leukemia. Biol Pharm Bull. 38:645–654. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Lue HW, Cole B, Rao SA, Podolak J, Van
Gaest A, King C, Eide CA, Wilmot B, Xue C, Spellman PT, et al: Src
and STAT3 inhibitors synergize to promote tumor inhibition in renal
cell carcinoma. Oncotarget. 6:44675–44687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Ma L, Wei J, Su GH and Lin J: Dasatinib
can enhance paclitaxel and gemcitabine inhibitory activity in human
pancreatic cancer cells. Cancer Biol Ther. 20:855–865. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Wang M and Yuang-Chi Chang A: Molecular
mechanism of action and potential biomarkers of growth inhibition
of synergistic combination of afatinib and dasatinib against
gefitinib-resistant non-small cell lung cancer cells. Oncotarget.
9:16533–16546. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Chen J, Lan T, Zhang W, Dong L, Kang N, Fu
M, Liu B, Liu K, Zhang C, Hou J and Zhan Q: Dasatinib enhances
cisplatin sensitivity in human esophageal squamous cell carcinoma
(ESCC) cells via suppression of PI3K/AKT and Stat3 pathways. Arch
Biochem Biophys. 575:38–45. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Gambacorti-Passerini C, le Coutre P and
Piazza R: The role of bosutinib in the treatment of chronic myeloid
leukemia. Future Oncol. 16:4395–4408. 2020. View Article : Google Scholar
|
|
182
|
Deng B, Jiang XL, Tan ZB, Cai M, Deng SH,
Ding WJ, Xu YC, Wu YT, Zhang SW, Chen RX, et al: Dauricine inhibits
proliferation and promotes death of melanoma cells via inhibition
of Src/STAT3 signaling. Phytother Res. 35:3836–3847. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Ha YNE, Dai Y, Wufuer D, Pidayi M,
Anasihan G and Chen L: Second-generation Src/Abl inhibitor
bosutinib effectively induces apoptosis in human esophageal
squamous cell carcinoma (ESCC) cells via inhibiting Src/Abl
signaling. Neoplasma. 67:54–60. 2020. View Article : Google Scholar
|
|
184
|
Sanchez-Rangel E and Inzucchi SE:
Metformin: Clinical use in type 2 diabetes. Diabetologia.
60:1586–1593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Lv Z and Guo Y: Metformin and its benefits
for various diseases. Front Endocrinol (Lausanne). 11:1912020.
View Article : Google Scholar
|
|
186
|
Feng Y, Ke C, Tang Q, Dong H, Zheng X, Lin
W, Ke J, Huang J, Yeung SC and Zhang H: Metformin promotes
autophagy and apoptosis in esophageal squamous cell carcinoma by
downregulating Stat3 signaling. Cell Death Dis. 5:e10882014.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Chen W, Mook RJ Jr, Premont RT and Wang J:
Niclosamide: Beyond an antihelminthic drug. Cell Signal. 41:89–96.
2018. View Article : Google Scholar
|
|
188
|
Barbosa EJ, Löbenberg R, de Araujo GLB and
Bou-Chacra NA: Niclosamide repositioning for treating cancer:
Challenges and nano-based drug delivery opportunities. Eur J Pharm
Biopharm. 141:58–69. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Wei W, Liu H, Yuan J and Yao Y: Targeting
Wnt/β-catenin by anthelmintic drug niclosamide overcomes paclitaxel
resistance in esophageal cancer. Fund Clin Pharmacol. 35:165–173.
2021. View Article : Google Scholar
|
|
190
|
Ferreira PMP, Sousa RWR, Ferreira JRO,
Militão GCG and Bezerra DP: Chloroquine and hydroxychloroquine in
antitumor therapies based on autophagy-related mechanisms.
Pharmacol Res. 168:1055822021. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Tomeh MA, Hadianamrei R and Zhao X: A
review of curcumin and its derivatives as anticancer agents. Int J
Mol Sci. 20:10332019. View Article : Google Scholar :
|
|
192
|
Zheng BZ, Liu TD, Chen G, Zhang JX and
Kang X: The effect of curcumin on cell adhesion of human esophageal
cancer cell. Eur Rev Med Pharmacol Sci. 22:551–560. 2018.PubMed/NCBI
|
|
193
|
Liu Y, Wang X, Zeng S, Zhang X, Zhao J,
Zhang X, Chen X, Yang W, Yang Y, Dong Z, et al: The natural
polyphenol curcumin induces apoptosis by suppressing STAT3
signaling in esophageal squamous cell carcinoma. J Exp Clin Canc
Res. 37:3032018. View Article : Google Scholar
|
|
194
|
Wang Y, Zhou P, Qin S, Xu D, Liu Y, Fu W,
Ruan B, Zhang L, Zhang Y, Wang X, et al: The curcumin analogs
2-Pyridyl cyclohexanone induce apoptosis via inhibition of the
JAK2-STAT3 pathway in human esophageal squamous cell carcinoma
cells. Front Pharmacol. 9:8202018. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Cao Y, Xu W, Huang Y and Zeng X:
Licochalcone B, a chalcone derivative from Glycyrrhiza inflata, as
a multifunctional agent for the treatment of Alzheimer's disease.
Nat Prod Res. 34:736–739. 2020. View Article : Google Scholar
|
|
196
|
Song M, Yoon G, Choi JS, Kim E, Liu X, Oh
HN, Chae JI, Lee MH and Shim JH: Janus kinase 2 inhibition by
licochalcone B suppresses esophageal squamous cell carcinoma
growth. Phytother Res. 34:2032–2043. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Wu YH, Wu YR, Li B and Yan ZY:
Cryptotanshinone: A review of its pharmacology activities and
molecular mechanisms. Fitoterapia. 145:1046332020. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Ji Y, Liu Y, Xue N, Du T, Wang L, Huang R,
Li L, Yan C and Chen X: Cryptotanshinone inhibits esophageal
squamous-cell carcinoma in vitro and in vivo through the
suppression of STAT3 activation. Onco Targets Ther. 12:883–896.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Imran M, Rauf A, Khan IA, Shahbaz M,
Qaisrani TB, Fatmawati S, Abu-Izneid T, Imran A, Rahman KU and
Gondal TA: Thymoquinone: A novel strategy to combat cancer: A
review. Biomed Pharmacother. 106:390–402. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Hu X, Ma J, Vikash V, Li J, Wu D, Liu Y,
Zhang J and Dong W: Thymoquinone augments cisplatin-induced
apoptosis on esophageal carcinoma through mitigating the activation
of JAK2/STAT3 pathway. Digest Dis Sci. 63:126–134. 2018. View Article : Google Scholar
|
|
201
|
Luo YH, Li JQ, Zhang Y, Wang JR, Xu WT,
Zhang Y, Feng YC, Li SZ and Jin CH: Quinalizarin induces cycle
arrest and apoptosis via reactive oxygen species-mediated signaling
pathways in human melanoma A375 cells. Drug Dev Res. 80:1040–1050.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Zang YQ, Zhai YQ, Feng YY, Ju XY and Zuo
F: Molecular mechanisms of quinalizarin induces apoptosis and G0/G1
cell cycle of human esophageal cancer HCE-4 cells depends on MAPK,
STAT3, and NF-κB signaling pathways. Environ Toxicol. 36:276–286.
2021. View Article : Google Scholar
|
|
203
|
Yin Z, Zhang J, Chen L, Guo Q, Yang B,
Zhang W and Kang W: Anticancer effects and mechanisms of action of
plumbagin: Review of research advances. Biomed Res Int.
2020:69409532020. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Thangavel P, Puga-Olguín A,
Rodríguez-Landa JF and Zepeda RC: Genistein as potential
therapeutic candidate for menopausal symptoms and other related
diseases. Molecules. 24:38922019. View Article : Google Scholar :
|
|
205
|
Riaz A, Rasul A, Kanwal N, Hussain G, Shah
MA, Sarfraz I, Ishfaq R, Batool R, Rukhsar F and Adem Ş:
Germacrone: A potent secondary metabolite with therapeutic
potential in metabolic diseases, cancer and viral infections. Curr
Drug Metab. 21:1079–1090. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
He C, Wang Z and Shi J: Pharmacological
effects of icariin. Adv Pharmacol. 87:179–203. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Gu ZF, Zhang ZT, Wang JY and Xu BB:
Icariin exerts inhibitory effects on the growth and metastasis of
KYSE70 human esophageal carcinoma cells via PI3K/AKT and STAT3
pathways. Environ Toxicol Phar. 54:7–13. 2017. View Article : Google Scholar
|
|
208
|
Derosa G, Maffioli P and Sahebkar A:
Ellagic acid and its role in chronic diseases. Adv Exp Med Biol.
928:473–479. 2016. View Article : Google Scholar : PubMed/NCBI
|