As the fourth most common malignant cancer type
worldwide, colorectal cancer (CRC) is the second leading cause of
cancer-related death. Its high morbidity and mortality rates
indicate that this disease has a serious impact on human health,
with the third (10%) and second (9.4%) highest morbidity and
mortality rates, respectively, for all tumors globally in 2020
(1,2). In the past, CRC was mainly treated by
surgery, chemotherapy and radiotherapy, but recurrence and distant
metastasis after treatment remain challenges in the treatment of
CRC (3,4). It has been demonstrated that the
Wnt/β-catenin signaling pathway has an important role in the
development, progression, metastasis and recurrence of CRC
(5-7). In most patients with CRC, the
Wnt/β-catenin signaling pathway has abnormal levels of activation
(8) and blocking the Wnt/β-catenin
signaling pathway may inhibit the development, progression,
invasion and metastasis of CRC. The development of therapeutic
drugs that target the Wnt/β-catenin signaling pathway is of current
interest in the research community. Therefore, the Wnt/β-catenin
signaling pathway is expected to become a target for the treatment
of CRC.
The development and procession of CRC are complex
processes that are related to several individual factors, including
lifestyle, environmental factors and genetics, but the exact
mechanism remains to be fully elucidated, while a previous study
has indicated that individuals with adenomas have a higher risk of
developing CRC (9). Among the
different risk factors, heredity is an independent risk factor for
CRC and patients without a family history only occasionally develop
CRC. Although several CRC-related genetic factors remain elusive,
mutations in genes associated with CRC (such as single nucleotide
polymorphisms) have been successfully identified (10). Thus, more in-depth studies are
required to confirm the genetic mechanisms of CRC. Of note, due to
the insidious early symptoms of CRC, at the time of diagnosis, half
of the patients have already progressed to the advanced stage of
the disease. The prognosis of patients is poor and the 5-year
overall survival (OS) rate is <15% (11,12).
In recent years, with progress in the related fields
of immunology and oncology, members of the T-cell regulator CD28
superfamily, including programmed cell death protein-1 (PD-1),
programmed cell death ligand-1 (PD-L1) and their inhibitors,
represent new targets in strategies for anti-CRC therapy (13). Compared with traditional therapy,
such as chemotherapy and radiotherapy alone, PD-1/PD-LI inhibitors
significantly shorten the time to remission and there is a lower
rate of adverse events following treatment (3,14).
However, patients with tumors that display microsatellite stability
and a low proficiency of mismatch repair, which includes the vast
majority of patients with advanced CRC, do not exhibit any signs of
obvious remission, and most do not respond to immunotherapy
(15,16). Therefore, research on the Wnt
pathway is of practical significance and value, particularly in
light of the current status of therapeutic treatments.
It has been demonstrated that the Wnt signaling
pathway, the transforming growth factor-β (TGF-β) signaling pathway
and the Hedgehog signaling pathway all have a role in colorectal
tumors (17,18). Among them, the Wnt signal
transduction pathway, which leads to tumor invasion, recurrence and
metastasis, is abnormally activated in most patients with CRC and
it has an important role in the development and progression of CRC
(19).
The Wnt protein family is involved in numerous
cellular functions and comprises 19 secreted glycoproteins, which
may transmit extracellular signals intracellularly through cell
surface receptors; therefore, these proteins participate in a wide
variety of biological processes, such as embryonic development and
organ formation, cell proliferation, cell differentiation and stem
cell self-renewal (20,21). The Wnt signaling pathway may be
generally divided into the following two categories: The
β-catenin-dependent canonical pathway and the β-catenin-independent
noncanonical pathway (22).
The Wnt/β-catenin signaling pathway is the classical
pathway of Wnt signaling and its abnormal activation may lead to
the development, progression, invasion, metastasis and recurrence
of different types of tumor, including CRC, in humans (8). The key mechanism underlying the
Wnt-induced promotion of tumor-cell proliferation and neoplasia is
β-catenin protein stabilization and nuclear translocation, which is
mediated by Wnt.
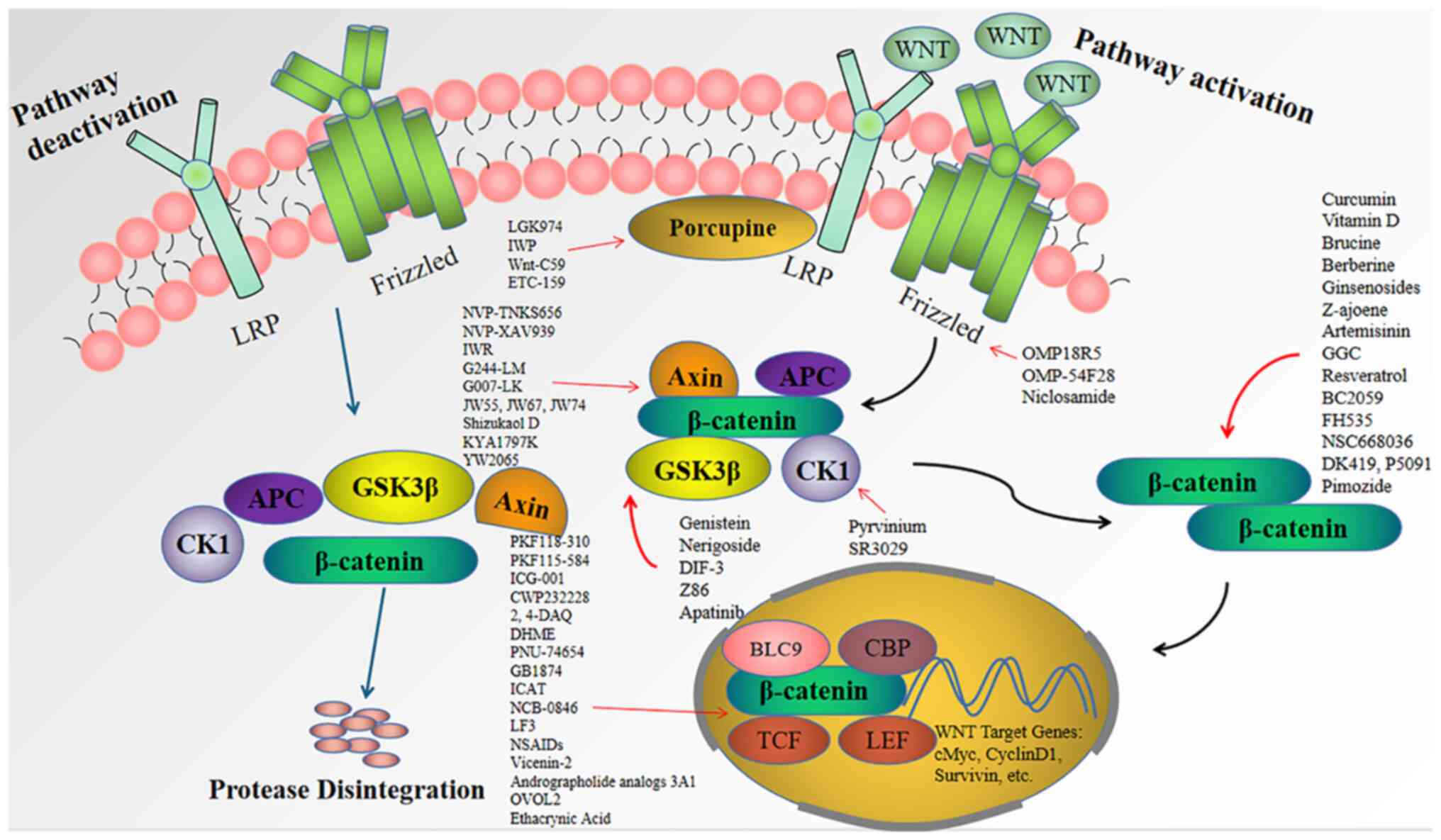
In the absence of Wnt activation, the destruction
complex composed of glycogen synthase kinase 3-β (GSK-3β), casein
kinase 1α (CK1α), adenomatous polyposis coli (APC) and Axin may
enhance the ubiquitination of β-catenin through regulation of the
β-transducin-repeat-containing protein, which mediates ubiquitin
ligase E3 and eventually leads to the absorption and degradation of
β-catenin in the cytoplasm (23).
In the case of Wnt activation, Wnt binds to the transmembrane
Frizzled (FZD) receptor and the low-density lipoprotein
receptor-related protein (LRP)5 and LRP6 coreceptors to trigger the
dissociation of GSK-3β from the destruction complex. At the same
time, Dishevelled (DVL) and Axin accumulate in the cell membrane,
inhibiting the formation of the destruction complex (24,25).
Based on this, β-catenin cannot be degraded and tends to be
stabilized, accumulate in the cytoplasm and translocate to the
nucleus to bind to the T-cell factor/lymphoid enhancer-binding
factor (TCF/LEF) family of transcription factors and coactivators.
This binding induces the transcription of genes important for Wnt
final activity, including c-Myc, survivin, c-Jun, cyclin D1 and
Wnt1-inducible-signaling pathway protein 1 (Wisp1), which leads to
the development and progression of CRC (7,20).
The specific mechanism is as follows: Proto-oncogenes, such as
c-Jun, cyclin D1 and c-Myc, promote cell proliferation,
differentiation and maturation and eventually lead to
tumorigenesis. While survivin is a new member of the apoptosis
suppressor protein family, it is specifically expressed only in
tumor and embryonic tissues, and it promotes tumor cell
differentiation, proliferation, infiltration and metastasis. Wisp1
inhibits the caspase cascade reaction and suppresses apoptosis,
thereby inducing tumorigenesis.
Of note, Wnt binding to FZD-activated DVL causes
calcium to be released from the endoplasmic reticulum and activates
calcium-binding proteins, including protein kinase C and
calcium/calmodulin-dependent protein kinase II, which enhances the
phosphorylation of TCF/LEF (27-29).
This phosphorylation blocks the canonical Wnt pathway and inhibits
the expression of its downstream target genes, which suppresses the
development of colorectal tumors (27-29).
There is also a clear interaction between the canonical and
noncanonical Wnt pathways, wherein the noncanonical pathway may
reduce the occurrence of colorectal tumors by blocking the
canonical pathway (7,26-29).
The maintenance of normal intestinal function and
stem-cell homeostasis requires normal activation of Wnt signaling.
It has been verified that inhibition of the Wnt/β-catenin signaling
pathway may lead to a lack of intestinal stem cells (30), whereas the number of intestinal
stem cells increases at the bottom of intestinal crypts (31), where the activity of the
Wnt/β-catenin signaling pathway is highest (32,33).
These findings indicate that Wnt/β-catenin signaling has a critical
role in the maintenance and self-renewal of intestinal stem cells,
particularly epithelial stem cells at the base of intestinal crypts
(34,35).
It has been indicated that abnormal activation of
the Wnt/β-catenin signaling pathway is a cause of CRC development
and progression, and most patients with CRC have mutations in this
pathway (19). Abnormal activation
of the Wnt/β-catenin pathway may lead to accumulation of the
β-catenin protein in the nucleus, causing massive cell
proliferation (7,36,37).
Initially, this leads to the formation of adenomas, but with the
accumulation of several mutations, adenomas eventually develop into
CRC (7,36). In patients with sporadic CRC, a
β-catenin mutation may be found in 10% of patients, while the rate
of mutation in the APC gene is up to 80% in patients (7,37).
The absorption and degradation of β-catenin is suppressed in
patients that have an APC mutation (7,37,38).
This suppression causes further accumulation of β-catenin in the
cytoplasm and an increase in the amount of it translocating into
the nucleus, both of which contribute to the progression of CRC
(7,36,37).
In addition, the Wnt/β-catenin signaling pathway may induce tumor
resistance and targeted inhibition of the Wnt/β-catenin pathway in
digestive tract tumors may enhance the tumors' sensitivity to
chemotherapeutic drugs (39,40).
Wnt/β-catenin signaling may also induce epithelial-mesenchymal
transition (EMT), which has a vital role in the physiological
processes of intestinal and CRC stem cells (41,42).
More importantly, abnormal activation of the Wnt/β-catenin
signaling pathway and accumulation of β-catenin has been confirmed
in >80% of patients with CRC (43). Therefore, inhibiting Wnt/β-catenin
signaling and the accumulation of β-catenin in the nucleus may
inhibit the development, progression, invasion, metastasis and
recurrence of CRC. Overall, the Wnt/β-catenin signaling pathway may
become an effective target for CRC treatment and studies
investigating how to inhibit this pathway are of practical and
clinical significance.
Studies investigating the targeting of the Wnt
signaling pathway and blocking its signal transduction have
indicated that this pathway may stabilize the formation of the
destruction complex, promote the degradation of β-catenin and
interfere with β-catenin-dependent transcription factors, all of
which hinder the development, progression, invasion, metastasis and
recurrence of colorectal tumors (44). Research aimed at inhibiting the
Wnt/β-catenin signaling pathway may provide treatments that benefit
patients with CRC by improving their quality of life and prolonging
their survival time. Inhibitors of the Wnt/β-catenin signaling
pathway that have been discovered in previous years are summarized
below and are outlined in Table I
and Fig. 1.
The TCF/LEF family is a group of transcription
factors that bind DNA through high mobility group motifs, and they
recruit the coactivator β-catenin protein to enhance the expression
of the genetic elements they target. The typical Wnt signaling
pathway is activated by a combination of β-catenin and TCF/LEF
transcription factors, which then activates target genes that are
downstream of the Wnt pathway, such as cyclin D1 and c-Myc and
other proto-oncogenes, leading to cell proliferation and mediating
the development and progression of tumors (20). Therefore, inhibiting the formation
of the β-catenin-TCF/LEF complex may interrupt the expression of
downstream target genes and block the formation, invasion and
metastasis of colorectal tumors.
The Ras gene has a critical role in the development
and progression of tumors in humans. Clinically, the Kirsten rat
sarcoma viral oncogene (KRAS) mutation is the most common mutation
of the Ras gene and patients with a KRAS mutation have unfavorable
prognosis and a higher level of tumor invasion and metastasis
(52). Approximately half of the
patients with colon cancer express the KRAS mutation, and in CRC
cells expressing activated KRAS, the Wnt/β-catenin signaling
pathway is activated (53).
Furthermore, KRAS mutation disrupts the negative feedback mechanism
of its target microRNA (miR)-139-5p and Wnt signaling, resulting in
downregulation of the tumor suppressor miR-139-5p and promotion of
tumor growth (54). Hence, the
simultaneous downregulation of β-catenin and KRAS is key to
blocking the development and progression of CRC. Mologni et
al (55) indicated that the
combination of PKF115-584 and the KRAS inhibitor S-trans was
superior to any monotherapy in arresting cell growth, causing cell
death and downregulating c-Myc and survivin.
DHME, a chemically synthesized derivative of the
fungal polyphenol hispolon has been indicated to block signaling of
the Wnt canonical pathway by inhibiting the transcriptional
activity of the β-catenin/TCF complex. It exhibits strong
apoptosis-inducing activity and has a significant role in blocking
the development and progression of CRC (59).
PNU-74654 is a small molecule that disrupts Wnt
pathway signaling by suppressing the interaction between β-catenin
and TCF4. Of note, Wu et al (60) found that pleomorphic adenoma gene
like-2 (PLAGL2) has a role in promoting CRC cells by upregulating
and stabilizing the level of β-catenin and promoting the
translocation of β-catenin into the nucleus, and as PNU-74654
blocks PLAGL2 activity in CRC cells, it further prevents the
development, progression and metastasis of CRC. GB1874 blocks Wnt
signaling by targeting the β-catenin-TCF4 protein-protein
interaction, thereby inhibiting xenograft tumor growth in
vivo and CRC stem cell proliferation in vitro (61).
ICAT was found to be a polypeptide that
downregulates the expression of genes that are targeted by the Wnt
pathway, such as cyclin D1 and Myc, by blocking the β-catenin-TCF
interaction, which significantly reduces the growth of CRC cells
and tumor formation (62).
Furthermore, patients with CRC and high ICAT levels have a
favorable prognosis. The study provides a scientific basis for
treatment based on ICAT and opens a new avenue for their use in
patients with CRC.
Traf2 and Nck-interacting protein kinase (TNIK) is
an important component of the β-catenin/TCF-4 transcription
complex, which is crucial for the development and progression of
CRC (63). Downregulation of TNIK
reduces the transcriptional activity of the β-catenin/TCF-4
complex, thereby inhibiting the growth of CRC and xenograft tumor
cells (64). The TNIK inhibitor
NCB-0846 reduced the expression of Wnt target genes and inhibited
the development of colorectal tumors and the stemness of CRC cells.
Furthermore, NCB-0846 also inhibited oncogenic proteins, such as
FMS-like tyrosine kinase 3, platelet-derived growth factor-α and
cyclin-dependent kinase 2/cyclin A2 (64). Of note, NCB-0846 inhibited lung
cancer metastasis by downregulating the expression of TGF-β
receptor type-I and blocking the TGF-β pathway and the SMAD
signaling pathway (65,66). Of note, it has been indicated that
in patients with CRC carrying a KRAS mutation, the expression of
the antiapoptotic protein BCL-XL is increased, and the BCL-XL
inhibitor ABT-263 synergized with NCB-0846 to induce cell death and
inhibit tumor cell proliferation (67).
LF3, a novel inhibitor of the β-catenin and TCF4
interaction, blocks the self-renewal capacity of CSCs (68). LF3 significantly inhibits tumor
cell hypermobility and progression of the cell cycle. Its
inhibitory effect on tumor cells is dose-dependent and it has a
certain therapeutic effect in Wnt/β-catenin-dependent cancers
(69).
NSAIDs may reduce the transcriptional activity of
the β-catenin-TCF/LEF complex and inhibit the classic Wnt/β-catenin
signaling pathway (70). Previous
studies have confirmed that after treatment with diclofenac,
celecoxib and sulindac, the degradation of intracellular β-catenin
was increased, β-catenin translocation to the nucleus was reduced,
and the expression of Wnt target genes, such as Axin2, cyclin D1
and c-Myc, was also reduced (71,72),
thus suppressing the proliferation of tumor cells and inducing
their apoptosis. Egashira et al (73) found that 2,5-dimethyl celecoxib, an
analog of celecoxib, reduced the expression of Wnt target genes and
further exhibited a significant antitumor effect. More importantly,
repeated use of 2,5-dimethyl celecoxib was able to significantly
reduce the size and number of tumors without any drug toxicity,
suggesting that 2,5-dimethyl celecoxib is a new potential
anticancer treatment. Of note, the combination of an NSAID
(sulindac) and rexinoid (bexarotene) resulted in a durable
amelioration of familial adenomatous polyposis, thereby preventing
the development of CRC (74).
Furthermore, a clinical trial is currently investigating the role
of aspirin as an adjunctive drug for the prevention of
postoperative CRC recurrence and metastasis (NCT02607072).
Wnt ligands bind to corresponding receptors, such as
FZD receptors and LRP5 and LRP6 coreceptors, thereby initiating
Wnt/β-catenin signaling and allowing β-catenin to accumulate in the
cytoplasm, translocate into the nucleus and activate downstream
target genes to induce tumor growth (79). OMP-18R5 (vantictumab) is a
monoclonal antibody that binds to members of the FZD family and
inhibits the cascade of tumorigenesis and progression by blocking
Wnt/β-catenin signaling (80). It
has been indicated that OMP-18R5 significantly inhibited the growth
of CRC cells and xenograft tumors, and OMP-18R5 synergized with
irinotecan (80). In other tumor
models, such as those of pancreatic cancer and gastric cancer,
OMP-18R5 inhibited the growth of tumor tissue (81,82).
It is worth noting that in a phase Ib clinical trial of OMP-18R5
for breast cancer, although there was a good response rate, the
common occurrence of fractures limited its development in the
clinic (83).
OMP-54F28 (ipafricept) is a fusion protein of FZD8
and immunoglobulins, and inhibits Wnt signaling by competing with
FZD8 receptors for ligands (84).
In addition, OMP-54F28 has been indicated to inhibit tumor cell
proliferation to a greater extent when combined with
chemotherapeutic drugs, such as gemcitabine, as compared with its
use as a monotherapy (84). A
phase I study in patients with advanced solid tumors suggested that
OMP-54F28 prolonged survival in both patients with desmoid and germ
cell carcinoma; furthermore, the patients remained stable and the
drug was well tolerated (85). In
a phase Ib clinical trial of OMP-54F28 combined with paclitaxel and
gemcitabine for the treatment of metastatic pancreatic cancer, the
rate of patients who had a clinical benefit was 81%, with 12 cases
of complete response (46.2%) and 9 cases of partial response
(34.6%). The median progression-free survival (PFS) was 5.9 months
and the median OS was 9.7 months (86). In a clinical trial of OMP-54F28
combined with paclitaxel and carboplatin for ovarian cancer, the
overall response rate was 75.7%, PFS was 10.3 months and OS was 33
months (87). The above studies
confirmed the effectiveness of OMP-54F28-mediated inhibition of
tumor growth, invasion and metastasis. In addition, Le et al
(88) reported that OMP-18R5 and
OMP-54F28 significantly inhibited the growth of xenograft tumors
and blocked Wnt signaling at the tumor epithelial-stromal boundary.
It was evident that both OMP-18R5 and OMP-54F28 inhibit tumor
growth; however, the associated risk of fractures should be noted.
However, the addition of bone protectants, such as alendronate, may
reduce bone toxicity and bone loss after Wnt ligand and receptor
inhibitor therapy (89).
The secretion of Wnt proteins requires
palmitoylation. Palmitoylation is the attachment of the 16-carbon
saturated fatty acid palmitate as a lipid donor to one or more
cysteine residues in a protein to correctly guide signal
transduction between targets and enhance the efficiency and
specificity of signal transmission (90). PPN protein, a member of the
membrane-bound O acyltransferase family that is located in the
endoplasmic reticulum, is a multiple transmembrane protein that is
required for the palmitoylation of Wnt proteins (91,92).
Therefore, targeted inhibition of the PPN protein may effectively
block excessive activation of the Wnt signaling pathway.
LGK974 is a PPN inhibitor that mitigates Wnt
palmitate transferase activity to block Wnt signaling. Bagheri
et al (93) demonstrated
that combinations of LGK974 and aspirin may lead to cell cycle
arrest of CRC cell lines and induce apoptosis. In vivo
studies have demonstrated that LGK974 inhibits tumor-cell invasion,
metastasis and xenograft tumor growth (94-97).
Of note, a recent study reported that the chemotherapeutic drug
5-fluorouracil (5-FU) increases both the translation and
transcription of the sequence-specific transcription factor p53,
which may promote Wnt3 transcription and activate the Wnt/β-catenin
signaling pathway, ultimately leading to the recurrence of CRC
(98). When combining LGK974 with
5-FU for the treatment of CRC, 5-FU-induced CSC activation was
significantly inhibited and the rate of tumor recurrence was
significantly decreased, indicating that the combination of Wnt
inhibitors with 5-FU may improve the therapeutic effects of
treatment compared with chemotherapy alone (98).
In other types of cancer, LGK974 complexed with
cyclodextrin may synergistically decrease the expression of Wnt
target genes in lung cancer organoids and transplants, reduce the
enterotoxicity of the drug and improve the safety of drug therapy
(99). In the treatment of
neuroendocrine tumors, LGK974 reduced cell viability by causing
cell cycle arrest in the G1 and G2/M phases
(100). In glioblastoma and
ovarian cancer, LGK974 exhibited synergistic effects when combined
with chemotherapeutic agents (101,102). LGK974 also inhibited the growth
of prostate cancer xenografts (103). Of note, a previous preclinical
study confirmed that LGK974 had a strong inhibitory effect on ring
finger protein (RNF)43-mutated cancer cells (104), while ~18% of patients with CRC
have RNF43 mutations (105). A
recent phase I clinical trial confirmed the safety and efficacy of
LGK974 for the treatment of advanced solid tumors; of note, the
study found that LGK974 enhances the activity of immune checkpoint
inhibitors but may affect immune-cell recruitment in tumors
(106).
IWP is a PPN inhibitor that may inhibit the
palmitoylation of Wnt3A. In addition, IWP may suppress the
phosphorylation of LRP5/6 and DVL2 and stabilize the Axin2
destruction complex to block Wnt signaling. Mo et al
(107) verified the therapeutic
effect of IWP in gastric cancer cells. The results indicated that
IWP decreased the invasion and metastasis ability of MKN28 cells.
Wnt-C59 is also a PPN protein that blocks the acylation of Wnt by
inhibiting the coprecipitation of Wnt3A and the carrier protein
Wntless, thereby blocking the activation of Wnt signaling (108,109). Studies including that by Boulter
et al (110) have
indicated that Wnt-C59 has an important role in inhibiting tumor
cell proliferation and promoting tumor cell apoptosis (110,111). Furthermore, Zhang et al
(112) confirmed that Wnt-C59 may
block EMT and suppress the growth of xenograft tumors. Of note,
Wnt-C59 also exerts anti-inflammatory effects by blocking the
interaction of β-catenin with nuclear factor κB (NF-κB) (113,114). NF-κB has a vital role in the
development, angiogenesis and metastasis of cancer and is closely
related to the development of resistance to anticancer drugs.
Hence, there is potential clinical value in performing further
clinical trials on Wnt-C59.
ETC-159 is a novel oral PNN inhibitor that may block
the secretion and activity of all Wnts. In addition,
xenotransplantation in patients with CRC with R-spondin
translocation was markedly inhibited (115). Zhong et al (116) confirmed that the combined use of
ETC-159 and the pan-PI3K inhibitor GDC-0941 markedly suppressed the
growth of RNF43-mutant xenografts in vivo by downregulating
cell proliferation and glucose metabolism. More importantly, Kaur
et al (117) found that
the combination treatment of ETC-159 with the poly(ADP-ribose)
polymerase (PARP) inhibitor olaparib resulted in the inhibition of
tumor-selective DNA repair pathways.
TNKS, a PARP that promotes the degradation of
destruction complexes through the ubiquitin-proteasome pathway, is
key to increasing the breakdown of Axin, which leads to the
activation of the Wnt signaling pathway. Of note, suppression of
TNKS increased the stability of Axin, decreased the degradation of
the destruction complex and decreased the cytoplasmic accumulation
and translocation of β-catenin into the nucleus, thereby blocking
the activation of the Wnt/β-catenin signaling pathway (118).
Both NVP-TNKS656 and NVP-XAV939 are TNKS inhibitors,
of which NVP-TNKS656 selectively inhibits TNKS2. Both are able to
stabilize Axin, lessen the accumulation of β-catenin in the
cytoplasm and block the activation of Wnt signaling (119,120). Previous studies have reported
that NVP-TNKS656 and NVP-XAV939 may inhibit the growth of
colorectal tumors to a greater extent than other drugs (120). In a study in which NVP-XAV939 was
combined with 5-FU/cisplatin (DDP)-treated SW480 and SW620 colon
cancer cells, Axin levels increased, cytoplasmic and nuclear
β-catenin levels decreased and SW480 cell apoptosis induced by
5-FU/DDP increased (121). Of
note, Yu et al (122)
reported that knockdown of caudal-related homeobox transcription
factor 2 (CDX2) in colon cancer cells promoted cell proliferation
and tumor formation in vivo, while XAV939 significantly
inhibited colon cancer cell proliferation and tumor formation after
CDX2 knockdown. In addition, XAV939 inhibited the expression of
β-catenin, cyclin D1 and c-Myc induced by overexpression of
ubiquitin-specific protease 6 N-terminal-like protein, thereby
slowing colon tumor progression (123). More importantly, XAV939 also
reversed the inhibition of Axin expression and the activation of
β-catenin caused by prohibitin 1 (PHB1) deficiency (124). PHB1 is an important regulator of
gene transcription and may block the Wnt/β-catenin signaling
pathway and the formation of intestinal tumors. In addition, XAV939
may also reduce the degree of drug resistance in
chemotherapy-resistant colon tumor cells (125,126) and overcome the resistance to 5-FU
observed in CRC cells carrying a short truncating APC mutation
(127), which is expected to be a
therapeutic strategy for chemotherapy-resistant CRC.
IWR is a TNKS inhibitor that is able to impair the
self-renewal capacity of CSCs. A study investigating the
tumor-suppressive effect of IWR-1 combined with doxorubicin
indicated that in vitro, IWR-1 and doxorubicin have
synergistic effects and reverse the drug resistance of tumor stem
cells; in vivo, IWR-1 synergistically inhibits tumor-cell
proliferation with doxorubicin and considerably slows tumor
progression (128). A study by
Cheng et al (129)
confirmed that Numb is related to EMT and its knockdown promoted
EMT through the Wnt signaling pathway, thereby accelerating the
formation of colon tumors; furthermore, IWR-1 attenuated EMT caused
by Numb knockdown to inhibit the development and progression of
colon tumors (129). Of note, it
was recently indicated that the TNKS inhibitor bis-quinazolinone
5/Cpd 5 is superior to IWR-1 based on its inhibitory activity and
stability, providing a new basis for the development of TNKS
inhibitors (130).
G007-LK and G244-LM are also TNKS inhibitors.
Studies have confirmed that G007-LK and G244-LM are able to
completely block Wnt/β-catenin signaling and inhibit the growth of
colorectal tumors caused by APC mutations. Their key mechanism is
to block the cell cycle and reduce colony formation (131,132). In addition, G007-LK is well
tolerated and inhibits Wnt signaling and the proliferation of
LGR5+ intestinal stem cells, which hinders the formation
of intestinal tumors (133).
Furthermore, in glioblastoma, G007-LK decreased the expression of
Wnt/β-catenin signaling pathway-related proteins and genes in CSCs,
suppressing cell proliferation and tumor formation (134). Of note, G007-LK may also block
Wnt/β-catenin signaling in melanoma cells and increase their
sensitivity to anti-PD-1 immunotherapy, and when used in
combination with immunotherapy, it has a synergistic effect that is
dependent on the absence of β-catenin in tumor cells (135). A study on G007-LK combined with
PI3K (BKM120) and EGFR (erlotinib) inhibitors indicated that this
combination synergistically suppressed tumor growth in patients
with CRC, implying that combinations of TNKS inhibitors with PI3K
and EGFR inhibitors may expand the treatment options for CRC
(136), which warrants additional
in-depth studies.
Numerous natural compounds have been proven to block
the Wnt/β-catenin signaling pathway by acting on different targets,
further changing the pathology of CRC and effectively preventing
and treating CRC.
Curcumin is a natural compound derived from the
rhizome of turmeric. In recent years, it has been indicated that
curcumin inhibits the proliferation of various types of cancer
cells, and its key mechanism relies on blocking the Wnt signal
transduction pathway. Previous studies have demonstrated that
curcumin promotes the stalling of p53- and p21-independent cells in
the G2/M phase and induces apoptosis in HCT-116 colon
cancer cells (138-140). In addition, the curcumin-mediated
reduction of cell proliferation and increase of apoptosis may also
be achieved by inhibiting CDX2, an intestinal-specific nuclear
transcription factor in the Wnt/β-catenin signaling pathway
(141,142). It is worth noting that a phase I
trial initially explored the efficacy and safety of curcumin in
combination with 5-FU in 5-FU-resistant metastatic colon cancer
(NCT02724202) and in combination with irinotecan for the treatment
of metastatic CRC (NCT01859858). In addition, a clinical trial
evaluating whether curcumin is beneficial for patients with rectal
cancer prior to chemotherapy and radiotherapy (NCT00745134) is in
progress, indicating that curcumin is a promising traditional
medicine for the treatment of CRC.
Genistein is a soybean-derived isoflavone and
phytoestrogen that has a key role in blocking Wnt/β-catenin
signaling by upregulating GSK-3β and E-cadherin expression
(143). A phase I/II clinical
trial (NCT01985763) indicated that genistein combined with
chemotherapy is an effective treatment for metastatic CRC, with
good safety and tolerability (144). Of note, genistein also reverses
chemoresistance (145) and is
expected to have a role in the treatment of chemoresistant CRC.
Previous epidemiological studies have demonstrated
that the amount of vitamin D in the active form is inversely
associated with the risk of CRC (146), and vitamin D deficiency increases
the risk and mortality of patients with CRC (147,148). A study by Fernández-Barral et
al (149) indicated that
1,25-(OH)2D3 reduced the proliferation of
colon CSCs and inhibited the development and progression of colon
cancer. Of note, a study evaluating the anti-colorectal tumor
activity of vitamin D-nanoemulsion (NVD) revealed that NVD induced
apoptosis in HCT-116 cells in a dose- and time-dependent manner,
and this apoptotic effect was mediated by the targeted inhibition
of β-catenin (150). Recently, a
pilot study examined the effect of vitamin D supplementation on the
survival of patients with stage II-III CRC undergoing chemotherapy
(NCT02603757). In addition, a phase I trial investigating the
vitamin D receptor in relation to CRC is ongoing (NCT02172651).
Ginsenosides are steroid compounds that are derived
from the plant ginseng. Previous studies have confirmed that
ginsenosides may inhibit the proliferation of tumor cells and
suppress their self-renewal ability by blocking Wnt/β-catenin
signaling (155,156). A study by Yuan et al
(157) found that
20(S)-ginsenoside Rh2 inhibited CRC lymph node metastasis in a
dose-dependent manner in vivo. It is worth noting that in
breast cancer models, ginsenosides may increase the sensitivity of
tumor cells to chemotherapeutic drugs, thereby improving the
efficacy of chemotherapy (158).
Tanshinone IIA is one of the main active components of the
traditional Chinese herbal medicine Salvia miltiorrhiza. It
has been confirmed that it is able to downregulate the level of
hypoxia-inducible factors-1α (HIF-1α) and block the translocation
of β-catenin into the nucleus, which subsequently blocks the
activation of TCF/LEF to decrease angiogenesis (159). This finding provides a new
therapeutic strategy for the development and progression of CRC.
Z-ajoene, a simple sulfur-containing compound extracted from
garlic, has been reported to inhibit CRC cell growth. Z-ajoene also
facilitates β-catenin degradation by regulating CK1α activity
(160). Studies using Z-ajoene
for the treatment of gastric cancer suggested that the
proliferation of MGC-803 gastric cancer cells was considerably
inhibited (161,162). Artemisinin is an active
sesquiterpene extracted from Artemisia annua. It has been
confirmed that artemisinin blocks Wnt/β-catenin signaling and
diminishes the proliferation, invasion and migration of tumor cells
(163-165).
Nerigoside (NG), derived from the Umbelliferae
celery plant, facilitates the apoptosis of a variety of tumor
cells, but its role in CRC has remained to be elucidated. Wen et
al (166) indicated that the
viability and colony formation of the CRC cell lines HT29 and SW620
were inhibited after NG treatment, and the invasive ability of the
cells was also significantly reduced. NG was also observed to
enhance the expression and stability of GSK3β to block the
Wnt/β-catenin pathway signaling (166). More importantly, the drug
toxicity associated with NG treatment is very low. Vicenin-2, an
NG, blocks Wnt/β-catenin signaling by inhibiting the transcription
of TCF/LEF (167). Ginkgolide C
is an extract derived from Ginkgo biloba leaves that blocks
the Wnt/β-catenin signaling cascade and exhibits strong anti-CRC
effects (168). Resveratrol is an
antitoxin produced by numerous plants when they are stimulated that
may inhibit CRC cell proliferation and tumor formation by blocking
the Wnt/β-catenin signaling cascade (169). A phase I trial suggested that
micronized trans-resveratrol (SRT501) treatment significantly
increased the level of the apoptosis marker caspase-3 in patients
with CRC liver metastases, and SRT501 was well tolerated (170). In addition, a phase I trial is
exploring whether resveratrol in grape skins is able to prevent
colon cancer (NCT00578396).
Andrographolide analog 3A.1 is an analog the main
active ingredient of the plant Andrographis paniculata.
Previous studies suggested that analog 3A.1 significantly reduced
the viability of human CRC HT29 cells and promoted their apoptosis
by significantly suppressing the activity of the TCF/LEF promoter
(171). DIF-3 is a small molecule
compound identified in Dictyostelium discoideum. It may
activate GSK3β and promote the degradation of β-catenin, thereby
hindering cell proliferation and promoting apoptosis (172,173). It is worth noting that the use of
DIF-3 in patients undergoing chemotherapy may significantly reduce
the severity of adverse events such as nausea and vomiting
(174). Z86 is a novel inhibitor
of the Wnt signaling pathway. Its key mechanism is to block the
phosphorylation of GSK-3β, thereby increasing the degradation of
β-catenin, downregulating the expression of endogenous Wnt
signaling target genes and ultimately suppressing CRC cell
proliferation (175).
KYA1797K is a β-catenin inhibitor that may bind
Axin and suppress tumors by promoting the instability of β-catenin
(176). It has been indicated
that high expression of β-catenin and subunit of the
oligosaccharyltransferase complex, homolog A/B (STT3A/B) in colon
cancer tissue is related to poor prognosis (177). KYA1797K may inhibit the signal
transduction of β-catenin/STT3, thereby inhibiting the
proliferation of colon CSCs and inducing apoptosis. Of note,
KYA1797K may also reduce the stability of PD-L1 on the surface of
colon tumor cells, disrupt the immune evasion of colon tumors and
contribute to the effectiveness of immunotherapies (177). In addition, KYA1797K suppresses
CSC stemness and blocks the growth of tumor organoids and
xenografts (178). BC2059
(tegatrabetan) is a novel β-catenin inhibitor that may reduce the
levels of β-catenin in the cytoplasm and nucleus and reduce the
transcriptional activity of the TCF4/LEF complex. Studies have
indicated that BC2059 may prolong the OS of patients and the side
effects are tolerable (179,180). The above studies demonstrated
that BC2059 is a potent and safe inhibitor of β-catenin and is
worthy of further study in colorectal tumor models.
ICG-001 is a small molecule β-catenin/TCF inhibitor
and its target gene myeloid ecotropic viral integration site 1
(MEIS1) promotes the proliferation and metastasis of colorectal
tumor cells. Studies have confirmed that ICG-001 significantly
inhibits the stemness and metastasis of colorectal CSCs by
inhibiting MEIS1, which reduces tumor formation and invasion
(181). Of note, ICG-001 also
reversed Wnt4-induced EMT and angiogenesis in vitro and
in vivo, significantly inhibiting colorectal tumor formation
(182,183). ICG-001 is expected to improve the
poor prognosis of patients with CRC metastases.
SR3029 is a CK1 inhibitor. Amino-terminal enhancer
of split (AES) has been indicated to suppress tumor metastasis in
CRC. Wang et al (188)
identified a CK1δ/ε-AES axis in CRC cells, in which CK1δ/ε promotes
the degradation of AES and significantly increases the
proliferation, invasion and migration of CRC cells by upregulating
the expression of Wnt target genes. After treatment with SR3029,
the growth of colorectal tumors and liver metastasis in mice was
significantly diminished.
Napabucasin (BBI608) is an orally administered
inhibitor of signal transducer and activators of transcription
(STAT)3 and cancer cell pluripotency. Abnormal expression and
activation of STAT3 has been reported in various tumor diseases and
STAT3 has been indicated to regulate β-catenin expression during
CSC self-renewal. Various studies have confirmed that BBI608 may
significantly inhibit tumor cell proliferation and colony formation
(189-192). A phase III trial of napabucasin
for refractory advanced CRC indicated no difference in OS between
the treatment and placebo groups. However, STAT3 may be an
important target for the treatment of CRC with elevated
phosphorylated STAT3 levels (193). In addition, phase II clinical
trials of BBI608 in combination with chemotherapy drugs for
patients with advanced metastatic CRC are ongoing (NCT01776307,
NCT02851004 and NCT03647839).
SM08502 is a novel small molecule under
investigation for the clinical treatment of solid tumors. SM08502
may block signaling of the Wnt pathway and downstream target gene
expression by inhibiting the activity of CDC-like kinase (194). A phase I clinical trial of
SM08502 for patients with advanced solid tumors is ongoing
(NCT03355066). ST2825 is a myeloid differentiation factor 88
(MyD88) inhibitor. MyD88 is an adapter protein that is used for
signal transduction in the IL-1β/toll-like receptor (TLR) pathway.
A preclinical study indicated that MyD88 upregulates NF-κB and
promotes Wnt/β-catenin signaling in intestinal tumor cells.
However, after treatment with ST2825, the growth of intestinal
tumor cells was inhibited and apoptosis was increased, which
significantly hindered the development of intestinal tumors
(195). FH535 is an inhibitor of
the Wnt/β-catenin pathway and peroxisome proliferator-activated
receptors with antitumor activity; thus, it has considerable
anti-proliferative and proapoptotic effects (196). In addition, Tu et al
(197) used FH535 to treat human
CRC cells (DLD-1 and SW620 cells) and found that the cell cycle was
arrested in the G2/M phase and the ability of cells to
proliferate and migrate was significantly inhibited.
Obatoclax is a B-cell CLL/Lymphoma 2 (BCL-2)
antagonist that inhibits BCL-2, BCL-xL and MCL-1 to induce
apoptosis. Obatoclax reduces the stability of β-catenin and
downregulates the transcriptional activity of the TCF/LEF complex,
significantly suppressing the growth of HCT-116 cells. More
importantly, obatoclax significantly reduced the levels of survivin
mRNA and survivin promoter activity, which induced apoptosis
(201), and revealed the
potential of obatoclax as a Wnt pathway antagonist for the
treatment of CRC.
Further possible CRC therapeutic targets have been
discovered through various general studies. For example, Gan et
al (202) found that
knockdown of protein tyrosine phosphatase receptor type F may
significantly down-regulate the expression of CSC-related genes
downstream of the Wnt/β-catenin pathway to cause an
antiproliferative effect in CRC cells. Song et al (203) found that sine oculis homeobox 1
(Six1), a homeodomain transcription factor, promotes Wnt/β-catenin
pathway signaling. After Six1 knockdown, the growth of CRC cells
was significantly decreased and cell metastasis and invasion were
significantly weakened. p38α is a novel β-catenin-associated kinase
that increases the proliferation, invasion and metastasis of
colorectal tumor cells by promoting the transcription of β-catenin
target genes. By contrast, targeted inhibition of p38α may inhibit
the activity of CSCs and increase the sensitivity of tumor cells to
chemotherapeutic drugs (204).
Clinical trials for p38α inhibition are ongoing.
Mucin 13 (MUC13), a cell surface mucin, is highly
expressed in patients with CRC and is associated with poor
prognosis. Sheng et al (205) revealed a new role for MUC13 in
the development and progression of colorectal tumors. MUC13
inhibits β-catenin degradation by interacting with GSK-3β and
promoting canonical Wnt signaling. As tumor cells are less
sensitive to immune checkpoint inhibitors (205), inhibition of MUC13 may benefit
patients with CRC. It has been reported that overexpression of RNF6
in CRC cells promotes the phosphorylation of GSK-3β, which in turn
upregulates the activation of the Wnt/β-catenin signaling pathway
and increases the formation and metastasis of colorectal tumors.
Therefore, inhibition of RNF6 is crucial in patients with CRC
(206).
Lysine-specific histone demethylase 1 (LSD1) has
been indicated be a potential target for cancer therapy. Peng et
al (207) studied the
anti-CRC activity of the LSD1 inhibitor ZY0511 when combined with
5-fluorouracil (5-HU). The results suggested that ZY0511
significantly inhibited the growth of CRC cells and induced
apoptosis both in vivo and in vitro. Furthermore,
ZY0511 synergized with 5-HU and significantly reduced the invasive
and metastatic abilities of CRC cells. Further investigation
indicated that the antitumor effect of ZY0511 was achieved by
inhibiting the Wnt/β-catenin pathway signaling.
NSC668036 is an organic inhibitor that has been
demonstrated to inhibit β-catenin-driven gene transcription in
vitro (210). Apicularen and
bafilomycin act on ATPases and inhibit the activation of the Wnt
pathway (211,212). DK419, a derivative of
niclosamide, inhibits the growth of CRC cells and xenografts in
vitro by blocking the Wnt/β-catenin signaling (213). P5091 attenuates Wnt pathway
activity by promoting β-catenin degradation, which inhibits CRC
cell proliferation and promotes apoptosis in vitro (214). 36-077, a
Phosphoinositide-3-kinase, Class 3/Vacuolar Protein Sorting 34
(PIK3C3/VPS34) kinase inhibitor, reduces 5-FU resistance in CRC
cells and induces autophagy in CRC cells by blocking Wnt/β-catenin
pathway signaling (215). Ovo
like zinc finger 2 (OVOL2) is a Wnt signaling inhibitor that
decreases the expression of related target genes by promoting the
recruitment of histone deacetylase 1 to the TCF4-β-catenin complex;
however, the level of OVOL2 is reduced in patients with CRC.
Therefore, upregulating the level of OVOL2 may be a strategy to
suppress the development and progression of CRC (216). All the above-mentioned compounds
may inhibit the signal transduction of the Wnt pathway, which
further inhibits the development and progression of colorectal
tumors.
Various drugs that are used to treat other diseases
in the clinic have also been found to inhibit Wnt/β-catenin
signaling. The safety of these drugs has been confirmed in clinical
applications, and they may be effective and safe options for the
treatment of Wnt/β-catenin signaling-dependent cancers. Niclosamide
is an antiparasitic drug that may inhibit the binding of FZD
receptors to Wnt ligands, thereby blocking Wnt/β-catenin signaling
(217). Pimozide is a neuroleptic
drug, but studies have indicated that it may also suppress the
Wnt/β-catenin signaling pathway (218,219). Ethacrynic acid is a highly potent
diuretic that was also found to inhibit the activity of the
β-catenin/LEF-1 transcription complex in liver cancer cells,
thereby reducing the expression of downstream target genes as part
of its anticancer effect (220,221). Although the above-mentioned drugs
have so far not been used for the treatment of CRC, their
inhibitory effect on the Wnt/β-catenin pathway signaling and
clinical drug safety make them favorable options for the treatment
of CRC.
The Wnt protein family has a role in cell
proliferation and differentiation, the self-renewal of stem cells
and the transmission of extracellular signals intracellularly
through cell-surface receptors, thereby driving the development and
progression of tumors. The Wnt/β-catenin pathway is the classical
Wnt signaling pathway and its dysregulation in malignant tumors has
been widely reported, particularly in digestive system tumors. For
instance, abnormal expression of β-catenin and other Wnt pathway
proteins in the cytoplasm and nucleus of CRC cells is common.
Therefore, inhibitors that target the Wnt/β-catenin signaling
pathway have good prospects for tumor therapy, as they may act on
different upstream and downstream Wnt-related factors to suppress
or impede the development and progression of CRC.
For instance, a β-catenin-TCF/LEF complex inhibitor
may downregulate the expression of proto-oncogenes, such as cyclin
D1 and c-Myc, which are downstream of Wnt. This downregulation
inhibits cell proliferation and promotes cell apoptosis, thereby
inhibiting the formation, invasion and metastasis of colorectal
tumors. Although the anti-CRC effects of numerous β-catenin-TCF/LEF
complex inhibitors have been demonstrated in preclinical studies,
their efficacy and safety in humans require to be confirmed in
clinical trials.
There are numerous types of Wnt ligands and
receptor inhibitors, and their antitumor mechanisms mainly involve
blocking the binding of Wnt ligands to their corresponding
receptors, such as FZD receptors and LRP5 and LRP6 coreceptors.
This inhibits Wnt/β-catenin signaling, which further promotes the
degradation of β-catenin in the cytoplasm, reduces β-catenin
translocation into the nucleus and downregulates the expression of
downstream target genes to inhibit the growth, invasion and
metastasis of colorectal tumors. To date, much progress has been
made in the research and development of Wnt ligands and receptor
inhibitors, and there are ongoing clinical trials to assess their
clinical benefits. Of note, Wnt ligands and receptor inhibitors may
also synergize with chemotherapeutic drugs to increase the
sensitivity of tumors to immunotherapy. These studies are expected
to provide clinicians with effective and safe treatment strategies
for patients with CRC, and an increased population of patients with
CRC that benefit from treatment.
Certain natural compounds and their derivatives
extracted from traditional Chinese medicines have been indicated to
change the pathology of CRC and effectively prevent and treat CRC
by acting on different targets in the Wnt/β-catenin signaling
pathway. In addition, studies have indicated that natural compounds
have low pharmacological toxicity and fewer adverse events in
patients. Of note, when certain natural compounds are combined with
chemotherapeutic drugs, they may synergistically exert antitumor
effects, while also limiting the adverse effects of chemotherapy.
In short, these natural compounds deserve more thorough research to
explore their anti-CRC mechanisms and to determine how combinations
may benefit patients with CRC.
There are numerous other substances that block
Wnt/β-catenin pathway signaling, and there have also been newly
identified targets within the Wnt/β-catenin signaling pathway.
Furthermore, even drugs that have been approved for the treatment
of other diseases have demonstrated significant anti-CRC
effects.
Although further advances in research have led to
the identification of numerous CRC targets in the classical Wnt
signaling pathway, which has provided additional options to treat
colorectal tumors, there are potential drawbacks to inhibiting the
Wnt pathway. Previous studies have indicated that the Wnt/β-catenin
signaling pathway is involved in numerous biological processes,
such as osteoblast differentiation, neurodevelopment and insulin
secretion; therefore, inhibition of the Wnt/β-catenin signaling
pathway may cause dysfunction in the body, leading to adverse
events such as fragility-related bone fractures, neurodegenerative
changes and metabolic dysregulation (7,222).
More importantly, research on the classical Wnt signaling pathway
still presents various challenges. For instance, the Wnt/β-catenin
signaling pathway affects CRC cells through a complex mechanism
that involves multiple receptors and protein families. In addition,
mutations in downstream targets of the Wnt/β-catenin signaling
pathway create difficulties in the research progress, as mutations
in each factor may lead to abnormal activation of this pathway. Of
note, most of the compounds targeting the Wnt/β-catenin pathway are
currently only at the preclinical stage and no new drugs have been
developed for use in patients. However, research on molecules that
target the Wnt pathway is promising and worthwhile.
The present review comprehensively summarized the
progress of research on the Wnt signaling pathway and related
active compounds with the aim of providing new targets and ideas
for the development of effective anti-CRC drugs with low toxicity.
With the progress of research, an increased number of anti-CRC
drugs that target the Wnt canonical signaling pathway will be
developed in the future, providing further therapeutic options and
individualized therapy regimens for patients with CRC. This will
allow clinicians to increase the population of patients with CRC
who benefit from treatment.
Not applicable.
YC and KD contributed to the conception and design
of the review. YC wrote the first draft of the manuscript. KD and
MC wrote sections of the manuscript. All authors contributed to
manuscript revision. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The corresponding author (KD) was supported by the National
Natural Science Foundation of China (grant no. 81600511), Science
Foundation of Sichuan Health and Family Planning Commission (grant
no. 20PJYY0314), Sichuan Science and Technology Program (grant nos.
22ZD YF1618 and 2021YFH0005), 1.3.5 Project for Disciplines of
Excellence, West China Hospital, Sichuan University (grant nos.
2019-051 and 2021-150) and Operation Funding of Sichuan
University-University of Oxford Huaxi Joint Centre for
Gastrointestinal Cancer (grant no. 161200021).
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen Y, Liu C, Zhu S, Liang X, Zhang Q,
Luo X, Yuan L and Song L: PD-1/PD-L1 immune checkpoint
blockade-based combinational treatment: Immunotherapeutic
amplification strategies against colorectal cancer. Int
Immunopharmacol. 96:1076072021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kuipers EJ, Grady WM, Lieberman D,
Seufferlein T, Sung JJ, Boelens PG, van de Velde CJ and Watanabe T:
Colorectal cancer. Nat Rev Dis Primers. 1:150652015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang Y and Wang X: Targeting the
Wnt/β-catenin signaling pathway in cancer. J Hematol Oncol.
13:1652020. View Article : Google Scholar
|
|
6
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng X, Xu X, Chen D, Zhao F and Wang W:
Therapeutic potential of targeting the Wnt/β-catenin signaling
pathway in colorectal cancer. Biomed Pharmacother. 110:473–481.
2019. View Article : Google Scholar
|
|
8
|
Krishnamurthy N and Kurzrock R: Targeting
the Wnt/betacatenin pathway in cancer: Update on effectors and
inhibitors. Cancer Treat Rev. 62:50–60. 2018. View Article : Google Scholar
|
|
9
|
Dekker E, Tanis PJ, Vleugels JLA, Kasi PM
and Wallace MB: Colorectal cancer. Lancet. 394:1467–1480. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar
|
|
11
|
Lee YH, Kung PT, Wang YH, Kuo WY, Kao SL
and Tsai WC: Effect of length of time from diagnosis to treatment
on colorectal cancer survival: A population-based study. PLoS One.
14:e02104652019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Banerjee A, Pathak S, Subramanium VD, G D,
Murugesan R and Verma RS: Strategies for targeted drug delivery in
treatment of colon cancer: Current trends and future perspectives.
Drug Discov Today. 22:1224–1232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Siegel RL, Miller KD, Goding Sauer A,
Fedewa SA, Butterly LF, Anderson JC, Cercek A, Smith RA and Jemal
A: Colorectal cancer statistics, 2020. CA Cancer J Clin.
70:145–164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Payandeh Z, Khalili S, Somi MH,
Mard-Soltani M, Baghbanzadeh A, Hajiasgharzadeh K, Samadi N and
Baradaran B: PD-1/PD-L1-dependent immune response in colorectal
cancer. J Cell Physiol. 235:5461–5475. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao Y and Freeman GJ: The microsatellite
instable subset of colorectal cancer is a particularly good
candidate for checkpoint blockade immunotherapy. Cancer Discov.
5:16–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Overman MJ, McDermott R, Leach JL, Lonardi
S, Lenz HJ, Morse MA, Desai J, Hill A, Axelson M, Moss RA, et al:
Nivolumab in patients with metastatic DNA mismatch repair-deficient
or microsatellite instability-high colorectal cancer (CheckMate
142): An open-label, multicentre, phase 2 study. Lancet Oncol.
18:1182–1191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Silva VR, Santos LS, Dias RB, Quadros CA
and Bezerra DP: Emerging agents that target signaling pathways to
eradicate colorectal cancer stem cells. Cancer Commun (Lond).
41:1275–1313. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang
J, Zhang G, Wang X, Dong Z, Chen F and Cui H: Targeting cancer stem
cell pathways for cancer therapy. Signal Transduct Target Ther.
5:82020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Voorneveld PW, Kodach LL, Jacobs RJ, van
Noesel CJ, Peppelenbosch MP, Korkmaz KS, Molendijk I, Dekker E,
Morreau H, van Pelt GW, et al: The BMP pathway either enhances or
inhibits the Wnt pathway depending on the SMAD4 and p53 status in
CRC. Br J Cancer. 112:122–130. 2015. View Article : Google Scholar :
|
|
20
|
Nusse R and Clevers H: Wnt/β-Catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Duchartre Y, Kim YM and Kahn M: The Wnt
signaling pathway in cancer. Crit Rev Oncol Hematol. 99:141–149.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Taciak B, Pruszynska I, Kiraga L, Bialasek
M and Krol M: Wnt signaling pathway in development and cancer. J
Physiol Pharmacol. 69:Jul 4–2018.Epub ahead of print. PubMed/NCBI
|
|
23
|
Wang D, Zhang Q, Li F, Wang C, Yang C and
Yu H: β-TrCP-mediated ubiquitination and degradation of Dlg5
regulates hepatocellular carcinoma cell proliferation. Cancer Cell
Int. 19:2982019. View Article : Google Scholar
|
|
24
|
DeBruine ZJ, Xu HE and Melcher K: Assembly
and architecture of the Wnt/β-catenin signalosome at the membrane.
Br J Pharmacol. 174:4564–4574. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qi J, Lee HJ, Saquet A, Cheng XN, Shao M,
Zheng JJ and Shi DL: Autoinhibition of Dishevelled protein
regulated by its extreme C terminus plays a distinct role in
Wnt/β-catenin and Wnt/planar cell polarity (PCP) signaling
pathways. J Biol Chem. 292:5898–5908. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schatoff EM, Leach BI and Dow LE: Wnt
signaling and colorectal cancer. Curr Colorectal Cancer Rep.
13:101–110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gómez-Orte E, Sáenz-Narciso B, Moreno S
and Cabello J: Multiple functions of the noncanonical Wnt pathway.
Trends Genet. 29:545–553. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De A: Wnt/Ca2+ signaling pathway: A brief
overview. Acta Biochim Biophys Sin (Shanghai). 43:745–756. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chae WJ and Bothwell ALM: Canonical and
Non-Canonical Wnt signaling in immune cells. Trends Immunol.
39:830–847. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
van Es JH, Haegebarth A, Kujala P,
Itzkovitz S, Koo BK, Boj SF, Korving J, van den Born M, van
Oudenaarden A, Robine S and Clevers H: A critical role for the Wnt
effector Tcf4 in adult intestinal homeostatic self-renewal. Mol
Cell Biol. 32:1918–1927. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sato T, Vries RG, Snippert HJ, van de
Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters
PJ and Clevers H: Single Lgr5 stem cells build crypt-villus
structures in vitro without a mesenchymal niche. Nature.
459:262–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Farin HF, Jordens I, Mosa MH, Basak O,
Korving J, Tauriello DV, de Punder K, Angers S, Peters PJ, Maurice
MM and Clevers H: Visualization of a short-range Wnt gradient in
the intestinal stem-cell niche. Nature. 530:340–343. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen M, Lin W, Li N, Wang Q, Zhu S, Zeng A
and Song L: Therapeutic approaches to colorectal cancer via
strategies based on modulation of gut microbiota. Front Microbiol.
13:9455332022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shoshkes-Carmel M, Wang YJ, Wangensteen
KJ, Tóth B, Kondo A, Massasa EE, Itzkovitz S and Kaestner KH:
Subepithelial telocytes are an important source of Wnts that
supports intestinal crypts. Nature. 557:242–246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kahn M: Wnt signaling in stem cells and
cancer stem cells: A tale of two coactivators. Prog Mol Biol Transl
Sci. 153:209–244. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aceto GM, Catalano T and Curia MC:
Molecular aspects of colorectal adenomas: The interplay among
microenvironment, oxidative stress, and predisposition. Biomed Res
Int. 2020:17263092020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bright-Thomas RM and Hargest R: APC,
beta-Catenin and hTCF-4; an unholy trinity in the genesis of
colorectal cancer. Eur J Surg Oncol. 29:107–117. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang L and Shay JW: Multiple roles of APC
and its therapeutic implications in colorectal cancer. J Natl
Cancer Inst. 109:djw3322017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu N, Shen C, Luo Y, Xia L, Xue F, Xia Q
and Zhang J: Upregulated miR-130a increases drug resistance by
regulating RUNX3 and Wnt signaling in cisplatin-treated HCC cell.
Biochem Biophys Res Commun. 425:468–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shen DY, Zhang W, Zeng X and Liu CQ:
Inhibition of Wnt/β-catenin signaling downregulates P-glycoprotein
and reverses multi-drug resistance of cholangiocarcinoma. Cancer
Sci. 104:1303–1308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Krausova M and Korinek V: Wnt signaling in
adult intestinal stem cells and cancer. Cell Signal. 26:570–579.
2014. View Article : Google Scholar
|
|
42
|
Roy S and Majumdar AP: Signaling in colon
cancer stem cells. J Mol Signal. 7:112012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sebio A, Kahn M and Lenz HJ: The potential
of targeting Wnt/β-catenin in colon cancer. Expert Opin Ther
Targets. 18:611–615. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tai D, Wells K, Arcaroli J, Vanderbilt C,
Aisner DL, Messersmith WA and Lieu CH: Targeting the WNT signaling
pathway in cancer therapeutics. Oncologist. 20:1189–1198. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Paluszczak J, Kleszcz R, Studzińska-Sroka
E and Krajka-Kuźniak V: Lichen-derived caperatic acid and physodic
acid inhibit Wnt signaling in colorectal cancer cells. Mol Cell
Biochem. 441:109–124. 2018. View Article : Google Scholar :
|
|
46
|
Gekas C, D'Altri T, Aligué R, González J,
Espinosa L and Bigas A: β-Catenin is required for T-cell leukemia
initiation and MYC transcription downstream of Notch1. Leukemia.
30:2002–2010. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li L, Peng W, Zhou Q, Wan JP, Wang XT and
Qi HB: LRP6 regulates Rab7-mediated autophagy through the
Wnt/β-catenin pathway to modulate trophoblast cell migration and
invasion. J Cell Biochem. 121:1599–1609. 2020. View Article : Google Scholar
|
|
48
|
Matsuzaki S and Darcha C: Involvement of
the Wnt/β-catenin signaling pathway in the cellular and molecular
mechanisms of fibrosis in endometriosis. PLoS One. 8:e768082013.
View Article : Google Scholar
|
|
49
|
Kim JY, Park G, Krishnan M, Ha E and Chun
KS: Selective Wnt/β-catenin Small-molecule Inhibitor CWP232228
impairs tumor growth of colon cancer. Anticancer Res. 39:3661–3667.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jang GB, Hong IS, Kim RJ, Lee SY, Park SJ,
Lee ES, Park JH, Yun CH, Chung JU, Lee KJ, et al: Wnt/β-Catenin
small-molecule inhibitor CWP232228 preferentially inhibits the
growth of breast cancer stem-like cells. Cancer Res. 75:1691–1702.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim JY, Lee HY, Park KK, Choi YK, Nam JS
and Hong IS: CWP232228 targets liver cancer stem cells through
Wnt/β-catenin signaling: A novel therapeutic approach for liver
cancer treatment. Oncotarget. 7:20395–20409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kazi A, Xiang S, Yang H, Delitto D,
Trevino J, Jiang RHY, Ayaz M, Lawrence HR, Kennedy P and Sebti SM:
GSK3 suppression upregulates β-catenin and c-Myc to abrogate
KRas-dependent tumors. Nat Commun. 9:51542018. View Article : Google Scholar
|
|
53
|
Wong CC, Xu J, Bian X, Wu JL, Kang W, Qian
Y, Li W, Chen H, Gou H, Liu D, et al: In colorectal cancer cells
with mutant KRAS, SLC25A22-Mediated Glutaminolysis Reduces DNA
demethylation to increase WNT signaling, stemness, and drug
resistance. Gastroenterology. 159:2163–2180.e6. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Du F, Cao T, Xie H, Li T, Sun L, Liu H,
Guo H, Wang X, Liu Q, Kim T, et al: KRAS Mutation-Responsive
miR-139-5p inhibits colorectal cancer progression and is repressed
by Wnt signaling. Theranostics. 10:7335–7350. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mologni L, Brussolo S, Ceccon M and
Gambacorti-Passerini C: Synergistic effects of combined Wnt/KRAS
inhibition in colorectal cancer cells. PLoS One. 7:e514492012.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chen Z, Venkatesan AM, Dehnhardt CM, Dos
Santos O, Delos Santos E, Ayral-Kaloustian S, Chen L, Geng Y, Arndt
KT, Lucas J, et al: 2,4-Diamino-quinazolines as inhibitors of
beta-catenin/Tcf-4 pathway: Potential treatment for colorectal
cancer. Bioorg Med Chem Lett. 19:4980–4983. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dehnhardt CM, Venkatesan AM, Chen Z,
Ayral-Kaloustian S, Dos Santos O, Delos Santos E, Curran K,
Follettie MT, Diesl V, Lucas J, et al: Design and synthesis of
novel diaminoquinazolines with in vivo efficacy for
beta-catenin/T-cell transcriptional factor 4 pathway inhibition. J
Med Chem. 53:897–910. 2010. View Article : Google Scholar
|
|
58
|
Chang TS, Lu CK, Hsieh YY, Wei KL, Chen
WM, Tung SY, Wu CS, Chan MWY and Chiang MK:
2,4-Diamino-Quinazoline, a Wnt signaling inhibitor, suppresses
gastric cancer progression and metastasis. Int J Mol Sci.
21:59012020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fan HC, Hsieh YC, Li LH and Chang CC,
Janoušková K, Ramani MV, Subbaraju GV, Cheng KT and Chang CC:
Dehydroxyhispolon methyl ether, a hispolon derivative, inhibits
WNT/β-Catenin signaling to elicit human colorectal carcinoma cell
apoptosis. Int J Mol Sci. 21:88392020. View Article : Google Scholar
|
|
60
|
Wu L, Zhou Z, Han S, Chen J, Liu Z, Zhang
X, Yuan W, Ji J and Shu X: PLAGL2 promotes epithelial-mesenchymal
transition and mediates colorectal cancer metastasis via
β-catenin-dependent regulation of ZEB1. Br J Cancer. 122:578–589.
2020. View Article : Google Scholar
|
|
61
|
Low JL, Du W, Gocha T, Oguz G, Zhang X,
Chen MW, Masirevic S, Yim DGR, Tan IBH, Ramasamy A, et al:
Molecular docking-aided identification of small molecule inhibitors
targeting β-catenin-TCF4 interaction. iScience. 24:1025442021.
View Article : Google Scholar
|
|
62
|
Hu J, Wang Z, Chen J, Yu Z, Zhang J, Li W,
Lin M, Yang X and Liu H: Overexpression of ICAT inhibits the
progression of colorectal cancer by binding with β-Catenin in the
cytoplasm. Technol Cancer Res Treat. 20:153303382110412532021.
View Article : Google Scholar
|
|
63
|
Masuda M, Uno Y, Ohbayashi N, Ohata H,
Mimata A, Kukimoto-Niino M, Moriyama H and Kashimoto S: TNIK
inhibition abrogates colorectal cancer stemness. Nat Commun.
7:125862016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yamada T and Masuda M: Emergence of TNIK
inhibitors in cancer therapeutics. Cancer Sci. 108:818–823. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sugano T, Masuda M, Takeshita F, Motoi N,
Hirozane T, Goto N, Kashimoto S, Uno Y, Moriyama H, Sawa M, et al:
Pharmacological blockage of transforming growth factor-β signalling
by a Traf2- and Nck-interacting kinase inhibitor, NCB-0846. Br J
Cancer. 124:228–236. 2021. View Article : Google Scholar
|
|
66
|
Sekita T, Yamada T, Kobayashi E, Yoshida
A, Hirozane T, Kawai A, Uno Y, Moriyama H, Sawa M, Nagakawa Y, et
al: Feasibility of targeting Traf2-and-Nck-Interacting kinase in
synovial sarcoma. Cancers (Basel). 12:12582020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Jung HR, Oh Y, Na D, Min S, Kang J, Jang
D, Shin S, Kim J, Lee SE, Jeong EM, et al: CRISPR screens identify
a novel combination treatment targeting BCL-XL and WNT
signaling for KRAS/BRAF-mutated colorectal cancers. Oncogene.
40:3287–3302. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhao L, Sun L, Lu Y, Li F and Xu H: A
small-molecule LF3 abrogates β-catenin/TCF4-mediated suppression of
NaV 1.5 expression in HL-1 cardiomyocytes. J Mol Cell
Cardiol. 135:90–96. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Fang L, Zhu Q, Neuenschwander M, Specker
E, Wulf-Goldenberg A, Weis WI, von Kries JP and Birchmeier W: A
small-molecule antagonist of the β-Catenin/TCF4 interaction blocks
the self-renewal of cancer stem cells and suppresses tumorigenesis.
Cancer Res. 76:891–901. 2016. View Article : Google Scholar
|
|
70
|
Gurpinar E, Grizzle WE and Piazza GA:
NSAIDs inhibit tumorigenesis, but how? Clin Cancer Res.
20:1104–1113. 2014. View Article : Google Scholar :
|
|
71
|
Sareddy GR, Kesanakurti D, Kirti PB and
Babu PP: Nonsteroidal anti-inflammatory drugs diclofenac and
celecoxib attenuates Wnt/β-catenin/Tcf signaling pathway in human
glioblastoma cells. Neurochem Res. 38:2313–2322. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Li N, Xi Y, Tinsley HN, Gurpinar E, Gary
BD, Zhu B, Li Y, Chen X, Keeton AB, Abadi AH, et al: Sulindac
selectively inhibits colon tumor cell growth by activating the
cGMP/PKG pathway to suppress Wnt/β-catenin signaling. Mol Cancer
Ther. 12:1848–1859. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Egashira I, Takahashi-Yanaga F, Nishida R,
Arioka M, Igawa K, Tomooka K, Nakatsu Y, Tsuzuki T, Nakabeppu Y,
Kitazono T and Sasaguri T: Celecoxib and 2,5-dimethylcelecoxib
inhibit intestinal cancer growth by suppressing the Wnt/β-catenin
signaling pathway. Cancer Sci. 108:108–115. 2017. View Article : Google Scholar
|
|
74
|
Bowen CM, Walter L, Borras E, Wu W, Ozcan
Z, Chang K, Bommi PV, Taggart MW, Thirumurthi S, Lynch PM, et al:
Combination of sulindac and bexarotene for prevention of intestinal
carcinogenesis in familial adenomatous polyposis. Cancer Prev Res
(Phila). 14:851–862. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wu M, Guan J, Li C, Gunter S, Nusrat L, Ng
S, Dhand K, Morshead C, Kim A and Das S: Aberrantly activated Cox-2
and Wnt signaling interact to maintain cancer stem cells in
glioblastoma. Oncotarget. 8:82217–82230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Bindu S, Mazumder S and Bandyopadhyay U:
Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A
current perspective. Biochem Pharmacol. 180:1141472020. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Grosser T, Ricciotti E and FitzGerald GA:
The cardiovascular pharmacology of nonsteroidal anti-inflammatory
drugs. Trends Pharmacol Sci. 38:733–748. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Walker C and Biasucci LM: Cardiovascular
safety of non-steroidal anti-inflammatory drugs revisited. Postgrad
Med. 130:55–71. 2018. View Article : Google Scholar
|
|
79
|
Yan KS, Janda CY, Chang J, Zheng GXY,
Larkin KA, Luca VC, Chia LA, Mah AT, Han A, Terry JM, et al:
Non-equivalence of Wnt and R-spondin ligands during
Lgr5+ intestinal stem-cell self-renewal. Nature.
545:238–242. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gurney A, Axelrod F, Bond CJ, Cain J,
Chartier C, Donigan L, Fischer M, Chaudhari A, Ji M, Kapoun AM, et
al: Wnt pathway inhibition via the targeting of Frizzled receptors
results in decreased growth and tumorigenicity of human tumors.
Proc Natl Acad Sci USA. 109:11717–11722. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang Y, Morris JP IV, Yan W, Schofield
HK, Gurney A, Simeone DM, Millar SE, Hoey T, Hebrok M and Pasca di
Magliano M: Canonical wnt signaling is required for pancreatic
carcinogenesis. Cancer Res. 73:4909–4922. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Flanagan DJ, Barker N, Costanzo NSD, Mason
EA, Gurney A, Meniel VS, Koushyar S, Austin CR, Ernst M, Pearson
HB, et al: Frizzled-7 is required for Wnt signaling in gastric
tumors with and without apc mutations. Cancer Res. 79:970–981.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Diamond JR, Becerra C, Richards D, Mita A,
Osborne C, O'Shaughnessy J, Zhang C, Henner R, Kapoun AM, Xu L, et
al: Phase Ib clinical trial of the anti-frizzled antibody
vantictumab (OMP-18R5) plus paclitaxel in patients with locally
advanced or metastatic HER2-negative breast cancer. Breast Cancer
Res Treat. 184:53–62. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Le PN, McDermott JD and Jimeno A:
Targeting the Wnt pathway in human cancers: Therapeutic targeting
with a focus on OMP-54F28. Pharmacol Ther. 146:1–11. 2015.
View Article : Google Scholar
|
|
85
|
Jimeno A, Gordon M, Chugh R, Messersmith
W, Mendelson D, Dupont J, Stagg R, Kapoun AM, Xu L, Uttamsingh S,
et al: A First-in-human phase I study of the anticancer stem cell
agent ipafricept (OMP-54F28), a decoy receptor for Wnt ligands, in
patients with advanced solid tumors. Clin Cancer Res. 23:7490–7497.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Dotan E, Cardin DB, Lenz HJ, Messersmith
W, O'Neil B, Cohen SJ, Denlinger CS, Shahda S, Astsaturov I, Kapoun
AM, et al: Phase Ib study of wnt inhibitor ipafricept with
gemcitabine and nab-paclitaxel in patients with previously
untreated stage IV pancreatic cancer. Clin Cancer Res.
26:5348–5357. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Moore KN, Gunderson CC, Sabbatini P,
McMeekin DS, Mantia-Smaldone G, Burger RA, Morgan MA, Kapoun AM,
Brachmann RK, Stagg R, et al: A phase 1b dose escalation study of
ipafricept (OMP54F28) in combination with paclitaxel and
carboplatin in patients with recurrent platinum-sensitive ovarian
cancer. Gynecol Oncol. 154:294–301. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Le PN, Keysar SB, Miller B, Eagles JR,
Chimed TS, Reisinger J, Gomez KE, Nieto C, Jackson BC, Somerset HL,
et al: Wnt signaling dynamics in head and neck squamous cell cancer
tumor-stroma interactions. Mol Carcinog. 58:398–410. 2019.
View Article : Google Scholar :
|
|
89
|
Madan B, McDonald MJ, Foxa GE, Diegel CR,
Williams BO and Virshup DM: Bone loss from Wnt inhibition mitigated
by concurrent alendronate therapy. Bone Res. 6:172018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Resh MD: Palmitoylation of ligands,
receptors, and intracellular signaling molecules. Sci STKE.
2006:re142006. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Torres VI, Godoy JA and Inestrosa NC:
Modulating Wnt signaling at the root: Porcupine and Wnt acylation.
Pharmacol Ther. 198:34–45. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Nile AH and Hannoush RN: Fatty acylation
of Wnt proteins. Nat Chem Biol. 12:60–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Bagheri M, Tabatabae Far MA, Mirzaei H and
Ghasemi F: Evaluation of antitumor effects of aspirin and LGK974
drugs on cellular signaling pathways, cell cycle and apoptosis in
colorectal cancer cell lines compared to oxaliplatin drug. Fundam
Clin Pharmacol. 34:51–64. 2020. View Article : Google Scholar
|
|
94
|
Hayashi M, Baker A, Goldstein SD, Albert
CM, Jackson KW, McCarty G, Kahlert UD and Loeb DM: Inhibition of
porcupine prolongs metastasis free survival in a mouse xenograft
model of Ewing sarcoma. Oncotarget. 8:78265–78276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Rudy SF, Brenner JC, Harris JL, Liu J, Che
J, Scott MV, Owen JH, Komarck CM, Graham MP, Bellile EL, et al: In
vivo Wnt pathway inhibition of human squamous cell carcinoma growth
and metastasis in the chick chorioallantoic model. J Otolaryngol
Head Neck Surg. 45:262016. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Liu J, Pan S, Hsieh MH, Ng N, Sun F, Wang
T, Kasibhatla S, Schuller AG, Li AG, Cheng D, et al: Targeting
Wnt-driven cancer through the inhibition of Porcupine by LGK974.
Proc Natl Acad Sci USA. 110:20224–20229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li J, Wu G, Xu Y, Li J, Ruan N, Chen Y,
Zhang Q and Xia Q: Porcupine Inhibitor LGK974 Downregulates the Wnt
signaling pathway and inhibits clear cell renal cell carcinoma.
Biomed Res Int. 2020:25276432020.PubMed/NCBI
|
|
98
|
Cho YH, Ro EJ, Yoon JS, Mizutani T, Kang
DW, Park JC, Il Kim T, Clevers H and Choi KY: 5-FU promotes
stemness of colorectal cancer via p53-mediated WNT/β-catenin
pathway activation. Nat Commun. 11:53212020. View Article : Google Scholar
|
|
99
|
Guimaraes PPG, Tan M, Tammela T, Wu K,
Chung A, Oberli M, Wang K, Spektor R, Riley RS, Viana CTR, et al:
Potent in vivo lung cancer Wnt signaling inhibition via
cyclodextrin-LGK974 inclusion complexes. J Control Release.
290:75–87. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Jin XF, Spoettl G, Maurer J, Nölting S and
Auernhammer CJ: Inhibition of Wnt/β-Catenin signaling in
neuroendocrine tumors in vitro: Antitumoral effects. Cancers
(Basel). 12:3452020. View Article : Google Scholar
|
|
101
|
Suwala AK, Koch K, Rios DH, Aretz P,
Uhlmann C, Ogorek I, Felsberg J, Reifenberger G, Köhrer K, Deenen
R, et al: Inhibition of Wnt/beta-catenin signaling downregulates
expression of aldehyde dehydrogenase isoform 3A1 (ALDH3A1) to
reduce resistance against temozolomide in glioblastoma in vitro.
Oncotarget. 9:22703–22716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Boone JD, Arend RC, Johnston BE, Cooper
SJ, Gilchrist SA, Oelschlager DK, Grizzle WE, McGwin G Jr, Gangrade
A, Straughn JM Jr and Buchsbaum DJ: Targeting the Wnt/β-catenin
pathway in primary ovarian cancer with the porcupine inhibitor
WNT974. Lab Invest. 96:249–259. 2016. View Article : Google Scholar
|
|
103
|
Bland T, Wang J, Yin L, Pu T, Li J, Gao J,
Lin TP, Gao AC and Wu BJ: WLS-Wnt signaling promotes neuroendocrine
prostate cancer. iScience. 24:1019702021. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Jiang X, Hao HX, Growney JD, Woolfenden S,
Bottiglio C, Ng N, Lu B, Hsieh MH, Bagdasarian L, Meyer R, et al:
Inactivating mutations of RNF43 confer Wnt dependency in pancreatic
ductal adenocarcinoma. Proc Natl Acad Sci USA. 110:12649–12654.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Li Y, Li J, Wang R, Zhang L, Fu G, Wang X,
Wang Y, Fang C, Zhang D, Du D, et al: Frequent RNF43 mutation
contributes to moderate activation of Wnt signaling in colorectal
signet-ring cell carcinoma. Protein Cell. 11:292–298. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Rodon J, Argilés G, Connolly RM,
Vaishampayan U, de Jonge M, Garralda E, Giannakis M, Smith DC,
Dobson JR, McLaughlin ME, et al: Phase 1 study of single-agent
WNT974, a first-in-class Porcupine inhibitor, in patients with
advanced solid tumours. Br J Cancer. 125:28–37. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Mo ML, Li MR, Chen Z, Liu XW, Sheng Q and
Zhou HM: Inhibition of the Wnt palmitoyltransferase porcupine
suppresses cell growth and downregulates the Wnt/β-catenin pathway
in gastric cancer. Oncol Lett. 5:1719–1723. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Blyszczuk P, Müller-Edenborn B, Valenta T,
Osto E, Stellato M, Behnke S, Glatz K, Basler K, Lüscher TF,
Distler O, et al: Transforming growth factor-β-dependent Wnt
secretion controls myofibroblast formation and myocardial fibrosis
progression in experimental autoimmune myocarditis. Eur Heart J.
38:1413–1425. 2017.
|
|
109
|
Najdi R, Proffitt K, Sprowl S, Kaur S, Yu
J, Covey TM, Virshup DM and Waterman ML: A uniform human Wnt
expression library reveals a shared secretory pathway and unique
signaling activities. Differentiation. 84:203–213. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Boulter L, Guest RV, Kendall TJ, Wilson
DH, Wojtacha D, Robson AJ, Ridgway RA, Samuel K, Van Rooijen N,
Barry ST, et al: WNT signaling drives cholangiocarcinoma growth and
can be pharmacologically inhibited. J Clin Invest. 125:1269–1285.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Katoh M: Multi-layered prevention and
treatment of chronic inflammation, organ fibrosis and cancer
associated with canonical WNT/β-catenin signaling activation
(Review). Int J Mol Med. 42:713–725. 2018.PubMed/NCBI
|
|
112
|
Zhang J, Cai H, Sun L, Zhan P, Chen M,
Zhang F, Ran Y and Wan J: LGR5, a novel functional glioma stem cell
marker, promotes EMT by activating the Wnt/β-catenin pathway and
predicts poor survival of glioma patients. J Exp Clin Cancer Res.
37:2252018. View Article : Google Scholar
|
|
113
|
Jang J, Song J, Sim I, Kwon YV and Yoon Y:
Wnt-Signaling Inhibitor Wnt-C59 suppresses the cytokine
upregulation in multiple organs of lipopolysaccharide-induced
endotoxemic mice via reducing the interaction between β-Catenin and
NF-κB. Int J Mol Sci. 22:62492021. View Article : Google Scholar
|
|
114
|
Jang J, Song J, Sim I and Yoon Y: Wnt-C59
inhibits proinflammatory cytokine expression by reducing the
interaction between β-catenin and NF-κB in LPS-stimulated
epithelial and macrophage cells. Korean J Physiol Pharmacol.
25:307–319. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Madan B, Ke Z, Harmston N, Ho SY, Frois
AO, Alam J, Jeyaraj DA, Pendharkar V, Ghosh K, Virshup IH, et al:
Wnt addiction of genetically defined cancers reversed by PORCN
inhibition. Oncogene. 35:2197–2207. 2016. View Article : Google Scholar
|
|
116
|
Zhong Z, Sepramaniam S, Chew XH, Wood K,
Lee MA, Madan B and Virshup DM: PORCN inhibition synergizes with
PI3K/mTOR inhibition in Wnt-addicted cancers. Oncogene.
38:6662–6677. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kaur A, Lim JYS, Sepramaniam S, Patnaik S,
Harmston N, Lee MA, Petretto E, Virshup DM and Madan B: WNT
inhibition creates a BRCA-like state in Wnt-addicted cancer. EMBO
Mol Med. 13:e133492021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Shirai F, Mizutani A, Yashiroda Y, Tsumura
T, Kano Y, Muramatsu Y, Chikada T, Yuki H, Niwa H, Sato S, et al:
Design and discovery of an orally efficacious spiroindolinone-based
tankyrase inhibitor for the treatment of colon cancer. J Med Chem.
63:4183–4204. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Kulak O, Chen H, Holohan B, Wu X, He H,
Borek D, Otwinowski Z, Yamaguchi K, Garofalo LA, Ma Z, et al:
Disruption of Wnt/β-Catenin signaling and telomeric shortening are
inextricable consequences of tankyrase inhibition in human cells.
Mol Cell Biol. 35:2425–2435. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Arqués O, Chicote I, Puig I, Tenbaum SP,
Argilés G, Dienstmann R, Fernández N, Caratù G, Matito J,
Silberschmidt D, et al: Tankyrase Inhibition Blocks Wnt/β-Catenin
pathway and reverts resistance to PI3K and AKT inhibitors in the
treatment of colorectal cancer. Clin Cancer Res. 22:644–656. 2016.
View Article : Google Scholar
|
|
121
|
Wu X, Luo F, Li J, Zhong X and Liu K:
Tankyrase1 inhibitior XAV939 increases chemosensitivity in colon
cancer cell lines via inhibition of the Wnt signaling pathway. Int
J Oncol. 48:1333–1340. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Yu J, Liu D, Sun X, Yang K, Yao J, Cheng
C, Wang C and Zheng J: CDX2 inhibits the proliferation and tumor
formation of colon cancer cells by suppressing Wnt/β-catenin
signaling via transactivation of GSK-3β and Axin2 expression. Cell
Death Dis. 10:262019. View Article : Google Scholar
|
|
123
|
Sun K, He SB, Yao YZ, Qu JG, Xie R, Ma YQ,
Zong MH and Chen JX: Tre2 (USP6NL) promotes colorectal cancer cell
proliferation via Wnt/β-catenin pathway. Cancer Cell Int.
19:1022019. View Article : Google Scholar
|
|
124
|
Alula KM, Delgado-Deida Y, Jackson DN,
Venuprasad K and Theiss AL: Nuclear partitioning of Prohibitin 1
inhibits Wnt/β-catenin-dependent intestinal tumorigenesis.
Oncogene. 40:369–383. 2021. View Article : Google Scholar
|
|
125
|
Wang T, Ning K, Lu TX and Hua D: Elevated
expression of TrpC5 and GLUT1 is associated with chemoresistance in
colorectal cancer. Oncol Rep. 37:1059–1065. 2017. View Article : Google Scholar
|
|
126
|
Xu J, Lv G, Xu B and Jiang B:
Overexpression of UBE2M through Wnt/β-Catenin signaling is
associated with poor prognosis and chemotherapy resistance in
colorectal cancer. Transl Cancer Res. 9:5614–5625. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Siraj AK, Kumar Parvathareddy S,
Pratheeshkumar P, Padmaja Divya S, Ahmed SO, Melosantos R, Begum R,
Concepcion RMJA, Al-Sanea N, Ashari LH, et al: APC truncating
mutations in Middle Eastern Population: Tankyrase inhibitor is an
effective strategy to sensitize APC mutant CRC To 5-FU
chemotherapy. Biomed Pharmacother. 121:1095722020. View Article : Google Scholar
|
|
128
|
Martins-Neves SR, Paiva-Oliveira DI,
Fontes-Ribeiro C, Bovée JVMG, Cleton-Jansen AM and Gomes CMF:
IWR-1, a tankyrase inhibitor, attenuates Wnt/β-catenin signaling in
cancer stem-like cells and inhibits in vivo the growth of a
subcutaneous human osteosarcoma xenograft. Cancer Lett. 414:1–15.
2018. View Article : Google Scholar
|
|
129
|
Cheng C, Huang Z, Zhou R, An H, Cao G, Ye
J, Huang C and Wu D: Numb negatively regulates the
epithelial-to-mesenchymal transition in colorectal cancer through
the Wnt signaling pathway. Am J Physiol Gastrointest Liver Physiol.
318:G841–G853. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Okunlola FO, Akawa OB, Subair TI, Omolabi
KF and Soliman MES: Unravelling the mechanistic role of
quinazolinone pharmacophore in the inhibitory activity of
bis-quinazolinone derivative on tankyrase-1 in the treatment of
colorectal cancer (CRC) and non-small cell lung cancer (NSCLC): A
computational approach. Cell Biochem Biophys. 80:1–10. 2022.
View Article : Google Scholar
|
|
131
|
Lau T, Chan E, Callow M, Waaler J, Boggs
J, Blake RA, Magnuson S, Sambrone A, Schutten M, Firestein R, et
al: A novel tankyrase small-molecule inhibitor suppresses APC
mutation-driven colorectal tumor growth. Cancer Res. 73:3132–3144.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Katoh M: Canonical and non-canonical WNT
signaling in cancer stem cells and their niches: Cellular
heterogeneity, omics reprogramming, targeted therapy and tumor
plasticity (Review). Int J Oncol. 51:1357–1369. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Norum JH, Skarpen E, Brech A, Kuiper R,
Waaler J, Krauss S and Sørlie T: The tankyrase inhibitor G007-LK
inhibits small intestine LGR5+ stem cell proliferation
without altering tissue morphology. Biol Res. 51:32018. View Article : Google Scholar
|
|
134
|
Kierulf-Vieira KS, Sandberg CJ, Waaler J,
Lund K, Skaga E, Saberniak BM, Panagopoulos I, Brandal P, Krauss S,
Langmoen IA and Vik-Mo EO: A small-molecule tankyrase inhibitor
reduces glioma stem cell proliferation and sphere formation.
Cancers (Basel). 12:16302020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Waaler J, Mygland L, Tveita A, Strand MF,
Solberg NT, Olsen PA, Aizenshtadt A, Fauskanger M, Lund K, Brinch
SA, et al: Tankyrase inhibition sensitizes melanoma to PD-1 immune
checkpoint blockade in syngeneic mouse models. Commun Biol.
3:1962020. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Solberg NT, Waaler J, Lund K, Mygland L,
Olsen PA and Krauss S: TANKYRASE inhibition enhances the
antiproliferative effect of PI3K and EGFR inhibition, mutually
affecting β-CATENIN and AKT signaling in colorectal cancer. Mol
Cancer Res. 16:543–553. 2018. View Article : Google Scholar
|
|
137
|
Tang L, Zhu H, Yang X, Xie F, Peng J,
Jiang D, Xie J, Qi M and Yu L: Shizukaol D, a dimeric sesquiterpene
isolated from chloranthus serratus, represses the growth of human
liver cancer cells by modulating wnt signalling pathway. PLoS One.
11:e01520122016. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Pricci M, Girardi B, Giorgio F, Losurdo G,
Ierardi E and Di Leo A: Curcumin and colorectal cancer: From basic
to clinical evidences. Int J Mol Sci. 21:23642020. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Weng W and Goel A: Curcumin and colorectal
cancer: An update and current perspective on this natural medicine.
Semin Cancer Biol. 80:73–86. 2022. View Article : Google Scholar
|
|
140
|
Villegas C, Perez R, Sterner O,
González-Chavarría I and Paz C: Curcuma as an adjuvant in
colorectal cancer treatment. Life Sci. 286:1200432021. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Jiang X, Li S, Qiu X, Cong J, Zhou J and
Miu W: Curcumin inhibits cell viability and increases apoptosis of
SW620 human colon adenocarcinoma cells via the caudal type
homeobox-2 (CDX2)/Wnt/β-catenin pathway. Med Sci Monit.
25:7451–7458. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Bian Y, Wang G, Zhou J, Yin G, Liu T,
Liang L, Liang L, Yang X, Zhang W, Ni K, et al: Astragalus
membranaceus (Huangqi) and Rhizoma curcumae (Ezhu) decoction
suppresses colorectal cancer via downregulation of Wnt5/β-Catenin
signal. Chin Med. 17:112022. View Article : Google Scholar
|
|
143
|
Wu X, Yu N, Zhang Y, Ye Y, Sun W, Ye L, Wu
H, Yang Z, Wu L and Wang F: Radix Tetrastigma hemsleyani flavone
exhibits antitumor activity in colorectal cancer via Wnt/β-catenin
signaling pathway. Onco Targets Ther. 11:6437–6446. 2018.
View Article : Google Scholar :
|
|
144
|
Pintova S, Dharmupari S, Moshier E,
Zubizarreta N, Ang C and Holcombe RF: Genistein combined with
FOLFOX or FOLFOX-Bevacizumab for the treatment of metastatic
colorectal cancer: Phase I/II pilot study. Cancer Chemother
Pharmacol. 84:591–598. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Křížová L, Dadáková K, Kašparovská J and
Kašparovský T: Isoflavones. Molecules. 24:10762019. View Article : Google Scholar :
|
|
146
|
Dou R, Ng K, Giovannucci EL, Manson JE,
Qian ZR and Ogino S: Vitamin D and colorectal cancer: Molecular,
epidemiological and clinical evidence. Br J Nutr. 115:1643–1660.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Wesselink E, Kok DE, Bours MJL, de Wilt
JHW, van Baar H, van Zutphen M, Geijsen AMJR, Keulen ETP, Hansson
BME, van den Ouweland J, et al: Vitamin D, magnesium, calcium, and
their interaction in relation to colorectal cancer recurrence and
all-cause mortality. Am J Clin Nutr. 111:1007–1017. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Kim H, Lipsyc-Sharf M, Zong X, Wang X, Hur
J, Song M, Wang M, Smith-Warner SA, Fuchs C, Ogino S, et al: Total
Vitamin D intake and risks of early-onset colorectal cancer and
precursors. Gastroenterology 2021. 161:1208–1217.e9. 2021.
|
|
149
|
Fernández-Barral A, Costales-Carrera A,
Buira SP, Jung P, Ferrer-Mayorga G, Larriba MJ, Bustamante-Madrid
P, Domínguez O, Real FX, Guerra-Pastriá L, et al: Vitamin D
differentially regulates colon stem cells in patient-derived normal
and tumor organoids. FEBS J. 287:53–72. 2020. View Article : Google Scholar
|
|
150
|
Razak S, Afsar T, Almajwal A, Alam I and
Jahan S: Growth inhibition and apoptosis in colorectal cancer cells
induced by Vitamin D-Nanoemulsion (NVD): Involvement of
Wnt/β-catenin and other signal transduction pathways. Cell Biosci.
9:152019. View Article : Google Scholar
|
|
151
|
Ren H, Zhao J, Fan D, Wang Z, Zhao T, Li
Y, Zhao Y, Adelson D and Hao H: Alkaloids from nux vomica
suppresses colon cancer cell growth through Wnt/β-catenin signaling
pathway. Phytother Res. 33:1570–1578. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Seshadri VD: Brucine promotes apoptosis in
cervical cancer cells (ME-180) via suppression of inflammation and
cell proliferation by regulating PI3K/AKT/mTOR signaling pathway.
Environ Toxicol. 36:1841–1847. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Ruan H, Zhan YY, Hou J, Xu B, Chen B, Tian
Y, Wu D, Zhao Y, Zhang Y, Chen X, et al: Berberine binds RXRα to
suppress β-catenin signaling in colon cancer cells. Oncogene.
36:6906–6918. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Liu Y, Liu X, Zhang N, Yin M, Dong J, Zeng
Q, Mao G, Song D, Liu L and Deng H: Berberine diminishes cancer
cell PD-L1 expression and facilitates antitumor immunity via
inhibiting the deubiquitination activity of CSN5. Acta Pharm Sin B.
10:2299–2312. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Ham SW, Kim JK, Jeon HY, Kim EJ, Jin X,
Eun K, Park CG, Lee SY, Seo S, Kim JY, et al: Korean Red ginseng
extract inhibits glioblastoma propagation by blocking the Wnt
signaling pathway. J Ethnopharmacol. 236:393–400. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Kim D, Park M, Haleem I, Lee Y, Koo J, Na
YC, Song G and Lee J: Natural product ginsenoside
20(S)-25-Methoxyl-Dammarane-3β, 12β, 20-Triol in cancer treatment:
A review of the pharmacological mechanisms and pharmacokinetics.
Front Pharmacol. 11:5212020. View Article : Google Scholar
|
|
157
|
Yuan Y, Wang J, Xu M, Zhang Y, Wang Z,
Liang L and Sun P: 20(S)-ginsenoside Rh2 as agent for the treatment
of LMN-CRC via regulating epithelial-mesenchymal transition. Biosci
Rep. 40:BSR201915072020. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Hashemi F, Zarrabi A, Zabolian A, Saleki
H, Farahani MV, Sharifzadeh SO, Ghahremaniyeh Z, Bejandi AK,
Hushmandi K, Ashrafizadeh M and Khan H: Novel strategy in breast
cancer therapy: Revealing the bright side of ginsenosides. Curr Mol
Pharmacol. 14:1093–1111. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Sui H, Zhao J, Zhou L, Wen H, Deng W, Li
C, Ji Q, Liu X, Feng Y, Chai N, et al: Tanshinone IIA inhibits
β-catenin/VEGF-mediated angiogenesis by targeting TGF-β1 in
normoxic and HIF-1α in hypoxic microenvironments in human
colorectal cancer. Cancer Lett. 403:86–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Li H, Jeong JH, Kwon SW, Lee SK, Lee HJ
and Ryu JH: Z-Aj oene Inhibits Growth of Colon Cancer by Promotion
of CK1α Dependent β-Catenin Phosphorylation. Molecules. 25:7032020.
View Article : Google Scholar
|
|
161
|
Zhu ML, Zheng Z, Lou EZ, Zhao KT, He SY
and Chen JY: Anti-gastric cancer effects of Z Ajoene and its
molecular mechanisms. Zhongguo Ying Yong Sheng Li Xue Za Zhi.
37:514–519. 2021.In Chinese. PubMed/NCBI
|
|
162
|
Li N, Zeng A, Wang Q, Chen M, Zhu S and
Song L: Regulatory function of DNA methylation mediated lncRNAs in
gastric cancer. Cancer Cell Int. 22:2272022. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Tong Y, Liu Y, Zheng H, Zheng L, Liu W, Wu
J, Ou R, Zhang G, Li F, Hu M, et al: Artemisinin and its
derivatives can significantly inhibit lung tumorigenesis and tumor
metastasis through Wnt/β-catenin signaling. Oncotarget.
7:31413–31428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Wang CZ, Wan C, Luo Y, Zhang CF, Zhang QH,
Chen L, Liu Z, Wang DH, Lager M, Li CH, et al: Effects of
dihydroartemisinin, a metabolite of artemisinin, on colon cancer
chemoprevention and adaptive immune regulation. Mol Biol Rep.
49:2695–2709. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Gong RH, Yang DJ, Kwan HY, Lyu AP, Chen GQ
and Bian ZX: Cell death mechanisms induced by synergistic effects
of halofuginone and artemisinin in colorectal cancer cells. Int J
Med Sci. 19:175–185. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Wen SY, Chen YY, Deng CM, Zhang CQ and
Jiang MM: Nerigoside suppresses colorectal cancer cell growth and
metastatic potential through inhibition of ERK/GSK3β/β-catenin
signaling pathway. Phytomedicine. 57:352–363. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Yang D, Zhang X, Zhang W and Rengarajan T:
Vicenin-2 inhibits Wnt/β-catenin signaling and induces apoptosis in
HT-29 human colon cancer cell line. Drug Des Devel Ther.
12:1303–1310. 2018. View Article : Google Scholar :
|
|
168
|
Yang MH, Ha IJ, Lee SG, Lee J, Um JY and
Ahn KS: Ginkgolide C promotes apoptosis and abrogates metastasis of
colorectal carcinoma cells by targeting Wnt/β-catenin signaling
pathway. IUBMB Life. 73:1222–1234. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Pashirzad M, Johnston TP and Sahebkar A:
Therapeutic effects of polyphenols on the treatment of colorectal
cancer by regulating Wnt β-Catenin signaling pathway. J Oncol.
2021:36195102021. View Article : Google Scholar
|
|
170
|
Howells LM, Berry DP, Elliott PJ, Jacobson
EW, Hoffmann E, Hegarty B, Brown K, Steward WP and Gescher AJ:
Phase I randomized, double-blind pilot study of micronized
resveratrol (SRT501) in patients with hepatic metastases-safety,
pharmacokinetics, and pharmacodynamics. Cancer Prev Res (Phila).
4:1419–1425. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Reabroi S, Chairoungdua A, Saeeng R,
Kasemsuk T, Saengsawang W, Zhu W and Piyachaturawat P: A silyl
andrographolide analogue suppresses Wnt/β-catenin signaling pathway
in colon cancer. Biomed Pharmacother. 101:414–421. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Takahashi K, Kikuchi H, Nguyen VH, Oshima
Y, Ishigaki H, Nakajima-Shimada J and Kubohara Y: Biological
activities of novel derivatives of differentiation-inducing factor
3 from dictyostelium discoideum. Biol Pharm Bull. 40:1941–1947.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Totsuka K, Makioka Y, Iizumi K, Takahashi
K, Oshima Y, Kikuchi H and Kubohara Y: Halogen-Substituted
derivatives of dictyostelium differentiation-inducing factor-1
suppress serum-induced cell migration of human breast cancer
MDA-MB-231 cells in vitro. Biomolecules. 9:2562019. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Efe Ertürk N and Taşcı S: The effects of
peppermint oil on nausea, vomiting and retching in cancer patients
undergoing chemotherapy: An open label quasi-randomized controlled
pilot study. Complement Ther Med. 56:1025872021. View Article : Google Scholar
|
|
175
|
Li X, Bai B, Liu L, Ma P, Kong L, Yan J,
Zhang J, Ye Z, Zhou H, Mao B, et al: Novel β-carbolines against
colorectal cancer cell growth via inhibition of Wnt/β-catenin
signaling. Cell Death Discov. 1:150332015. View Article : Google Scholar
|
|
176
|
Cha PH, Hwang JH, Kwak DK, Koh E, Kim KS
and Choi KY: APC loss induces Warburg effect via increased PKM2
transcription in colorectal cancer. Br J Cancer. 124:634–644. 2021.
View Article : Google Scholar :
|
|
177
|
Ruan Z, Liang M, Lai M, Shang L, Deng X
and Su X: KYA1797K down-regulates PD-L1 in colon cancer stem cells
to block immune evasion by suppressing the β-catenin/STT3 signaling
pathway. Int Immunopharmacol. 78:1060032020. View Article : Google Scholar
|
|
178
|
Cho YH, Ro EJ, Yoon JS, Kwak DK, Cho J,
Kang DW, Lee HY and Choi KY: Small molecule-induced simultaneous
destabilization of β-catenin and RAS is an effective molecular
strategy to suppress stemness of colorectal cancer cells. Cell
Commun Signal. 18:382020. View Article : Google Scholar
|
|
179
|
Savvidou I, Khong T, Cuddihy A, McLean C,
Horrigan S and Spencer A: β-Catenin Inhibitor BC2059 Is efficacious
as monotherapy or in combination with proteasome inhibitor
bortezomib in multiple myeloma. Mol Cancer Ther. 16:1765–1778.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Savvidou I, Khong T, Whish S, Carmichael
I, Sepehrizadeh T, Mithraprabhu S, Horrigan SK, de Veer M and
Spencer A: Combination of histone deacetylase inhibitor
panobinostat (LBH589) with β-Catenin Inhibitor Tegavivint (BC2059)
exerts significant anti-myeloma activity both in vitro and in vivo.
Cancers (Basel). 14:8402022. View Article : Google Scholar
|
|
181
|
Choi JH, Jang TY, Jeon SE, Kim JH, Lee CJ,
Yun HJ, Jung JY, Park SY and Nam JS: The small-molecule Wnt
Inhibitor ICG-001 efficiently inhibits colorectal cancer stemness
and metastasis by suppressing MEIS1 Expression. Int J Mol Sci.
22:134132021. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Yang D, Li Q, Shang R, Yao L, Wu L, Zhang
M, Zhang L, Xu M, Lu Z, Zhou J, et al: WNT4 secreted by tumor
tissues promotes tumor progression in colorectal cancer by
activation of the Wnt/β-catenin signalling pathway. J Exp Clin
Cancer Res. 39:2512020. View Article : Google Scholar
|
|
183
|
Song Q, Han Z, Wu X, Wang Y, Zhou L, Yang
L, Liu N, Sui H, Cai J, Ji Q and Li Q: β-Arrestin1 promotes
colorectal cancer metastasis through GSK-3β/β-Catenin
signaling-mediated epithelial-to-mesenchymal transition. Front Cell
Dev Biol. 9:6500672021. View Article : Google Scholar
|
|
184
|
Li B, Orton D, Neitzel LR, Astudillo L,
Shen C, Long J, Chen X, Kirkbride KC, Doundoulakis T, Guerra ML, et
al: Differential abundance of CK1α provides selectivity for
pharmacological CK1α activators to target WNT-dependent tumors. Sci
Signal. 10:eaak99162017. View Article : Google Scholar
|
|
185
|
Zheng W, Hu J, Lv Y, Bai B, Shan L, Chen
K, Dai S and Zhu H: Pyrvinium pamoate inhibits cell proliferation
through ROS-mediated AKT-dependent signaling pathway in colorectal
cancer. Med Oncol. 38:212021. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Yang W, Li Y, Ai Y, Obianom ON, Guo D,
Yang H, Sakamuru S, Xia M, Shu Y and Xue F: Pyrazole-4-Carboxamide
(YW2065): A therapeutic candidate for colorectal cancer via dual
activities of Wnt/β-Catenin signaling inhibition and AMP-Activated
protein kinase (AMPK) activation. J Med Chem. 62:11151–11164. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Song P, Feng L, Li J, Dai D, Zhu L, Wang
C, Li J, Li L, Zhou Q, Shi R, et al: β-catenin represses miR455-3p
to stimulate m6A modification of HSF1 mRNA and promote its
translation in colorectal cancer. Mol Cancer. 19:1292019.
View Article : Google Scholar
|
|
188
|
Wang Z, Zhou L, Wang Y, Peng Q, Li H,
Zhang X, Su Z, Song J, Sun Q, Sayed S, et al: The CK1δ/ε-AES axis
regulates tumorigenesis and metastasis in colorectal cancer.
Theranostics. 11:4421–4435. 2021. View Article : Google Scholar :
|
|
189
|
Li Y, Rogoff HA, Keates S, Gao Y,
Murikipudi S, Mikule K, Leggett D, Li W, Pardee AB and Li CJ:
Suppression of cancer relapse and metastasis by inhibiting cancer
stemness. Proc Natl Acad Sci USA. 112:1839–1844. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
MacDonagh L, Gray SG, Breen E, Cuffe S,
Finn SP, O'Byrne KJ and Barr MP: BBI608 inhibits cancer stemness
and reverses cisplatin resistance in NSCLC. Cancer Lett.
428:117–126. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Han D, Yu T, Dong N, Wang B, Sun F and
Jiang D: Napabucasin, a novel STAT3 inhibitor suppresses
proliferation, invasion and stemness of glioblastoma cells. J Exp
Clin Cancer Res. 38:2892019. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Beyreis M, Gaisberger M, Jakab M,
Neureiter D, Helm K, Ritter M, Kiesslich T and Mayr C: The cancer
stem cell inhibitor napabucasin (BBI608) shows general cytotoxicity
in biliary tract cancer cells and reduces cancer stem cell
characteristics. Cancers (Basel). 11:2762019. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Jonker DJ, Nott L, Yoshino T, Gill S,
Shapiro J, Ohtsu A, Zalcberg J, Vickers MM, Wei AC, Gao Y, et al:
Napabucasin versus placebo in refractory advanced colorectal
cancer: A randomised phase 3 trial. Lancet Gastroenterol Hepatol.
3:263–270. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Tam BY, Chiu K, Chung H, Bossard C, Nguyen
JD, Creger E, Eastman BW, Mak CC, Ibanez M, Ghias A, et al: The CLK
inhibitor SM08502 induces anti-tumor activity and reduces Wnt
pathway gene expression in gastrointestinal cancer models. Cancer
Lett. 473:186–197. 2020. View Article : Google Scholar
|
|
195
|
Kajino-Sakamoto R, Fujishita T, Taketo MM
and Aoki M: Synthetic lethality between MyD88 loss and mutations in
Wnt/β-catenin pathway in intestinal tumor epithelial cells.
Oncogene. 40:408–420. 2021. View Article : Google Scholar
|
|
196
|
Chen Y, Rao X, Huang K, Jiang X, Wang H
and Teng L: FH535 inhibits proliferation and motility of colon
cancer cells by targeting wnt/β-catenin signaling pathway. J
Cancer. 8:3142–3153. 2017. View Article : Google Scholar :
|
|
197
|
Tu X, Hong D, Jiang Y, Lou Z, Wang K,
Jiang Y and Jin L: FH535 inhibits proliferation and migration of
colorectal cancer cells by regulating CyclinA2 and Claudin1 gene
expression. Gene. 690:48–56. 2019. View Article : Google Scholar
|
|
198
|
Hua F, Shang S, Yang YW, Zhang HZ, Xu TL,
Yu JJ, Zhou DD, Cui B, Li K, Lv XX, et al: TRIB3 Interacts With
β-Catenin and TCF4 to increase stem cell features of colorectal
cancer stem cells and tumorigenesis. Gastroenterology.
156:708–721.e15. 2019. View Article : Google Scholar
|
|
199
|
Shang S, Yang YW, Chen F, Yu L, Shen SH,
Li K, Cui B, Lv XX, Zhang C, Yang C, et al: TRIB3 reduces CD8(+) T
cell infiltration and induces immune evasion by repressing the
STAT1-CXCL10 axis in colorectal cancer. Sci Transl Med.
14:eabf09922022. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Cai X, Wei B, Li L, Chen X, Yang J, Li X,
Jiang X, Lv M, Li M, Lin Y, et al: Therapeutic potential of
apatinib against colorectal cancer by inhibiting VEGFR2-Mediated
Angiogenesis and β-Catenin Signaling. Onco Targets Ther.
13:11031–11044. 2020. View Article : Google Scholar :
|
|
201
|
Or CR, Huang CW and Chang CC, Lai YC, Chen
YJ and Chang CC: Obatoclax, a Pan-BCL-2 inhibitor, downregulates
survivin to induce apoptosis in human colorectal carcinoma cells
via suppressing WNT/β-catenin Signaling. Int J Mol Sci.
21:17732020. View Article : Google Scholar
|
|
202
|
Gan T, Stevens AT, Xiong X, Wen YA, Farmer
TN, Li AT, Stevens PD, Golshani S, Weiss HL, Evers BM and Gao T:
Inhibition of protein tyrosine phosphatase receptor type F
suppresses Wnt signaling in colorectal cancer. Oncogene.
39:6789–6801. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Song W, Ma J, Lei B, Yuan X, Cheng B, Yang
H, Wang M, Feng Z and Wang L: Sine oculis homeobox 1 promotes
proliferation and migration of human colorectal cancer cells
through activation of Wnt/β-catenin signaling. Cancer Sci.
110:608–616. 2019. View Article : Google Scholar :
|
|
204
|
Lepore Signorile M, Grossi V, Di Franco S,
Forte G, Disciglio V, Fasano C, Disciglio V, Fasano C, Sanese P, De
Marco K, et al: Pharmacological targeting of the novel β-catenin
chromatin-associated kinase p38α in colorectal cancer stem cell
tumorspheres and organoids. Cell Death Dis. 12:3162021. View Article : Google Scholar
|
|
205
|
Sheng YH, Wong KY, Seim I, Wang R, He Y,
Wu A, Patrick M, Lourie R, Schreiber V, Giri R, et al: MUC13
promotes the development of colitis-associated colorectal tumors
via β-catenin activity. Oncogene. 38:7294–7310. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Li Q, Wang G, Tao J and Chen W: RNF6
promotes colorectal cancer invasion and migration via the
Wnt/β-catenin pathway by inhibiting GSK3β activity. Pathol Res
Pract. 225:1535452021. View Article : Google Scholar
|
|
207
|
Peng W, Zhang H, Tan S, Li Y, Zhou Y, Wang
L, Liu C, Li Q, Cen X, Yang S and Zhao Y: Synergistic antitumor
effect of 5-fluorouracil with the novel LSD1 inhibitor ZY0511 in
colorectal cancer. Ther Adv Med Oncol. 12:17588359209374282020.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Zhu Y, Gu L, Lin X, Zhang J, Tang Y, Zhou
X, Lu B, Lin X, Liu C, Prochownik EV and Li Y: Ceramide-mediated
gut dysbiosis enhances cholesterol esterification and promotes
colorectal tumorigenesis in mice. JCI Insight. 7:e1506072022.
View Article : Google Scholar :
|
|
209
|
Chen Z, Wu J, Liu B, Zhang G, Wang Z,
Zhang L, Wang K, Fan Z and Zhu P: Identification of cis-HOX-HOXC10
axis as a therapeutic target for colorectal tumor-initiating cells
without APC mutations. Cell Rep. 36:1094312021. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Wang C, Dai J, Sun Z, Shi C, Cao H, Chen
X, Gu S, Li Z, Qian W and Han X: Targeted inhibition of disheveled
PDZ domain via NSC668036 depresses fibrotic process. Exp Cell Res.
331:115–122. 2015. View Article : Google Scholar
|
|
211
|
Pradhan TR and Mohapatra DK: A synthetic
study toward the core structure of (-)-apicularen A. Org Biomol
Chem. 16:8810–8818. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Mauvezin C and Neufeld TP: Bafilomycin A1
disrupts autophagic flux by inhibiting both V-ATPase-dependent
acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome
fusion. Autophagy. 11:1437–1438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Wang J, Mook RA Jr, Ren XR, Zhang Q, Jing
G, Lu M, Spasojevic I, Lyerly HK, Hsu D and Chen W: Identification
of DK419, a potent inhibitor of Wnt/β-catenin signaling and
colorectal cancer growth. Bioorg Med Chem. 26:5435–5442. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
214
|
An T, Gong Y, Li X, Kong L, Ma P, Gong L,
Zhu H, Yu C, Liu J, Zhou H, et al: USP7 inhibitor P5091 inhibits
Wnt signaling and colorectal tumor growth. Biochem Pharmacol.
131:29–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Kumar B, Ahmad R, Sharma S, Gowrikumar S,
Primeaux M, Rana S, Natarajan A, Oupicky D, Hopkins CR, Dhawan P
and Singh AB: PIK3C3 inhibition promotes sensitivity to colon
cancer therapy by inhibiting cancer stem cells. Cancers (Basel).
13:21682021. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Ye GD, Sun GB, Jiao P, Chen C, Liu QF,
Huang XL, Zhang R, Cai WY, Li SN, Wu JF, et al: OVOL2, an inhibitor
of WNT signaling, reduces invasive activities of human and mouse
cancer cells and is down-regulated in human colorectal tumors.
Gastroenterology. 150:659–671.e16. 2016. View Article : Google Scholar
|
|
217
|
Monin MB, Krause P, Stelling R, Bocuk D,
Niebert S, Klemm F, Pukrop T and Koenig S: The anthelmintic
niclosamide inhibits colorectal cancer cell lines via modulation of
the canonical and noncanonical Wnt signaling pathway. J Surg Res.
203:193–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Ren Y, Tao J, Jiang Z, Guo D and Tang J:
Pimozide suppresses colorectal cancer via inhibition of
Wnt/β-catenin signaling pathway. Life Sci. 209:267–273. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Fako V, Yu Z, Henrich CJ, Ransom T, Budhu
AS and Wang XW: Inhibition of wnt/β-catenin signaling in
hepatocellular carcinoma by an antipsychotic drug pimozide. Int J
Biol Sci. 12:768–775. 2016. View Article : Google Scholar :
|
|
220
|
Al-Dali AM, Weiher H and Schmidt-Wolf IGH:
Utilizing ethacrynic acid and ciclopirox olamine in liver cancer.
Oncol Lett. 16:6854–6860. 2018.PubMed/NCBI
|
|
221
|
Liu Q, Zeng A, Liu Z, Wu C and Song L:
Liver organoids: From fabrication to application in liver diseases.
Front Physiol. 13:9562442022. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Caspi M, Wittenstein A, Kazelnik M,
Shor-Nareznoy Y and Rosin-Arbesfeld R: Therapeutic targeting of the
oncogenic Wnt signaling pathway for treating colorectal cancer and
other colonic disorders. Adv Drug Deliv Rev. 169:118–136. 2021.
View Article : Google Scholar
|