Introduction
Salvia miltiorrhizae has been used
effectively as medicine in China for treatment against various
diseases (1). Besides other
compounds with clinical significance purified from Salvia
miltiorrhizae, salvianolic acid A (Sal A), one of the soluble
components has demonstrated various physiological characteristics
including anti-cruor, anti-platelet aggregation and anticancer
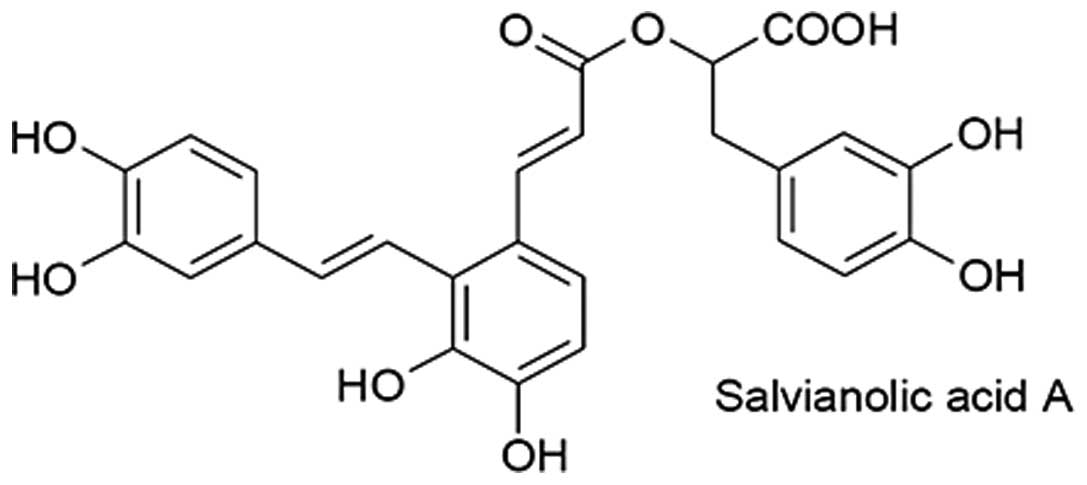
(2). Its molecular formula is shown
in Fig. 1. The combination of Sal A
and other drugs, such as 5-Fluorouracil, Mitomycin C and
Methotrexate, results in the inhibition of cancer growth (3). Preliminary data have indicated that
Sal A is able to inhibit cancer growth in mouse models, including
liver, breast and lung cancers. The mechanism of action of Sal A in
the regulation of cancer cell growth has yet to be elucidated.
Phosphoinositide 3-kinase (PI3K)/Akt signaling
pathway is essential in the regulation of cell survival, growth,
differentiation, apoptosis and autophagy (4). Activation of PI3K phosphorylates
phosphatidylinositol (4,5)-bisphosphate (PIP2) by converting it
into phosphatidylinositol (3,4,5)-triphosphate
(PIP3), thereby preventing cell apoptosis or autophagy. Akt, a key
kinase downstream of PI3K in the signaling pathway, is involved in
regulating cell status (5). There
are three homologies of Akt in mammalian cells. Of those, Akt1 is
pathologically expressed in human cancer cells, suggesting its
direct involvement in cancer onset or development (6). Multiple causes of oncogenic
transformation are involved in the regulation of Akt activity
(7,8). The enhanced Akt activity and the
downstream signaling strength result in the Akt-dependent
suppression of apoptosis and differentiation, but increase the cell
cycle progression (9).
Phosphatase and tensin homolog (PTEN) deleted on
chromosome 10, acts as a phosphatase to dephosphorylate its
substrates, causing tumor suppressor (10). This phosphatase is involved in the
regulation of the cell cycle by inhibiting cell growth and
division. PTEN as a lipid phosphatase is localized on the membrane
(11). In the cytoplasm, PTEN
inhibited PI3K signaling by transforming PIP3 into PIP2 (12). Due to the lack of PTEN in cytoplasm,
PIP3 was accumulated, both Akts and phosphoinositide-dependent
kinase-1 (PDK1) contain a PH domain that binds to membrane-bound
PIP3. Once in the membrane, PDK1 and mammalian target of rapamycin
1 (mTOR1) activate Akt through phosphorylation at various sites.
Thus, the activated Akt positively regulated cell growth or
activity, but negatively regulated cell autophagy and apoptosis
(13). PTEN has been demonstrated
to be the essential molecule regulating PI3K/AKT pathways and
consequently various cell destinations (14).
In this study, the A549 lung cancer cell line and
its response to Sal A treatment was examined. Results showed that
Sal A inhibits growth of the A549 lung cancer cell line, induces
the trend of apoptosis and at the molecular level, regulates
PI3K/AKT signaling pathway by positively mediating PTEN protein
stability.
Materials and methods
Cell culture and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide
(MTT)
A549 cells were purchased from the Shanghai Cell
Research Institute of the China Academy. Cells were cultured in
media with 10% Fas, at 37°C with 5% CO2. Cells were
grown to 80% confluence, then digested with 0.25% trypsin and
transferred into a new plate with fresh media. For MTT, cells were
seeded in 96-well plates with 3×103 cells/well. The
cells were collected at various time points and Sal A doses of 20
μl MTT (5 mg/ml) were added to each well prior to reaction for 4 h.
The supernatant was removed, the pellet was resuspended and 150 μl
dimethyl sulfoxide (DMSO) were added to each well, followed by
agitation at room temperature for 15 min to dissolve the
precipitates. The absorbance (A) value was measured at 490 nm. The
inhibition ratio was calculated as: the inhibition ratio of cell
growth = (1−value A of each sample/value A of control) × 100%.
Flow cytometry analysis
A549 cells were transferred to a 6-well plate with
2×105/ml and cultured at 37°C, with 5% CO2
for 24 h, followed by Sal A treatment at the concentrations of 10,
30 and 60 μg/ml, respectively. The control group was free of the
treatment. After 12 h of culturing, cells were stripped with 0.25%
trypsin and collected by centrifugation (1000 rpm) at 4°C for 5
min. The pellets were washed again with cold phosphate-buffered
saline (PBS), following the addition of 500 μl binding buffer for
cell suspension. The pellets were then mixed with 5 μl Annexin
V-FITC and 5 μl propidium iodide for 10 min in the dark and flow
cytometer was employed to determine the apoptosis ratio.
Measurement of A549 permeability during
Sal A treatment
After 24-h Sal A treatment A549 cells were washed
three times with PBS and mixed with 0.1 μM MitoTracker®
and 10 nM Hoechst for staining. After 30-min reaction at 37°C, the
mixture was washed with PBS three more times, and fixed with
4%-paraformaldehyde for 15 min. For the staining procedure, cells
were washed with PBS three times and 150 μl PBS was then added.
Cell Health Profiling software was applied to analyze the results
of Thermo Cellomics® ArrayScan® VTi.
Western blot analysis
A549 cells were washed twice with ice-cold PBS and
lysed in cold lysis buffer [40 mmol/l HEPES, pH 7.5, 150 mmol/l
NaCl, 1.5 mmol/l MgCl2, 1 mmol/l
ethylenediaminetetraacetic acid (EDTA), 1 mmol/l dithiothreitol and
1 mmol/l fresh phenylmethylsulfonyl fluoride]. Lysates were
incubated for 20 min on ice and centrifuged at 12000 x g for 15
min. The supernatant was collected, and the protein concentration
was determined by Lowry protein assay. The protein was
electrophoresed by sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene
fluoride (PVDF) membranes. The membranes were blocked with 50 g/l
non-fat dried milk in PBST (PBS, 0.5 ml/l Tween-20) for 2 h at room
temperature and incubated overnight at 4°C with the first antibody
against human PTEN, caspase-3 and a phosphorylated antibody against
AKT (first antibodies were purchased from Bioworlde, St. Louis
Park, MN, USA), followed by incubation with an HRP-conjugated
secondary antibody (Bioworlde) at room temperature for 1 h.
Enhanced chemiluminescence (ECL) was used to detect the
results.
Results
Sal A had a negative effect on A549 cell
growth
The A549 lung cancer cell line was used as a model
system to examine the effect of Sal A on its growth. A549 cells
were treated with Sal A at various concentrations of 0.01, 0.1, 1,
10 and 100 μg/ml, respectively. At various time points after
treatment, cells were counted and the results showed that cell
growth decreased at the concentration of ≥10 μg/ml (Table I). With the increase of Sal A
concentration, cell growth was significantly retarded. At 100
μg/ml, the inhibitory ratio of cell growth was ∼90%. At 12 h
treatment, cells became abnormal and floated. At 48 h, cell death
ratio was the highest. However, at 72 h, cells demonstrated
continued growth. This suggested that the growth-inhibitory effect
of Sal A was concentration- and time-dependent in the A549 lung
cancer cell line.
 | Table IA549 growth decreased under Sal A
treatment. |
Table I
A549 growth decreased under Sal A
treatment.
| Inhibitory ratio
(h) | Concentrations
|
|---|
| 0.01 | 0.1 | 1 | 10 | 100 |
|---|
| 24 | 2.95 | 4.10 | 10.00 | 13.29 | 96.79 |
| 48 | 34.14 | 38.96 | 40.76 | 40.11 | 89.95 |
| 72 | 9.00 | 4.36 | 5.37 | 12.93 | 89.98 |
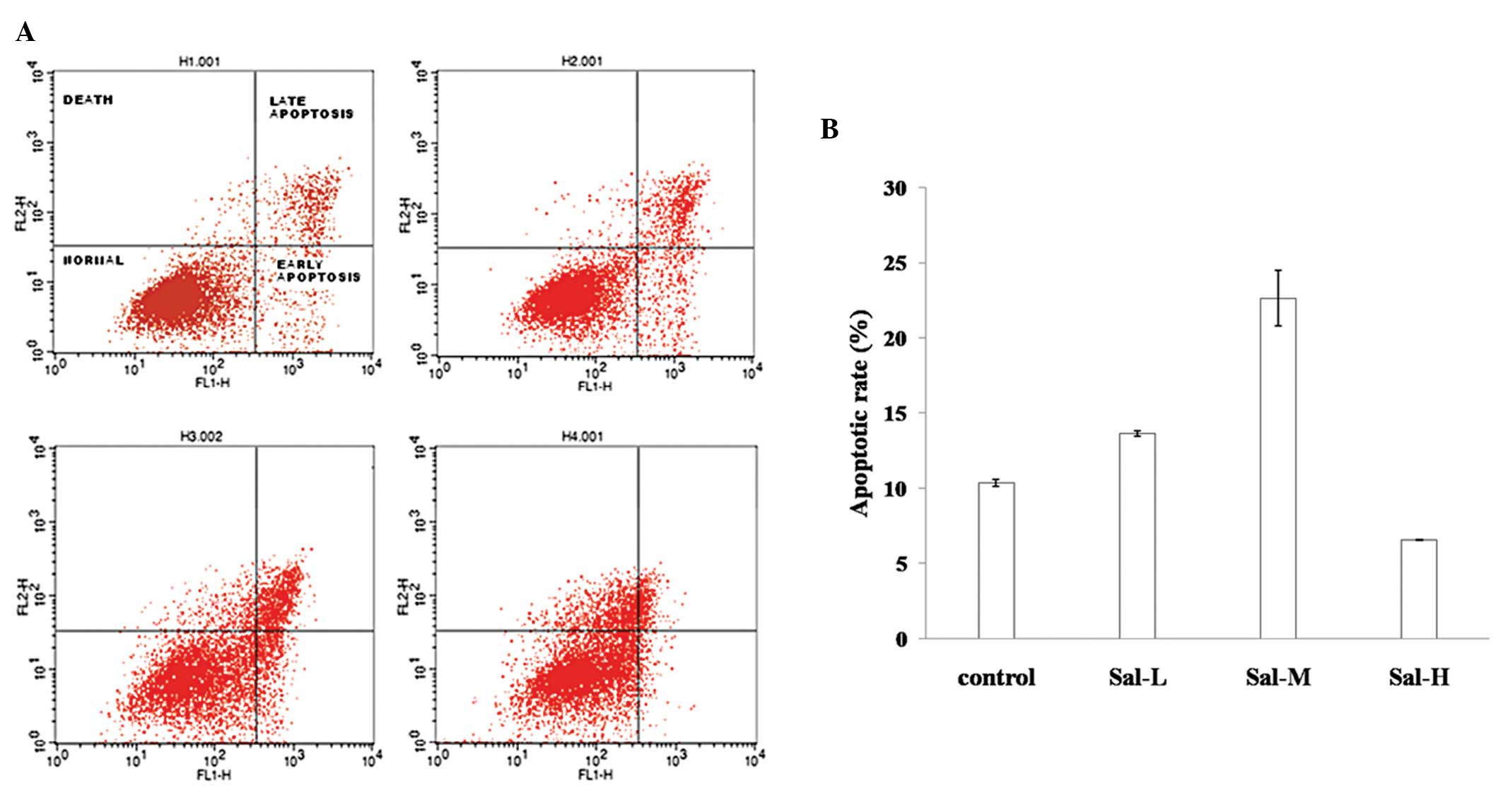
Sal A induced A549 cell apoptosis
There are multiple locations for treated cells
following cell growth inhibition. For instance, cells may undergo
autophagy, apoptosis, cell silence or even cell differentiation. To
determine the biological consequence of Sal A treatment on A549
cells, flow cytometry was used to examine the cell phases during
Sal A treatment. The results showed that cell growth was mostly
blocked at G1 phase (data not shown). In addition, at low
concentration (10 μg/ml), >16% cells underwent apoptosis
(including apoptosis at the early- and late-stages). With the
addition of Sal A the apoptotic rate increased. At high
concentrations (30 μg/ml), the apoptotic rate was only 6%, however,
most cells were floating and dead (Figs. 2A and B), suggesting that apoptosis
is one of the dose-dependent consequences of the Sal A-treated
A549.
Sal A increased mitochondrial membrane
permeability and the cleaved caspase-3 protein level
To better understand whether or not the induced
apoptosis is mitochondrial-dependent, Thermo Cellomics®
ArrayScan® VTi was applied to examine the membrane permeability of
A549 cells during Sal A treatment (Fig.
3A). In the control group treated with MitoTracker®
staining, the cell membrane exhibited a bright red color. Following
the addition of Sal A, the bright red color became weak, indicating
that Sal A promoted mitochondrial membrane permeability.
Furthermore, at the molecular level, Sal A consistently increased
the cleaved caspase-3 protein level (Fig. 3B).
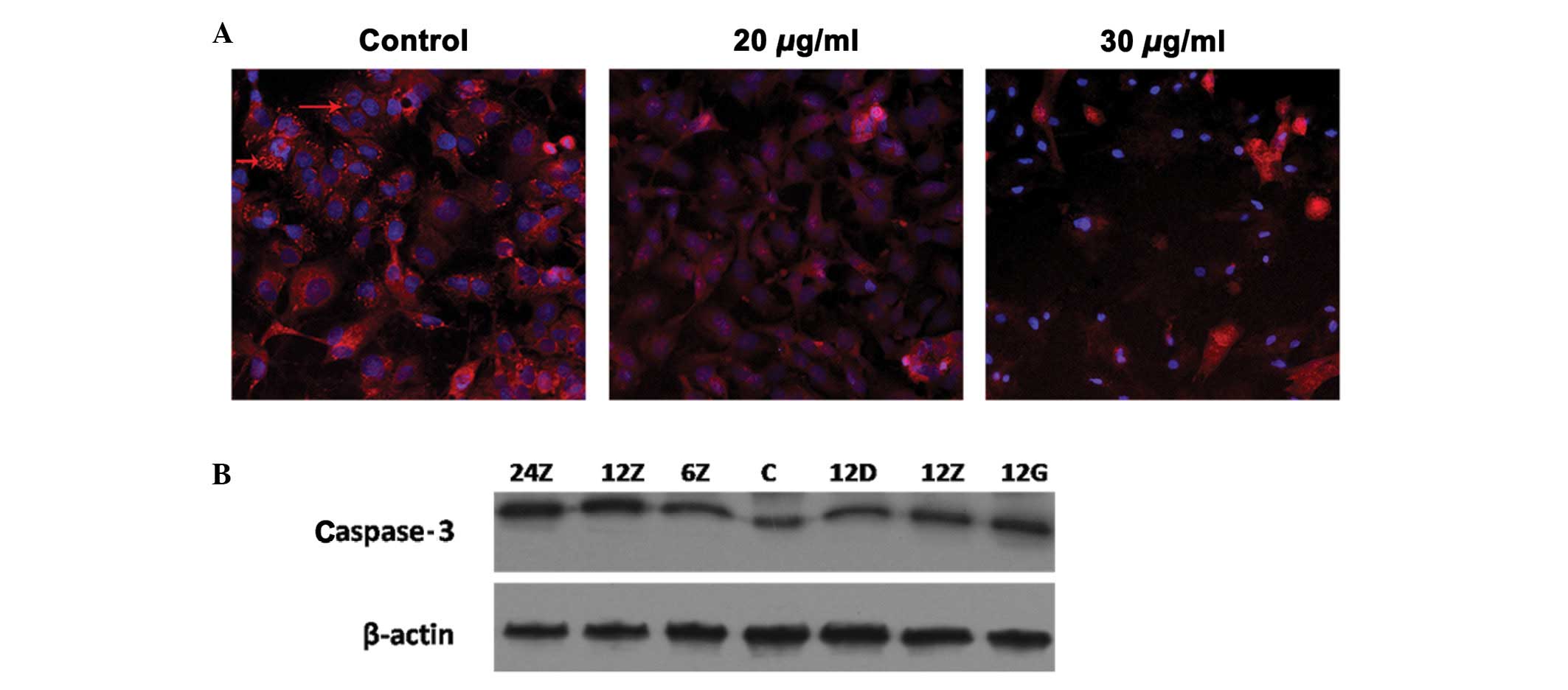 | Figure 3Permeability of mitochondrial membrane
regulated in response to salvianolic acid A (Sal A) treatment. A549
was treated with Sal A at 20 and 30 μg/ml. At 24-h treatment, cells
were stained with MitoTracker® staining, as the control:
the cell membrane exhibited bright red color (red arrow), at 20
μg/ml, the bright red color was weak and at 30 μg/ml, it almost
disappeared. (A) Cells became more condensed. (B) Proteins of A549
treated with the same condition as in Fig. 2. Western blot analysis was used to
detect the cleaved caspase-3 protein level. The concentrations 24,
12 and 6Z indicate medium dose (20 μg/ml) at 24, 12 and 6 h,
respectively. C, Control; 12D, 12Z and 12G are representative of
low (10 μg/ml), medium (20 μg/ml) and high concentrations (30
μg/ml), respectively. |
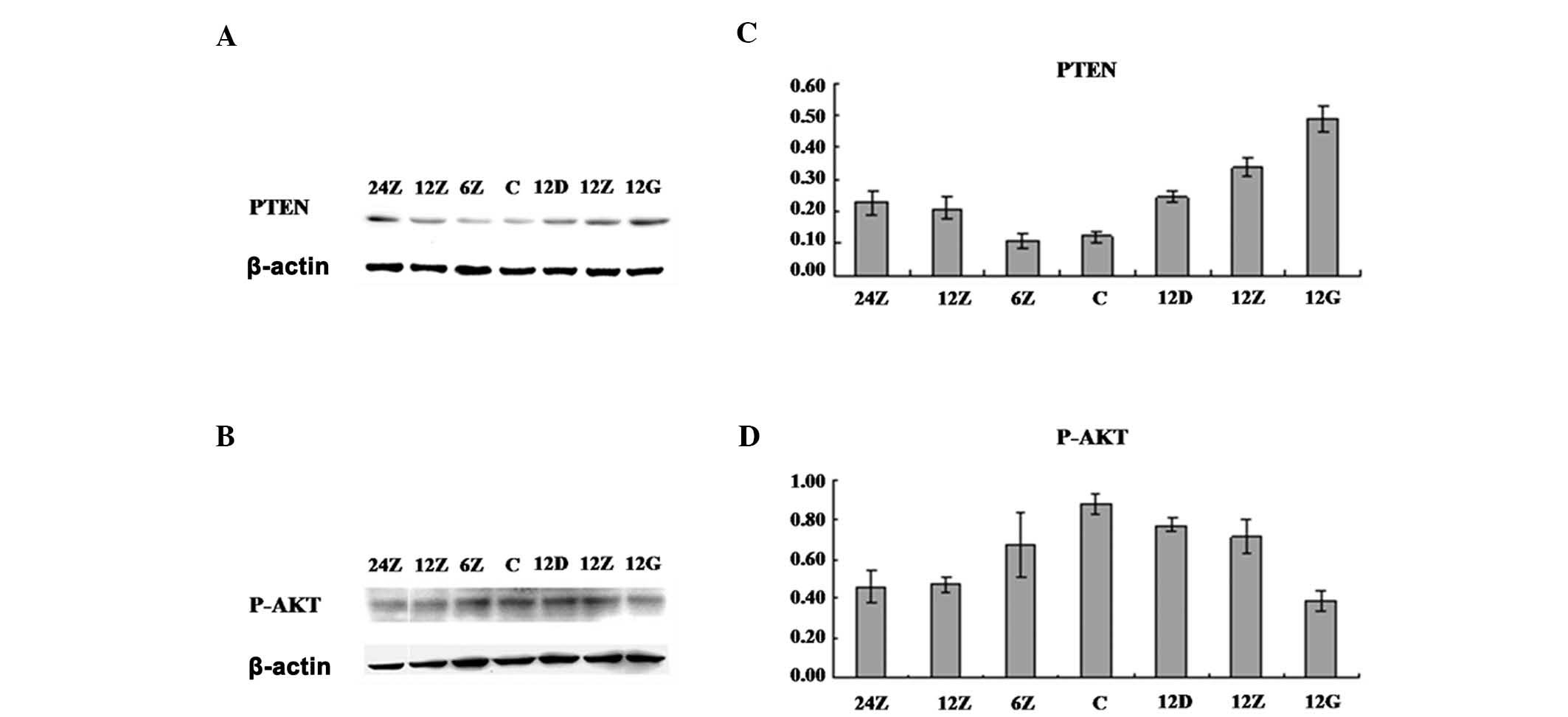
Sal A upregulated the PTEN protein level
and consistently downregulated PI3K/Akt/mTOR signaling
pathways
To understand the molecular role of Sal A in
regulating A549 cell apoptosis, we examined the PI3K/Akt signaling
pathway, which is crucial in regulating cell growth,
differentiation and apoptosis. By detecting protein levels using
western blot analysis, we found that with Sal A treatment, Akt S473
phosphorylation was downregulated at various time points and Sal A
concentrations (Fig. 4). When
examining the potential upstream regulation of Akt, we found that
the PTEN protein level was upregulated in the same treatment. These
results are consistent with cell apoptosis. In addition, to examine
whether or not the regulation of PTEN protein level arose from the
gene transcription or the protein stability, gene chips and qPCR
were performed to examine the PTEN mRNA level (data not shown). The
results suggested that at the mRNA level, no significant difference
was detected, even at high concentrations of Sal A treatment,
indicating that Sal A might positively regulate PTEN protein level
through increasing its stability.
Discussion
Salvia miltiorrhiza is a medical herb that
has been used in China as medicine for the treatment of patients
with various diseases. Recently, several effective compounds of
Salvia miltiorrhiza have been identified as key ingredients
in clinical treatments. To the best of our knowledge, our results
have demonstrated, for the first time, that Sal A induces apoptosis
in human lung cancer cells. Sal A is a hydrophilic molecule and as
yet, it is unknown whether its effect is intra- or extracellular.
The results of this study demonstrate that this acidic substance
has the ability to reduce the viability of lung cancer cells and
induce partial apoptosis. The apoptotic rate varied depending on
the concentration of Sal A treatment. The medium concentration of
Sal A (20 μg/ml) treatment induced a significantly high apoptotic
rate. However, at a high concentration of Sal A, cell growth was
significantly decreased even though the apoptotic rate was only 6%,
suggesting that apoptosis was only one of the consequences of A549
cells responding to Sal A treatment and various cell locations were
likely to be interactive.
To further examine the molecular mechanism of Sal A
action, we studied the regulation of PI3K/Akt signaling pathway. At
the molecular level, the results of repeated experiments suggested
that Sal A treatment significantly increased PTEN protein level and
consistently decreased phosphorylation of Akt at S473. PTEN
expression is reduced in various tumor types, such as lung and
breast cancers (15,16). The regulation of PTEN protein level
is highly involved in the cell growth. The manner in which the PTEN
protein level was upregulated following Sal A treatment remains
unknown. Our gene chips and real time PCR did not show any
difference of PTEN mRNA level at various time points and Sal A
concentrations (data not shown). This observation suggests that Sal
A might directly regulate PTEN protein stability, and thus the
negative regulation of AKT phosphorylation. However, the manner in
which PTEN was modified and its stability regulated has yet to be
elucidated. Although apoptosis was only partially regulated, the
consequence of Sal A treatment and the positive regulation of PTEN
stability in response to Sal A are highly correlated with A549
apoptosis.
Acknowledgements
This study was supported by a project
funded by the Priority Academic Program Development of the Jiangsu
Higher Education Institutions and a grant by the National Natural
Science Foundation of China (Grant no. 81072777).
References
|
1
|
Li Z, Zhang W, Zhao Y, et al: Research
progress of salvianolic acid A. Zhongguo Zhong Yao Za Zhi.
36:2603–2609. 2011.(In Chinese).
|
|
2
|
Zhang W and Lu Y: Advances in studies on
antitumor activities of compounds in Salvia miltiorrhiza.
Zhongguo Zhong Yao Za Zhi. 35:389–392. 2010.(In Chinese).
|
|
3
|
Zhang S, Su J and Zhen Y: Salvianolic acid
A inhibits nucleoside transport and potentiates the antitumor
activity of chemotherapeutic drugs. Yao Xue Xue Bao. 39:496–499.
2004.(In Chinese).
|
|
4
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nielsen-Preiss SM, Silva SR and Gillette
JM: Role of PTEN and Akt in the regulation of growth and apoptosis
in human osteoblastic cells. J Cell Biochem. 90:964–975. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koseoglu S, Lu Z, Kumar C, Kirschmeier P
and Zou J: AKT1, AKT2 and AKT3-dependent cell survival is cell
line-specific and knockdown of all three isoforms selectively
induces apoptosis in 20 human tumor cell lines. Cancer Biol Ther.
6:755–762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shinji K, Sasazawa Y, Imamichi Y, et al:
Caffeine induces apoptosis by enhancement of autophagy via
PI3K/Akt/mTOR/p70S6K inhibition. Autophagy. 7:176–187. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang WC and Hung MC: Induction of Akt
activity by chemotherapy confers acquired resistance. J Formos Med
Assoc. 108:180–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leslie NR and Downes CP: PTEN function:
how normal cells control it and tumour cells lose it. Biochem J.
382:1–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chu EC and Tarnawski AS: PTEN regulatory
functions in tumor suppression and cell biology. Med Sci Monit.
10:RA235–RA241. 2004.PubMed/NCBI
|
|
11
|
Lee JO, Yang H, Georgescu MM, et al:
Crystal structure of the PTEN tumor suppressor: implications for
its phosphoinositide phosphatase activity and membrane association.
Cell. 99:323–334. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Denley A, Gymnopoulos M, Kang S, et al:
Requirement of phosphatidylinositol (3,4,5) trisphosphate in
phosphatidylinositol 3-kinase-induced oncogenic transformation. Mol
Cancer Res. 7:1132–1138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Degtyarev M, De Mazière A, Orr C, et al:
Akt inhibition promotes autophagy and sensitizes PTEN-null tumors
to lysosomotropic agents. J Cell Biol. 183:101–116. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carnero A, Blanco-Aparicio C, Renner O, et
al: The PTEN/PI3K/AKT signaling pathway in cancer, therapeutic
implications. Curr Cancer Drug Targets. 8:187–198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pappas G, Zumstein LA, Munshi A, et al:
Adenoviral-mediated PTEN expression radiosensitizes non-small cell
lung cancer cells by suppressing DNA repair capacity. Cancer Gene
Ther. 14:543–549. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kappes H, Goemann C, Bamberger AM, et al:
PTEN expression in breast and endometrial cancer: correlations with
steroid hormone receptor status. Pathobiology. 69:136–142. 2001.
View Article : Google Scholar : PubMed/NCBI
|