Introduction
Prostate cancer (PCa) is common in Western
populations and is the second leading cause of cancer-related
mortality among males in North America (1). Over the last few years, the morbidity
of PCa in China and other Asian countries has also been on the
increase (2). Drug therapy or
surgery is currently undesirable. Therefore, further investigation
is required to elucidate the mechanisms underlying the development,
progression and prevention of PCa, in order to enable the design of
novel treatment strategies.
Autophagy is a conserved evolutionary process that
is associated with numerous cell responses (3) and is one of the main forms of protein
degradation through the lysosomal pathway. Autophagy is involved in
the majority of long half-life protein degradation. Cells are able
to recycle amino acids and other macromolecular materials for
biosynthesis through the process of autophagy (4). Normal cells possess the autophagic
ability to clear chemical carcinogens and organelles, mainly
mitochondria, that have been damaged due to radiation or oxidative
stress, thereby protecting cell DNA against damage from reactive
oxygen species, ensuring hereditary stability and reducing the
incidence of cell malignant transformation. Thus, autophagy is
crucial in maintaining genome stability (4–6).
Autophagy has been extensively studied in Saccharomyces
cerevisiae, particularly at the genetic level, leading to
identification of the autophagy-related Apg and Aut genes, now
collectively referred to as Atg genes (7,8). LC3
is the mammalian equivalent of yeast Atg8. It exists in two forms,
LC3-I and its proteolytic derivative LC3-II (18 and 16 kDa,
respectively), which are localized in the cytosol (LC3-I) or in
autophagosomal membranes (LC3-II). LC3-II may thus be used to
estimate the abundance of autophagosomes prior to their destruction
through fusion with lysosomes (9,10). In
addition, Beclin-1 is the mammalian orthologue of yeast Atg6
(11). Beclin-1 localizes to the
trans-Golgi network, belongs to the class III phosphatidylinositol
3-kinase complex and is involved in autophagosome formation
(12). It was suggested that
androgen deprivation may induce an autophagic process in LNCaP
cells, possibly causing PCa cells to become androgen-independent
(13,14). However, our study aimed to
investigate the role of autophagy-related genes Beclin-1 and LC3 in
PCa and benign prostatic hyperplasia (BPH) using western blot and
immunohistochemical (IHC) analyses.
Materials and methods
Study population and tissue
specimens
A total of 96 paraffin-embedded BPH tissue samples
were obtained during a two-year period (between July, 2010 and
December, 2012). The samples were divided into two groups, those
from patients who had received 5α-reductase inhibitor (n=55) and
the control group (n=41) and the expression of Beclin-1, LC3, p53,
Bcl-2 and p53 was measured using IHC analysis. The specimens were
provided by the Lu’an Affiliated Hospital of Anhui Medical
University (Lu’an, China) and the Union Hospital of Fujian Medical
University (Fuzhou, China). In addition, protein samples of fresh
specimens from 34 PCa and 50 BPH tissue samples were obtained
during surgery. All the specimens were confirmed by pathology.
Western blot analysis
Fresh specimens, including PCa and BPH tissues, were
obtained during surgery. Cell protein was extracted using IP cell
cracking liquid (Beyotime, Fuzhou, China). Following
electrophoresis, the proteins were loaded onto polyvinylidene
fluoride microporous membranes (Millipore, Billerica, MA, USA).
After blocking of non-specific binding with 5% bovine serum albumin
for 2 h at room temperature, the proteins were identified using a
primary antibody specific to Beclin-l/LC3 (dilution 1:200) in
phosphate-buffered saline (PBS) with Tween-20 under gentle
agitation at 4°C overnight. Western blot analysis was performed
with an anti-rabbit IgG secondary antibody (dilution 1:1000; Bioss,
Beijing, China) and the Enhanced Chemiluminescence Detection system
(ECL; Amersham Pharmacia Biotech, Freiburg, Germany). β-actin was
used as a loading control.
IHC analysis
IHC staining was performed using anti-Beclin-1
rabbit monoclonal antibody (Cell Signaling Technology, Danvers, MA,
USA), anti-LC3 rabbit monoclonal antibody (generously provided by
the Institute of Urology of the Union Hospital of Fujian Medical
University), anti-p53 and anti-Bcl-2 antibodies (Fuzhou Maixin
Biotechnology Development Co., Fuzhou, China). Briefly, the slides
were rehydrated and antigen retrieval was performed by microwave
for 15 min in citrate buffer. The slides were incubated in 3%
hydrogen peroxide for 30 min to block endogenous horseradish
peroxidase (HRP) activity, followed by incubation with normal goat
serum in PBS for 60 min at room temperature. The slides were then
incubated with the primary antibody (dilution 1:100) at 4°C
overnight. Subsequently, the slides were incubated with
biotin-labeled anti-rabbit IgG and preformed avidin-biotin
peroxidase complex. The slides were then counterstained with
hematoxylin, dehydrated and mounted.
Evaluation of degree of antibody
reactivity
The slides were investigated at a magnification of
×400 and a strong brown staining was identified in the nuclei for
p53 and in the cytoplasm for LC3, Beclin-1 and Bcl-2. The
proportion of positively-stained cells was determined in a minimum
of five fields of view. The expression in all the specimens was
classified by two pathologists in our institute according to the
criteria of Ohuchida et al(15) and the percentage of stained normal
or neoplastic epithelial cells was scored as follows: 0, no cells
stained; 1, <20% of cells stained; 2, 20–75% of cells stained;
and 3, >75% of cells stained. The intensity of immunoreactivity
was graded on a scale of 0–3. The total score was the product of
the scores for the intensity and extent of staining. Negative cases
had a score of 0, weakly positive cases had a score of 1–3,
moderately positive cases had a score of 4–6 and strongly positive
cases had a final score of >6.
Statistical analysis
For the results of IHC and western blot analysis,
the Mann-Whitney U test was used to analyze the statistical
contrast of different groups. The correlation coefficients (r and
P-values) among LC3, Beclin-1, Bcl-2 and p53 status were obtained
using the Spearman’s test. P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses were
performed using SPSS software, version 11.5 (SPSS Inc., Chicago,
IL, USA).
Results
Expression of Beclin-1 and LC3
To confirm the role of Beclin-1 and LC3 in the
development and progression of PCa, we investigated their
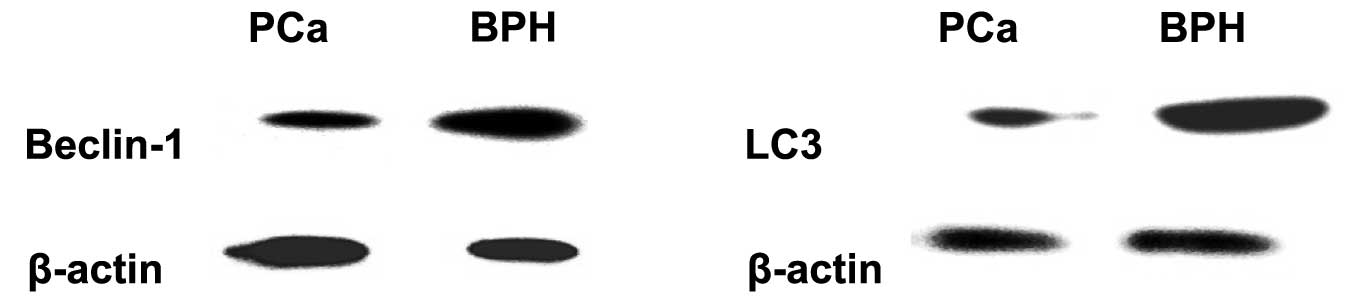
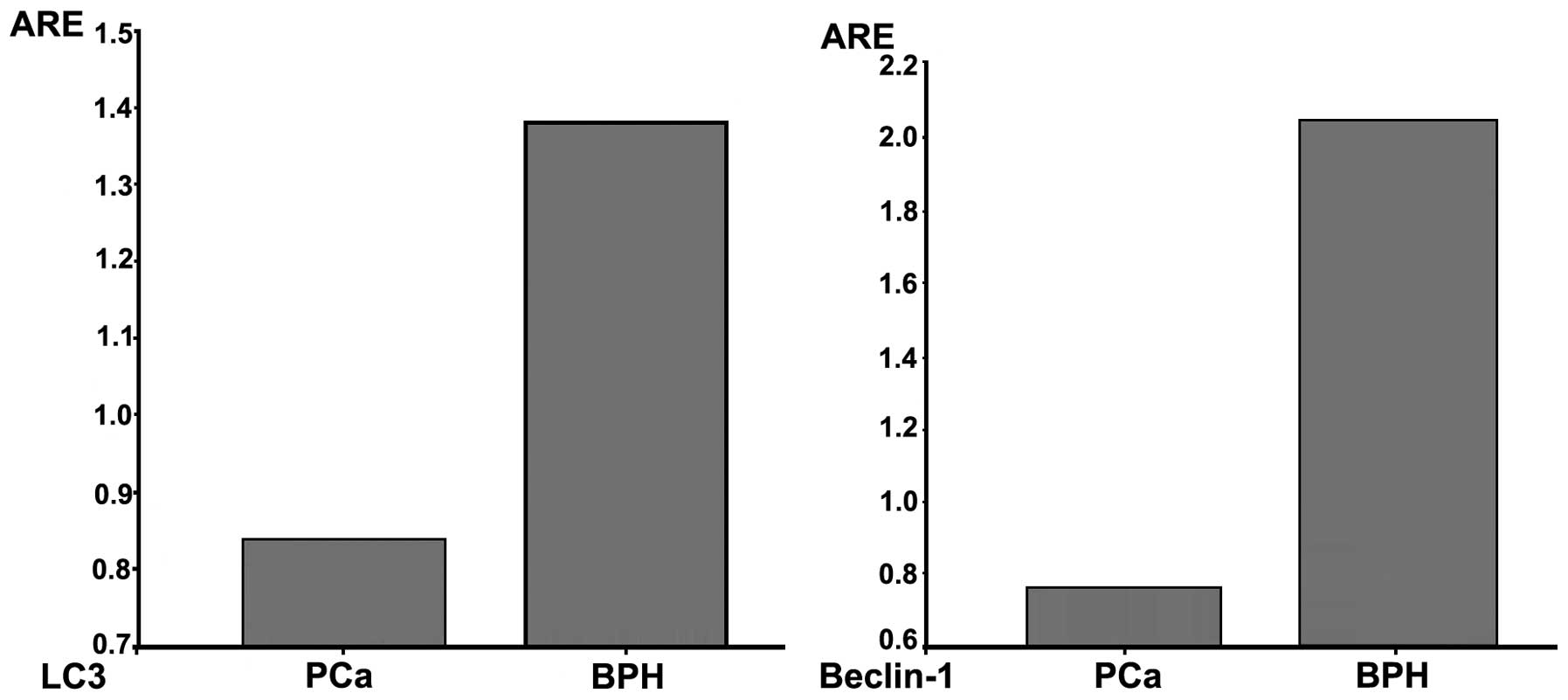
expression by western blot analysis. The average relative
expression value of Beclin-1 in BPH was 2.09±0.12 and that of LC3
was 1.38±0.04. The average relative expression value of Beclin-1
and LC3 in PCa was 0.77±0.06 and 0.84±0.03, respectively. The
statistical analysis demonstrated that the expression of Beclin-1
and LC3 was stronger in BPH compared to that in PCa (P<0.001)
(Figs. 1 and 2). In addition, the Gleason scores of the
34 PCa samples ranged from 6 to 8, with no statistically
significant differences between scores (P>0.05).
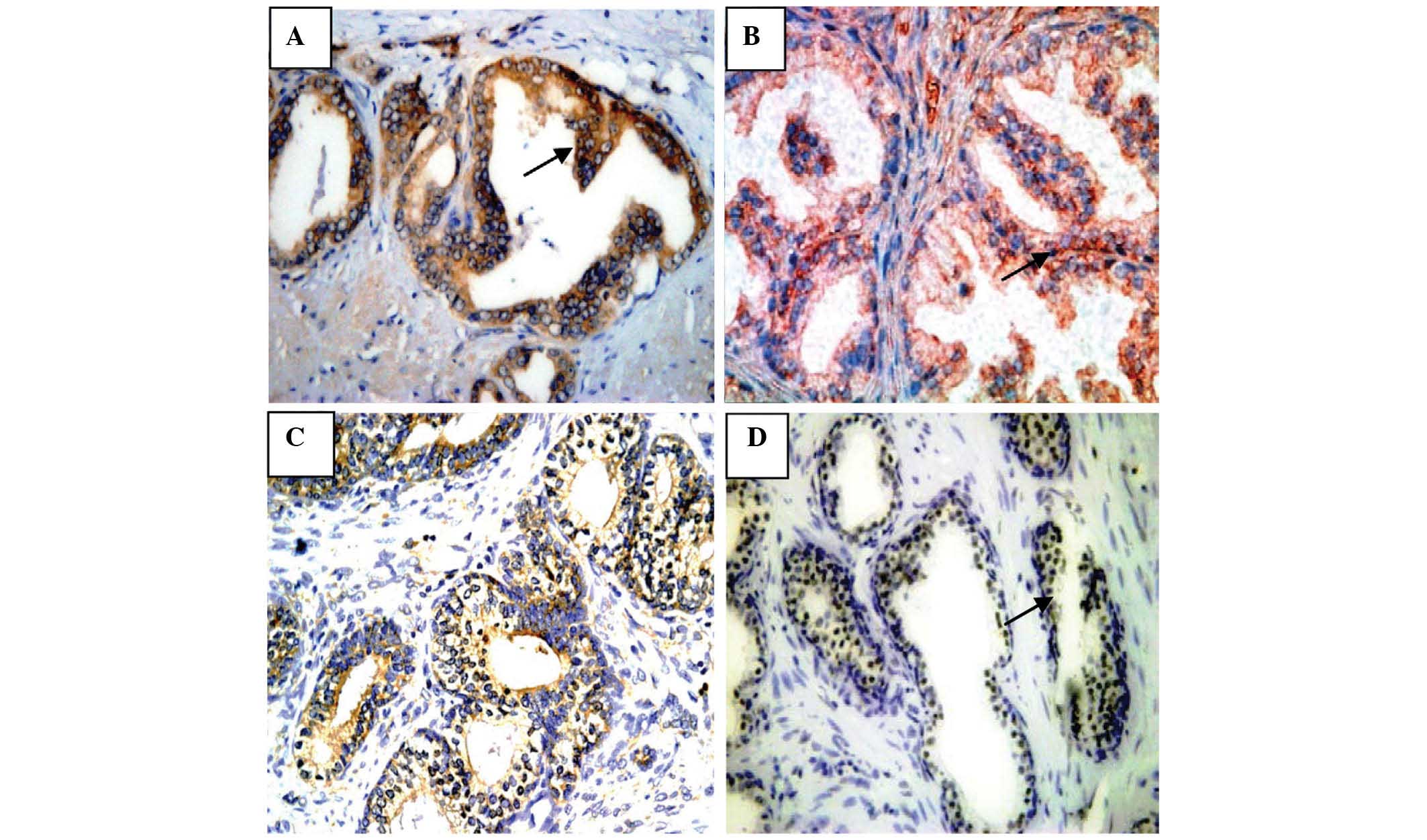
To determine the frequency of expression of the
different types of autophagy-related proteins in BPH between
patients who had received the 5α-reductase inhibitor and the
control group and its correlation with autophagy-related genes and
tumor-related genes, the expression of LC3, Beclin-1, Bcl-2 and p53
was investigated via IHC analysis. The results are presented in
Fig. 3.
Association of LC3 and Beclin-1
expression with the administration of 5α-reductase inhibitor, Bcl-2
and p53
Among the total of 55 BPH patients who had received
the 5α-reductase inhibitor, the positive expression rates of
Beclin-1, LC3, Bcl-2 and p53 were 61.82% (34/55), 52.73% (29/55),
40% (22/55) and 32.7% (18/55), respectively. In the control group,
the positive expression rates were 34.15% (14/41), 36.59% (15/41),
58.54% (24/41) and 51.2% (21/41), respectively. Therefore, the
protein expression of Beclin-1 and LC3 in the 5α-reductase
inhibitor group was significantly higher compared to the control
group (P=0.012 and 0.001), whereas the expression of Bcl-2 and p53
was lower (P=0.031 and 0.045) (Table
I). Beclin-1 expression exhibited a negative correlation with
Bcl-2 (r=−0.402, P<0.001), whereas LC3 expression exhibited a
positive correlation with Beclin-1 (r=0.345, P=0.001) and a
negative correlation with Bcl-2 (r=−0.216, P=0.035).
 | Table IComparison of statistical data of 96
BPH patients with ICH analysis. |
Table I
Comparison of statistical data of 96
BPH patients with ICH analysis.
| Groups | |
|---|
|
| |
|---|
| 5-α reductase
inhibitor | Control | P-value |
|---|
| Beclin-1 | 61.82% (34/55) | 34.15% (14/41) | 0.001 |
| LC3 | 52.73% (29/55) | 36.59% (15/41) | 0.012 |
| Bcl-2 | 40% (22/55) | 58.54% (24/41) | 0.031 |
| p53 | 32.7% (18/55) | 51.2% (21/41) | 0.045 |
Discussion
The phenomenon of autophagy is commonly encountered
in eukaryotic cells. The autophagic process may be initiated by
nutrient starvation, growth factor withdrawal, oxygen deficiency or
protein misfolding. In addition, when amino acid concentration
decreases, autophagy is induced to produce amino acids required for
cell survival. When the supply of amino acids is increased,
autophagy is suppressed. As regards the role of autophagy in
tumorigenesis and tumor progression and its level in different
organs of tumor patients, in the same organ with different types of
tumors, or even in different development stages of the same tumor,
the results vary widely among different studies. Autophagy may
remove damaged organelles, thus contributing to gene stability and
suppressing cell malignant transformation. Furthermore, as a type
of protective mechanism, autophagy protects cancer cells against
damage from a low supply of nutrients, ionizing radiation and
chemotherapy (5,6). In addition, excessive autophagy and
autophagic cell death may inhibit carcinogenesis. Therefore, we
hypothesize that autophagy acts as a double-edged sword regarding
tumorigenesis and tumor progression.
Autophagy-related genes Beclin-l and LC3 may
suppress tumor growth by inducing autophagy. Therefore, they are
considered a potential therapeutic target in cancer management. It
was previously reported that deletion mutations of the Beclin-1
gene were detected in 75% of ovarian cancers, 50% of breast cancers
and 40% of PCas (16). We used
western blot analysis to detect the expression of Beclin-l and LC3
in PCa and BPH tissues. None of the patients had received the
5α-reductase inhibitor, in order to minimize the interference
factors induced by oxygen deficiency and starvation. Following
in vitro sectioning, the surgical specimens were immediately
immersed in cell culture solution and total protein was extracted
after 30 min. The results demonstrated that the expression of
Beclin-1 and LC3 was lower in PCa compared to BPH tissues
(P<0.001), as previously described in the literature and
indicated that the reduction of Beclin-1 and LC3 expression may be
associated with the development of PCa. However, there were no
significant differences between tumors of different Gleason scores
(P>0.05). Therefore, whether autophagy is associated with tissue
differentiation and the prognosis of PCa remains unclear. However,
for PCa patients under androgen ablation treatment, autophagy may
exert a protective effect on PCa cells. It was previously indicated
(13,14) that androgen deprivation may decrease
the phosphorylation of p70S6K. Considering that p70S6K is a readout
of mammalian target of rapamycin (mTOR) activity and a downstream
effector of mTOR, androgen deprivation may induce autophagy in
LNCaP cells. Therefore, PCa cells may exploit the autophagic
pathway to antagonize apoptosis during androgen ablation therapy,
at least for a short period of time, and finally become
androgen-independent. When autophagy was inhibited, LNCaP cell
apoptosis was significantly increased in the absence of
dihydrotestosterone (13,14).
Subsequently, we aimed to investigate whether
autophagy in BPH tissues is induced in the absence of androgen and
elucidate the role of autophagy in the development of BPH. The
number of available studies on the association between autophagy
and BPH is limited. A previous study reported that the number of
autophagosomes was significantly increased in prostate epithelial
cells of castrated rats (17). We
compared the protein expression of LC3 and Beclin-1 using IHC
analysis between patients who had received 5α-reductase inhibitor
and those who had not, since the 5α-reductase inhibitor may reduce
androgen levels in the body (18).
Subsequently, we analyzed its correlation with the
apoptosis-related genes p53 and Bcl-2.
It has already been confirmed that a variety of
tumors are closely associated with the apoptosis-related genes p53
and Bcl-2. Bcl-2 may increase the risk of tumorigenesis and promote
tumor progression through the inhibition of cell apoptosis
(19,20), whereas mutations of wild-type p53,
one of the tumor suppressor genes in normal cells, may promote cell
proliferation and cancer development (21,22).
Wild-type p53 is difficult to detect with IHC analysis. By
contrast, mutant p53 exhibits the characteristics of long half-life
and accumulation in the tumor cell nucleus. Therefore, Oka et
al(23) suggested that all the
p53 protein detected in tumor tissues with IHC was of the mutant
type. In addition, autophagy-related genes are closely associated
with p53 and Bcl-2. The upregulation of Beclin-1 expression in
mammalian cells may induce autophagy; however Bcl-2 binds to
Beclin-1 and inhibits autophagy. Therefore, the cells may escape
death and produce cumulative variation, ultimately leading to
cancer development (24–26). The tumor-suppressor gene p53 may
induce the transcription of insulin-like growth factor binding
protein-3 (IGF-BP3), phosphatase and tensin homolog (PTEN) and
AMP-activated protein kinase (AMPK)-β1 under the condition of
response. These proteins may induce autophagy through
downregulation of the IGF/AKT-1/mTOR signaling pathway, thereby
inhibiting cell growth, division and proliferation (27).
In our study, the expression of Beclin-1 and LC3 in
the 5α-reductase inhibitor group was significantly higher compared
to the control group. However, the expression of Bcl-2 and p53 was
lower (P<0.05), indicating that autophagy was induced in BPH
tissues due to lack of androgen. We hypothesized that the
underlying mechanisms are the result of the androgen reduction as
follows: the gene expression of several transport glycoproteins and
amino acids was restrained and the mTOR signaling pathway was
downregulated, thus enhancing autophagy (28). Ischemia and oxygen deficiency in
prostate cells resulted from reduction of blood flow in the
microcirculation. Under these conditions of nutrient starvation,
autophagy was induced (29).
Futhermore, Beclin-1 expression exhibited a negative correlation
with Bcl-2 (r=−0.402, P<0.001). LC3 expression also exhibited a
negative correlation with Bcl-2 (r=−0.216, P=0.035). The autophagic
process was promoted in the 5α-reductase inhibitor group,
accompanied by a reduction of Bcl-2 and p53.
Therefore, the promotion of autophagy provides
adequate protection to cells from canceration, including BPH tissue
cells in patients who received 5α-reductase inhibitor. Our study
has demonstrated that the expression of Beclin-l and LC3 was
upregulated and associated with p53 and Bcl-2 in BPH patients who
had received 5α-reductase inhibitor. This may reduce the risk of
PCa, in accordance with previous studies suggesting that
finasteride (a type of 5α-reductase inhibitor) may lower the risk
of PCa, although an increase the pathological grade of PCa was
observed (30–32). In conclusion, autophagy-related
genes are crucial in the development of PCa and the canceration of
BPH cells, even in the transitional stage of PCa from
androgen-dependence to androgen-independence.
A previous study demonstrated that autophagy
restraint may enhance apoptosis induced by 5-FU (33). Another study reported that after
receiving a low dose of radiotherapy, the phenomenon of autophagic
vacuole accumulation was observed in the cells of PCa, breast and
colon cancer. Autophagic vacuoles as a defence mechanism may
protect cells against radiation. If the formation of autophagic
vacuoles is inhibited, the mortality rate of cells receiving
radiation may be higher (34).
Kessel et al(35) also
reported that anticancer drugs XK469 and its analogue SH80 may
induce autophagy and cause tumor cell growth stagnation at the G2/M
phase, leading to the elimination of chemoresistance.
In conclusion, autophagy inhibitors or autophagy
revulsants, used alone or combined with other anticancer drugs,
have achieved promising results. The assessment of the status of
Beclin-l and LC3 may prove useful in the treatment of PCa and the
prevention of canceration in BPH patients.
Acknowledgements
This study was supported by Professor Enci Xu
(Institute of Urology, Union Hospital of Fujian Medical University)
through the generous provision of anti-LC3 rabbit monoclonal
antibody. We also gratefully acknowledge Dr Meichun Zhang
(Department of Laboratorian, Lu’an Affiliated Hospital of Anhui
Medical University) for the critical advice.
References
|
1
|
Boring CC, Squires TS and Tong TL: Cancer
statistics, 1992. CA Cancer J Clin. 42:19–38. 1992. View Article : Google Scholar
|
|
2
|
Sim HG and Cheng CW: Changing demography
of prostate cancer in Asia. Eur J Cancer. 41:834–845. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gozuacik D and Kimchi A: Autophagy as a
cell death and tumor suppressor mechanism. Oncogene. 23:2891–2906.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mathew R, Karantza-Wadsworth V and White
E: Role of autophagy in cancer. Nat Rev Cancer. 7:961–967. 2007.
View Article : Google Scholar
|
|
6
|
Mathew R, Kongara S, Beaudoin B, et al:
Autophagy suppresses tumor progression by limiting chromosomal
instability. Genes Dev. 21:1367–1381. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klionsky DJ, Cregg JM, Dunn WA Jr, Emr SD,
Sakai Y, Sandoval IV, Sibirny A, Subramani S, Thumm M, Veenhuis M
and Ohsumi Y: A unified nomenclature for yeast autophagy-related
genes. Dev Cell. 5:539–545. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Reggiori F and Klionsky DJ: Autophagy in
the eukaryotic cell. Eukaryot Cell. 1:11–21. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mizushima N, Yamamoto A, Hatano M,
Kobayashi Y, Kabeya Y, Suzuki K, Tokuhisa T, Ohsumi Y and Yoshimori
T: Dissection of autophagosome formation using Apg5-deficient mouse
embryonic stem cells. J Cell Biol. 152:657–668. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liang XH, Jackson S, Seaman M, Brown K,
Kempkes B, Hibshoosh H and Levine B: Induction of autophagy and
inhibition of tumorigenesis by beclin 1. Nature. 402:672–676. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kihara A, Kabeya Y, Ohsumi Y and Yoshimori
T: Beclin-phosphatidylinositol 3-kinase complex functions at the
trans-Golgi network. EMBO Rep. 2:330–335. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li M, Jiang X, Liu D, et al: Autophagy
protects LNCaP cells under androgen deprivation conditions.
Autophagy. 4:54–60. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chhipa RR, Wu Y and Ip C: AMPK-mediated
autophagy is a survival mechanism in androgen-dependent prostate
cancer cells subjected to androgen deprivation and hypoxia. Cell
Signal. 23:1466–1472. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ohuchida K, Mizumoto K, Ishikawa N, et al:
The role of S100A6 in pancreatic cancer development and its
clinical implication as a diagnostic marker and therapeutic target.
Clin Cancer Res. 11:7785–7793. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aita VM, Liang XH, Murty W, et al: Cloning
and genomic organization of beclin 1, a candidate tumor suppressor
gene on chromosome 17q21. Genomics. 59:59–65. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lian J, Karnak D and Xu L: The
Bcl-2-Beclin 1 interaction in (-)-gossypol-induced autophagy versus
apoptosis in prostate cancer cells. Autophagy. 6:1201–1203. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Roehrborn CG, Bruskewitz R, Nickel GC, et
al: Urinary retention in patients with BPH treated with finasteride
or placebo over 4 years. Characterization of patients and ultimate
outcomes. The PLESS Study Group. Eur Urol. 37:528–536. 2000.
|
|
19
|
Almond JB and Cohen GM: The proteasome: a
novel target for cancer chemotherapy. Leukemia. 16:433–443. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kok SH, Cheng SJ, Hong CY, et al:
Norcantharidin-induced apoptosis in oral cancer cells is associated
with an increase of proapoptotic to antiapoptotic protein ratio.
Cancer Lett. 217:43–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tullo A, D’Erchia AM and Sbisà E: Methods
for screening tumors for p53 status and therapeutic explication.
Expert Rev Mol Diagn. 3:289–301. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Faried A, Faried LS, Kato H, et al:
Targeting p53 tumor suppressor to induce apoptosis and cell cycle
arrest in esophageal cancer cells by novel sugar-cholesterol
compounds. Poster Session. 7:83–84. 2008.
|
|
23
|
Oka K, Nakano T and Arai T: p53CM1
expression is not associated with prognosis in uterine cervical
carcinoma. Cancer. 72:160–164. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei Y, Pattingre S and Levine B: Bcl-2
phosphorylation regulates its antiautophagy function. Monterey:
Keystone Symposium on Autophagy in Health and Disease; 2007
|
|
25
|
Pattingre S, Tassa A, Qu X, et al: Bcl-2
antiapopotic proteins inhibit Beclin 1-dependent autophagy. Cell.
122:927–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu J, Xu Z, Jiang Y, et al: Cryptorchidism
induces mouse testicular germ cell apoptosis and changes in bcl-2
and bax protein expression. J Environ Pathol Toxicol Oncol.
19:25–33. 2000.PubMed/NCBI
|
|
27
|
Feng Z: P53 Regulation of the
IGF-1/AKT/mTOR pathways and the endosomal compartment. Cold Spring
Harb Perspect Biol. 2:a00010572010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Chen SY, Ross KN and Balk SP:
Androgens induce prostate cancer cell proliferation through
mammalian target of rapamycin activation and post-transcriptional
increases in cyclin D proteins. Cancer Res. 66:7783–7792. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lan RZ, Ye ZQ, Yang WM, et al:
Microcirculation changes in the ventral prostates of rats after
castration. Natl J Androl. 11:119–123. 2005.(In Chinese).
|
|
30
|
Thompson IM, Goodman PJ, Tangen CM, et al:
The influence of finastefide on the development of prostate cancer.
N Engl J Med. 349:215–224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kramer BS, Hagerty KL, Justman S, et al;
American Society of Clinical Oncology Health Services Committee;
American Urological Association Practice Guidelines Committee. Use
of 5-alpha-reductase inhibitors for prostate cancer
chemoprevention: American Society of Clinical Oncology/American
Urological Association 2008 Clinical Practice Guideline. J Clin
Oncol. 27:1502–1516. 2009. View Article : Google Scholar
|
|
32
|
Zhu J, Gao JP, Xu AX, et al: The influence
of benign prostatic hyperplasia drugs on incidence and pathology
grading of prostate cancer. Zhonghua Wai Ke Za Zhi. 48:761–763.
2010.(In Chinese).
|
|
33
|
Li J, Hou N, Faried A, et al: Inhibition
of autophagy by 3-MA enhances the effect of 5-FU-induced apoptosis
in colon cancer cells. Ann Surg Oncol. 16:761–771. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Paglin S, Hollister T, Delohery T, et al:
A novel response of cancer cells to radiation involves autophagy
and formation of acidic vesicles. Cancer Res. 61:439–445.
2001.PubMed/NCBI
|
|
35
|
Kessel D, Reiners JJ Jr, Haseldine ST, et
al: The role of autophagy in the death of L1210 leukemia cells
initiated by the new antitumor agents, XK469 and SH80. Mol Cancer
Ther. 6:370–379. 2007. View Article : Google Scholar : PubMed/NCBI
|