Introduction
Magnesium (Mg) is an essential mineral and cofactor
in >300 enzymatic reactions required for numerous structures and
functions in the human body (1).
Homeostatic serum or plasma Mg concentrations are closely
controlled and maintained, ensuring the appropriate activities for
metabolism, neuronal excitability and muscle contraction through a
variety of Mg transporters and ionic channels (2). Numerous Mg-sensitive genes, based on
molecular and functional characterizations, are responsible for
this homeostatic mechanism. In addition, Mg-sensitive gene
expression has been demonstrated to markedly fluctuate during
physiological and pathological events (2,3).
However, the mechanism responsible for regulating plasma Mg
concentrations remains to be determined.
Previous studies have demonstrated that Mg
deficiency reduces exercise performance and endurance (1,4).
Furthermore, Mg supplements have been reported to improve athletic
performance, and a short-term, high-intensity exercise has been
shown to increase plasma Mg concentrations (4). Numerous studies have been conducted on
the beneficial effects of Mg in exercise (4–6), but
the associations between exercise and the gene expression of
Mg-sensitive genes have not been studied. The solute carrier (SLC)
family 41 member A1 (SLC41A1) protein was first identified by
Wabakken et al (7), and then
validated as a Mg transporter by Goytain and Quamme (8). Findings of previous studies have
identified the SLC41A1 protein as a candidate Mg transporter and
confirmed that it most likely mediates the Mg efflux as a
Na+/Mg2+ exchanger (9–11).
Several Mg-sensitive genes have been identified thus far. However,
the extent to which transporter genes fluctuate in the blood cells
during exercise is unknown. In the present study, the role of
SLC41A1 in an animal model was investigated during exercise. The
plasma Mg levels are well-known to increase during exercise, but
the fluctuation of SLC41A1, a Na+/Mg2+
exchanger, expression during exercise has yet to be
established.
The aim of the study was to investigate the
expression of SLC41A1 in mice during treadmill exercise. Blood
samples were obtained at specific time-points and examined by
quantitative polymerase chain reaction (qPCR).
Materials and methods
Animals
Male C57BL/6JNarl mice (n=16, 8 weeks old) were used
in the study, and permission was obtained from the Animal Ethics
Committee of Taichung Veterans General Hospital (La-98593;
Taichung, Taiwan). The blood samples (200 μl) were collected from
the mice using retro-orbital sinus blood collection. It is
generally accepted that withdrawing a blood volume within 10% of
the body weight in an average 25-g mouse is safe. Following the
collection of the first blood sample, the mice remained under the
same conditions for 7 days. Subsequent to this period, eight mice
were intraperitoneally injected with saline and these mice
comprised the control group, whereas another eight mice received
intraperitoneal injection of magnesium sulfate (MgSO4,
90 mg/kg) and served as the Mg group.
The treadmill was equipped with wire loops and
retention sensors at one end of the belt in order for a mild
electric shock to be delivered to encourage the animals to run at
the speed required for the experiment. Mice were shocked at the
lowest level, 0.6 μA, with an inter-pulse interval of <2 sec.
Each mouse was forced to exercise for 3 h at a speed of 15 m/min,
then allowed to recover for an additional 24 h. Each mouse was
allocated a rest period of 1 min and 30 sec every 30 min during the
treadmill exercise (10).
RNA
The blood samples were lysed and the RNA was
purified immediately following collection to prevent the blood from
coagulating and the RNA from undergoing degradation, ensuring
accurate experimental data. Total RNA was isolated from the mice
blood samples using the PureLink™ RNA Mini kit (Ambion®,
Life Technologies, Carlsbad, CA, USA) according to the
manufacturer’s instructions. The RNA integrity assessment was
performed based on the RNA 6000 Nano LabChip (Agilent 2100
bioanalyzer; Agilent, Santa Clara, CA, USA). RNA (1 μg) was
reverse-transcribed using SuperScript® III First-Strand
Synthesis SuperMix for qPCR (Invitrogen, Life Technologies).
Semiquantitative determination of the
SLC41A1 gene expression
Semiquantitative determination of the SLC41A1 gene
expression in blood cells was carried out according to the method
described by Kolisek et al (9). The expression levels of SLC41A1 were
determined by the SYBR-Green based qRT-PCR (SYBR® Green Real-time
PCR Master Mixes; Life Technologies) with specific primers for
mouse SLC41A1 (NM_173854; forward, 5′-TTGGACGCTCGCCTTGCCTG-3′; and
reverse, 5′-TGGTGTGGAACACCTGCGCC-3′) and normalized to the
reference gene, β-actin. A customized oligonucleotide was
synthesized by Invitrogen Life Technologies. Amplifications were
performed using the StepOne Real-time PCR System (Applied
Biosystems, Life Technologies). The PCR conditions for SLC41A1
were: An initial 95°C for 10 min followed by 40 cycles of 95°C for
30 sec and 60°C for 1 min 30 sec, and a melting curve analysis with
95°C for 15 sec and 60°C 1 min, followed by a +0.3°C to 95°C
temperature change with 15 sec at 95°C as the final step. The
samples were assayed in triplicate and the gene expression levels
were determined based on the ΔΔCt method for relative
quantification, which includes efficiency correction, inter- and
intra-assay validation and enables the use of the reference gene,
β-actin (12).
Statistical analysis
Data are presented as means ± SEM. The differences
between and within the groups were evaluated using the Mann-Whitney
test and the Wilcoxon signed-ranks test, respectively. P<0.05
was considered to indicate a statistically significant
difference.
Results
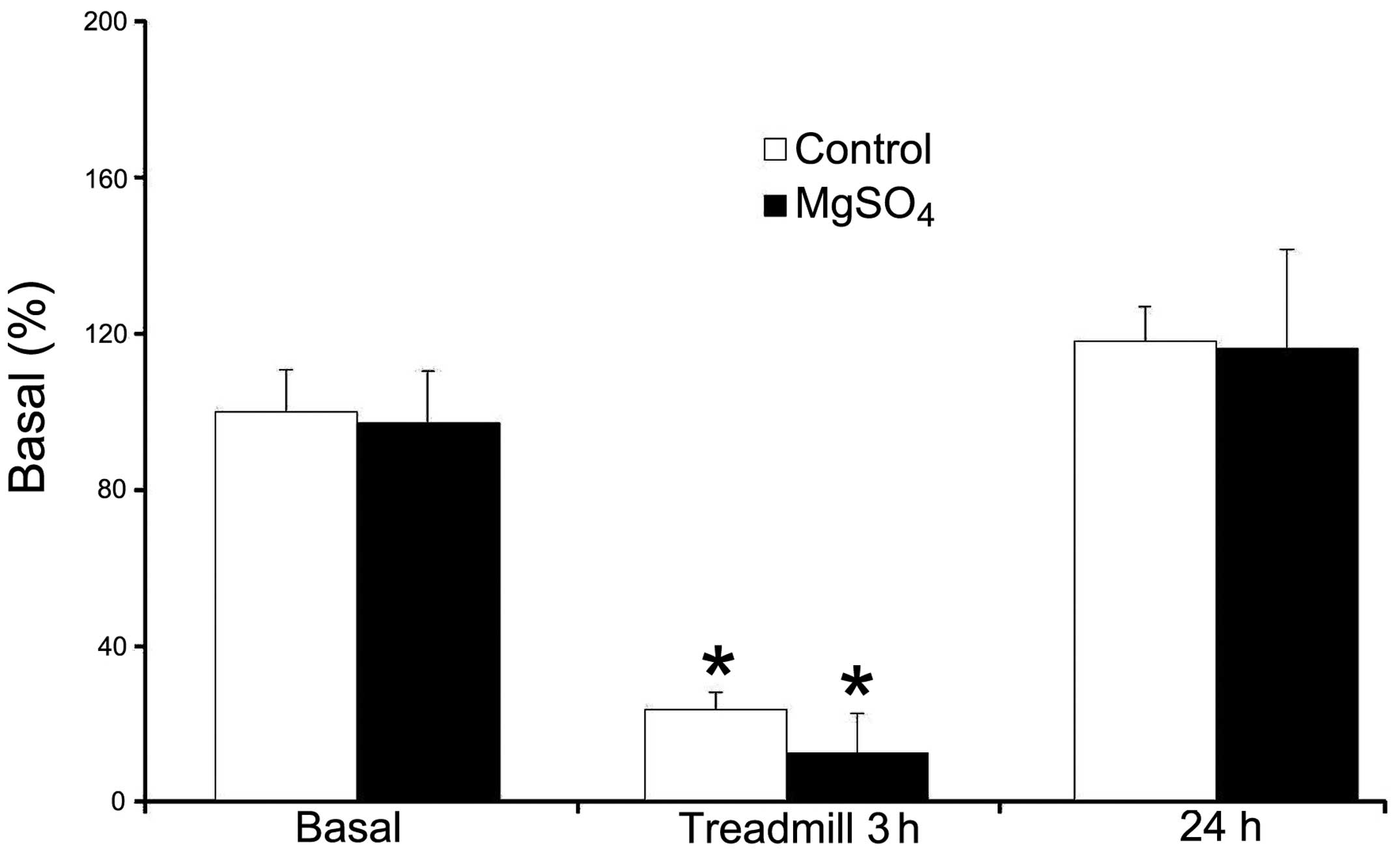
The SLC41A1 gene expression levels were 100.0±10.6
and 96.9±13.7% in the control and Mg groups, respectively (Fig. 1). There was no significant
difference between the two groups. After a 3-h treadmill exercise,
the mean gene expression levels of SLC41A1 were 23.6±4.6 and
12.6±10.2% in the control and Mg groups, respectively (Fig. 1). There was a significant
downregulation of SLC41A1 gene expression following exercise when
compared with prior to exercise in the two groups.
There was a 76.4% decrease in the expression levels
of SLC41A1 in the control group and a 87.0% decrease in the Mg
group as compared to the basal levels, resepectively. However, this
difference was not significant. After 24 h, the gene expression
levels of SLC41A1 were returned to 118.1±9.0 and 116.1±25.5% in the
control and Mg groups, respectively.
Discussion
In the present investigation, the SLC41A1 gene
expression levels were significantly downregulated following
exercise, decreasing between 76 and 87%, but then returning to the
basal levels 24 h after exercise. The mice had much lower SLC41A1
gene expression levels in the Mg group than the control group
following exercise, but this was not statistically significant.
Thus, the supplementation of Mg may lead to a decreased SLC41A1
gene expression following exercise. By contrast, this may be
attributed to small sample sizes or individual differences among
the mice in the present study.
The data show that the SLC41A1 gene was
downregulated in the blood cells of mice after 3 h of exercise.
Thus, SLC41A1 gene expression appears to be involved in the
physiological response to exercise. Recently, Nestler et al
(13) demonstrated that SLC41A1
gene expression is correlated to blood pressure and Mg homeostasis
in pregnant women. A downregulation of SLC41A1 gene expression was
interpreted as Mg deficiency in leukocytes during pregnancy.
Although the interpretation remains speculative, the data of the
present study are corresponds with the hypothesis of Nestler et
al (13). The hemodynamic
responses of the heart rate and blood pressure to exercise have
been extensively studied previously (4). An exaggerated blood pressure response
during exercise has been considered to be an indicator of the
exercise performance or cardiovascular risks. Therefore, a 70–80%
downregulation of SLC41A1 gene expression and highly elevated blood
pressure during exercise appears to be parallel and acceptable. A
possible explanation for the downregulated expression of the
SLC41A1 gene during exercise could be that a decreased SLC41A1
expression in the blood is an obligatory component of the metabolic
response to the immediate demands of exercise. The Mg content in
leukocytes is higher than that of mononuclear blood cells, which
may represent the downregulation of SLC41A1 expression in blood. In
our previous studies, plasma Mg concentrations increase in various
treadmill exercise modes (5,6). Thus
far, it may be concluded that the SLC41A1 gene expression is
associated with blood pressure and blood Mg content. However,
whether SLC41A1 expression is Mg-dependent remains unclear.
The rapid return to the basal levels within 24 h
after exercise implies that the SCL41A1 gene may be a fast-acting
gene that is highly responsive to exercise in mice. However, the
factors that facilitate Mg transportation also do not rely solely
on the SLC41A1 gene, and there are at least nine Mg-sensitive genes
(CNNM2, TRPM6, TRPM7, N33, SLC41A1, SLC41A2, SLC41A3, MagT1 and
NIPA1) that have been shown to be involved in manipulating intra-
and extracellular Mg transportation (2). In the present study, the SLC41A1 gene
was downregulated, resulting in an increase and then decrease of Mg
levels in the blood during the treadmill exercise. Although just
one gene was analyzed in the study, the effects of other Mg
transporter genes in response to exercise require further
investigation.
Thus, we hypothesize that a downregulated SLC41A1
gene by means of non-exercise or Mg supplementation leads to an
increase of Mg in the blood that could confer benefits similar to
those observed with exercise, as consumption of various Mg
supplements by Mg-deficient subjects is known to have beneficial
effects (1–4). Mg is also widely used in the treatment
of diseases, including but not limited to, asthma, heart disease,
blood clots, hypertension, osteoporosis and hypoglycemia.
Determining whether the part-knockdown of the SLC41A1 gene in in
vitro or animal models is likely to lead to the similar effects
as achieved by consumption of Mg supplements is useful. Future
investigations should therefore be conducted to determine whether
these similar effects exist.
The findings of the present study suggest that an
increased SLC41A1 expression leads to the excess transportation of
Mg out of the blood, which subsequently results in Mg deficiency.
Mg-deficient mice could be examined in future experiments to
determine whether they show higher rates of SLC41A1 expression, and
whether the upregulated expression results in decreased plasma Mg
levels leading to a poor health status.
There were certain limitations in the present study
and several potential confounders. First, subsequent to obtaining
an initial 200 μl of blood from the mice, the animals were allowed
to rest for a week. However, subsequent to obtaining 200 μl of
blood following exercise, the mice were allowed to rest for 24 h
prior to obtaining the third blood sample. The total blood volume
in a mouse weighing 25 g is <2.0 ml. Relatively large amounts of
blood (>400 μl) collected in a short period may have also
affected the SLC41A1 gene expression in the blood. In addition,
plasma or serum Mg levels were not determined, although results of
previous experiments have shown that plasma Mg levels in the blood
rise ~20% following exercise (5,6).
Furthermore, the SLC41A1 gene expression may vary in different
parts of the body. Although the plasma SLC41A1 gene expression was
downregulated, the gene expression in the kidney or small intestine
may be different, and even within the kidney the SLC41A1 gene
expression may vary in the cortex or medulla. Due to the
potentially high variability in SLC41A1 gene in different parts of
the body, the SLC41A1 gene expression in blood may therefore not be
completely reliable without further investigation.
In conclusion, the SLC41A1 gene expression in mice
was downregulated following exercise and returned to the basal
levels within 24 h. It is known that Mg levels in the blood
increase during exercise. Therefore, we hypothesize that the
SLC41A1 may play an important role in the interaction of blood Mg
levels. However, the exact mechanism of the association between the
SLC41A1 gene expression and blood Mg status during exercise remains
unclear.
Acknowledgements
This study was supported by Taichung Veterans
General Hospital, Taichung, Taiwan, R.O.C (TCVGH-1037313C).
References
|
1
|
Finstad EW, Newhouse IJ, Lukaski HC, et
al: The effects of magnesium supplementation on exercise
performance. Med Sci Sports Exerc. 33:493–498. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Romani AM: Cellular magnesium homeostasis.
Arch Biochem Biophys. 512:1–23. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cinar V, Polat Y, Mogulkoc R, et al: The
effect of magnesium supplementation on glucose and insulin levels
of tae-kwan-do sportsmen and sedentary subjects. Pak J Pharm Sci.
21:237–240. 2008.PubMed/NCBI
|
|
4
|
Bohl CH and Volpe SL: Magnesium and
exercise. Crit Rev Food Sci Nutr. 42:533–563. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen YJ, Chen HY, Wang MF, et al: Effects
of magnesium on exercise performance and plasma glucose and lactate
concentrations in rats using a novel blood-sampling technique. Appl
Physiol Nutr Metab. 34:1040–1047. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cheng SM, Yang LL, Chen SH, et al:
Magnesium sulfate enhances exercise performance and manipulates
dynamic changes in peripheral glucose utilization. Eur J Appl
Physiol. 108:363–369. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wabakken T, Rian E, Kveine M and Aasheim
HC: The human solute carrier SLC41A1 belongs to a novel eukaryotic
subfamily with homology to prokaryotic MgtE
Mg2+transporters. Biochem Biophys Res Commun.
306:718–724. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goytain A and Quamme GA: Functional
characterization of human SLC41A1, a Mg2+transporter
with similarity to prokaryotic MgtE Mg2+transporters.
Physiol Genomics. 21:337–342. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kolisek M, Launay P, Beck A, et al:
SLC41A1 is a novel mammalian Mg2+carrier. J Biol Chem.
283:16235–16247. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mandt T, Song Y, Scharenberg AM and Sahni
J: SLC41A1 Mg(2+) transport is regulated via Mg(2+)-dependent
endosomal recycling through its N-terminal cytoplasmic domain.
Biochem J. 439:129–139. 2011.
|
|
11
|
Kolisek M, Nestler A, Vormann J and
Schweigel-Röntgen M: Human gene SLC41A1 encodes for the Na+/Mg(2)+
exchanger. Am J Physiol Cell Physiol. 302:C318–C326.
2012.PubMed/NCBI
|
|
12
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nestler A, Rylander R, Kolisek M, et al:
Blood pressure in pregnancy and magnesium sensitive genes. Preg
Hyper: An Int J Women’s Card Health. 4:41–45. 2014.
|