Introduction
The genus Fusarium members are ubiquitous
fungi frequently found in soils and plants (1). Fusarium species have been
distinguished as a cause of localized infections (1,2). Due to bone
marrow grafts and immunosuppressive treatment, invasive
Fusarium infections have increased during the last few
decades. The host immunological status and the level of the
infection are the most significant aspects for the clinical effect
of Fusarium infections (3,4).
The Fusarium species are a widely spread
phytopathogen found in an extensive variety of hosts (5). It causes wilts and root rot disease,
which produces secondary metabolites such as T2-toxin, zearalenone
and trichothecene, causing huge economic problems through losing
crops (6,7). The genus Fusarium is seldom able
to cause human infections, such as onychomycosis, skin infections
or keratitis. The Fusarium genus is one of the most
heterogeneous fungi and is difficult to classify. Conversely,
identification at the species level is required for biological and
epidemiological reasons.
Conventional diagnostic techniques for
identification of the Fusarium species in culture or in
infected tissues are based on morphological features. This
procedure is time consuming, and it can frequently be difficult to
discriminate between similar species. Molecular approaches are more
sensitive and quicker. Furthermore, they are employed to the
specific detection of the Fusarium species. For these
reasons, the molecular biological technique has been established in
Fusarium systematics and the molecular variation at the DNA
level has been investigated in numerous studies (8–10). The use
of polymerase chain reaction (PCR) with primers targeted to the
internal transcribed spacer (ITS) region of the ribosomal DNA
(11,12)
for identification of the Fusarium species has been
reported. The ITS region sequences have shown to be highly variable
in Fusarium genus (13).
PCR-restriction fragment length polymorphism (RFLP)
analysis is a useful method in the detection of DNA polymorphism in
objective sequences. In the present study, the amplified ITS region
of rDNA was digested with 2 restriction endonucleases for genetic
variation among Fusarium spp. The aim of the study was to
identify the phylogenetic associations and usefulness of the ITS
region as a genetic marker within the most clinically important of
the Fusarium species.
Materials and methods
Microorganisms
A total of 50 strains of Fusarium spp. were
used in the study including environmental, clinical and reference
isolates. The following strains were used as a reference: F.
solani PFCC 5284, F. solani PFCC 5285, F.
oxysporum PFCC 30067, F. oxysporum PFCC 5115, F.
verticillidea PFCC 53–131, F. verticillidea PFCC 15–89,
F. proliferatum PFCC 48–125, F. proliferatum PFCC
12–86 and F. fujikuri PFCC 5144. Enviromental strains were
recovered from soil. Two strains were clinical strains, including
F. solani PFCC 5284.
DNA extraction
In total, 100 ml of YEPD medium in Erlenmeyer flasks
was inoculated with 1 ml of thick spore suspension. The flasks were
incubated at 200 rpm under agitation at 25°C for 72 h to obtain
mycelium growth. The mycelia were harvested, washed with 0.5 M EDTA
and sterile dH2O and ground into a fine powder with a
pestle and mortar using liquid nitrogen.
Approximately 100 mg of the mycelium powder was
transferred into a 1.5-ml tube and 400 µl of lysis buffer [100 mM
Tris-HCl (pH 8.0), 30 mM EDTA (pH 8.0) and 5% SDS w/v] was
added.
Following the incubation of the tubes at 100°C for
20 min, 150 µl of 3 M acetate potassium was added. The suspension
was maintained for 10 min at −20°C, and centrifuged at 14,000 × g
for 10 min in 4°C. The supernatant was transferred to a 1.5-ml
Eppendorf tube (Eppendorf AG, Hamburg, Germany), and subsequently,
250 µl of phenol-chloroform-isoamyl alcohol (25:24:1, v/v) was
added and the mixture was briefly vortexed and centrifuged at
14,000 × g for 10 min. Subsequent to transferring the upper aqueous
phase to a new 1.5-ml tube, 250 µl chloroform-isoamyl alcohol was
added. The tubes were briefly vortexed and centrifuged in 4°C at
14,000 × g for 10 min. The supernatant was transferred to a new
tube, an equal volume of ice-cold 2-propanol was added and the
mixture was incubated at −20°C for 10 min and subsequently
centrifuged at 14,000 × g for 10 min. The upper aqueous phase was
removed and the pellet was washed with 300 µl of ethanol 70%. The
ethanol was removed and the DNA pellet was air-dried and dissolved
in 50 µl of dH2O.
PCR amplification
The primer sets (ITS1, 5′-TCCGTAGGTGAACCTGCGG-3′)
and (ITS4, 5′-TCCTCCGCTTATTGATATGC-3′) were used to amplify an
~600-base pair (bp) DNA fragment of the ITS region. PCR reactions
were prepared to a final volume of 50 µl, containing reaction
buffer, 2.2 mM MgCl2, 200 µM of each dNTP, 2.5 unit of
Taq DNA polymerase (CinnaGen, Karaj, Iran), a 30-ng DNA template
and 50 pmol of each primer.
An initial denaturation step for 5 min at 95°C was
followed by 30 cycles of denaturation at 94°C for 40 sec, annealing
at 58°C for 40 sec and extension at 72°C for 40 sec, with a final
extension at 72°C for 5 min. The PCR product was run on a 1%
agarose gel in Tris-base, acetic acid and EDTA (TAE) buffer, and
stained with ethidium bromide. The PCR amplification of the ITS
region resulted in ~595-bp of fragment.
RFLP analysis
Two restriction enzymes were analyzed. HaeIII
and SmaI had restriction sites in the ITS region of the
Fusarium species. The reaction mixture for each enzyme was
carried out in a total volume of 20 µl containing 10 units of the
enzyme, 2 µl of the related buffer, 5 µl of the PCR product and
Ultrapure water to create the 20-µl volume. Digested PCR products
were subjected to electrophoresis on a 1.5% agarose gel in TAE
buffer, and stained with ethidium bromide.
Results
PCR amplification of the ITS
regions
PCR amplification of the ITS region with primers
ITS1 and ITS4 resulted in an ~550-bp band (Fig. 1). The fragment was obtained from all 50
Fusarium strains. The ITS-region products were sequenced
from several isolates, including the reference strains. A Basic
Local Alignment Search Tool search showed that the ITS PCR products
from 5 medically significant Fusarium reference strains,
which were F. solani, F. oxysporum, F.
verticillidea, F. proliferatum and F. fujikuri,
exhibited a 99% homology with the associated sequences in the
GenBank.
Restriction patterns for the Fusarium
strains
Different restriction patterns were obtained
following digestion with the HaeIII and SmaI enzymes
for the reference Fusarium strains, including F.
solani, F. oxysporum, F. verticillidea, F.
proliferatum and F. fujikuri (Table I and Fig.
2).
 | Figure 2.Agarose gel electrophoresis of the
internal transcribed spacer region base pair (bp) products of the
Fusarium species following digestion with HaeIII
(lanes 1–6) and SmaI (lanes 7–12). M, 100 bp ladder; lane 1,
F. oxysporum PFCC 5115; lane 2, F. verticillidea PFCC
15–89; lane 3, F. proliferatum PFCC 12–86; lane 4, F.
verticillidea PFCC 53–131; lane 5, F. fujikuri PFCC
5144; lane 6, F. solani PFCC 5284; lane 7, F.
oxysporum PFCC 5115; lane 8, F. verticillidea PFCC
15–89; lane 9, F. proliferatum PFCC 12–86; lane 10, F.
verticillidea PFCC 53–131; lane 11, F. fujikuri PFCC
5144; lane 12, F. solani PFCC 5284. |
 | Table I.Restriction fragment size of the ITS
region of the Fusarium species following digestion with
HaeIII and SmaI, according to the sequencing of the
strains. |
Table I.
Restriction fragment size of the ITS
region of the Fusarium species following digestion with
HaeIII and SmaI, according to the sequencing of the
strains.
| Fusarium
species | ITS size prior to
digestion, bp | HaeIII,
bp | SmaI,
bp |
|---|
| F.
oxysporum | 550 | 347, 88, 86 | 550 |
| F.
verticillidea | 550 | 347, 89, 85 | 550 |
| F.
proliferatum | 570 | 281, 90, 89,
77 | 570 |
| F.
fujikuri | 550 | 281, 91, 77,
61 | 324, 210 |
| F.
solani | 570 | 241, 123, 91,
65 | 325, 217 |
The restriction patterns of 6 environmental
Fusarium strains with HaeIII and SmaI are
shown in Fig. 3. Strain E42 was
recognized as F. oxysporum, based on the sequencing and
restriction enzymes pattern.
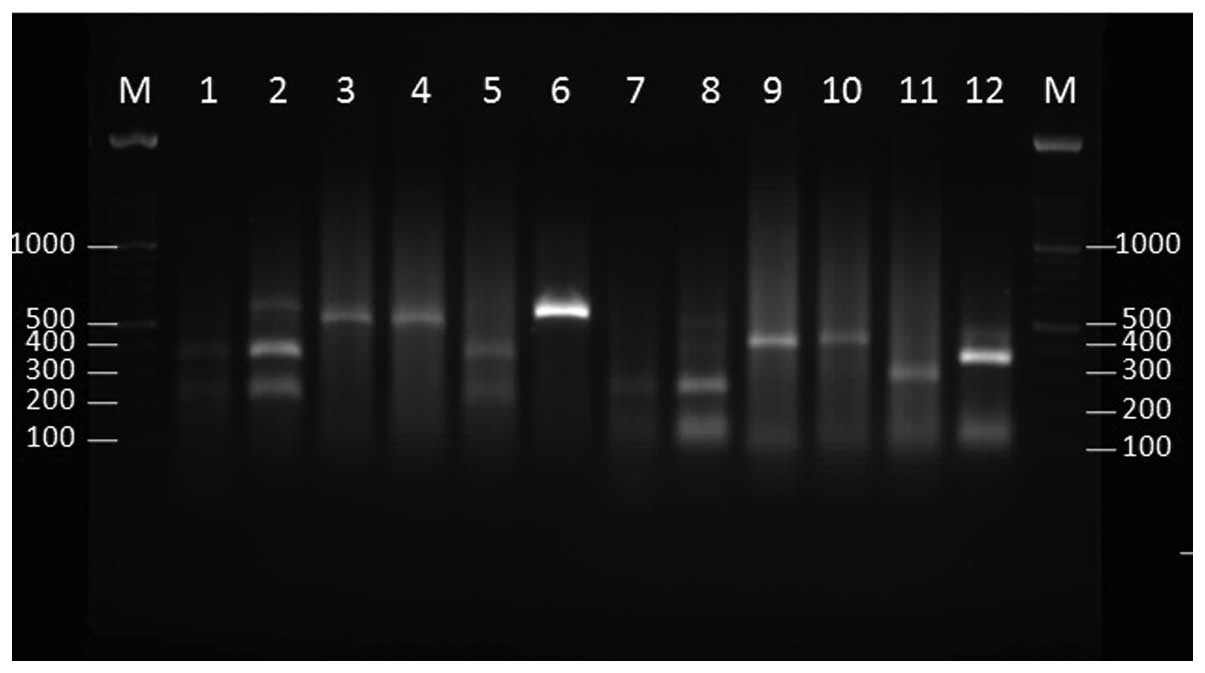 | Figure 3.Agarose gel electrophoresis of the
internal transcribed spacer region base pair (bp) products of the
Fusarium species (environmental isolates) following
digestion with SmaI (lanes 1–6) and HaeIII (lanes
7–12). M, 100 bp ladder; lane 1, E20; lane 2, E34; lane 3, E35;
lane 4, E36; lane 5, E39; lane 6, E42; lane 7, E20; lane 8, E34;
lane 9, E35; lane 10, E36; lane 11, E39; lane 12, E42. |
Discussion
The ITS region was confirmed in the present study to
be particularly suitable for the purpose of providing target genes
for molecular identification of the Fusarium species.
Variation in the nucleotide composition of the ITS region was
successfully employed for recognition among the species (11–13). A
variety of targets have been employed for DNA-based identification
and dicrimination of pathogenic Fusarium species. DNA
diversity in intergenic spacer (14)
or ITS regions (13), β-tubulin,
calmodulin, elongation factor 1α (15)
and mycotoxins biosynthetic genes (16) as target genes for identification of
Fusarium species have been tested with the PCR method.
Several studies have shown that ITS1 and ITS2 are useful targets
for detection of certain Fusarium species (13,17). The
study by O'Donnell (18) reported an
unexpected level of divergence for ITS sequences within the F.
sambucinum species. There are certain advantages for the ITS
region being a good target for detection reasons. The ITS region is
comparatively conserved within numerous fungal species. It is
present as multiple copies in the genome of fungi, and yields
adequate taxonomic resolution for the majority of fungi (19). Furthermore, there are a large number of
sequences from this locus in the GenBank, which enable the
comparison of the obtained sequences. Therefore, nucleotide
sequence heterogeneity in this region could be employed for
classification of the majority of pathogenic fungi (20).
In the present study, the ITS1 and ITS4 primers were
used to amplify the 5.8S rDNA gene. RFLP using HaeIII and
SmaI restriction enzymes provided a genus-specific assay for
the rapid identification of medically significant Fusarium
genus. According to the RFLP result, five species of
Fusarium, which were F. solani, F. oxysporum,
F. verticillidea, F. proliferatum and F.
fujikuri, were divided into five RFLP types when used with the
HaeIII and SmaI enzymes.
In conclusion, it appears that the present PCR-RFLP
method produces a restriction profile for the differentiation of
the most medically significant Fusarium species.
Acknowledgements
The present study was supported by the Health
Research Institute (Infectious and Tropical Diseases Research
Center, Ahvaz Jundishapur University of Medical Sciences, Ahvaz,
Iran) (grant no. 92118).
References
|
1
|
Kuruvilla TS and Dias M: Fusarium Solani:
A causative agent of skin and nail infections. Indian J Dermatol.
57:308–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Varon AG, Nouer SA, Barreiros G, Trope BM,
Magalhães F, Akiti T, Garnica M and Nucci M: Superficial skin
lesions positive for Fusarium are associated with subsequent
development of invasive fusariosis. J Infect. 68:85–89. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nucci M and Anaissie E: Fusarium
infections in immunocompromised patients. Clin Microbiol Rev.
20:695–704. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan R, Ng KP, Gan GG and Na SL:
Fusarium sp. infection in a patient with Acute Lymphoblastic
Leukaemia. Med J Malaysia. 68:479–480. 2013.PubMed/NCBI
|
|
5
|
Ma LJ, Geiser DM, Proctor RH, Rooney AP,
O'Donnell K, Trail F, Gardiner DM, Manners JM and Kazan K:
Fusarium pathogenomics. Annu Rev Microbiol. 67:399–416.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tamura M, Mochizuki N, Nagatomi Y,
Harayama K, Toriba A and Hayakawa K: A method for simultaneous
determination of 20 Fusarium toxins in cereals by
high-resolution liquid chromatography-Orbitrap mass spectrometry
with a pentafluorophenyl column. Toxins (Basel). 7:1664–1682. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barros G, Zanon MS, Palazzini JM,
Haidukowski M, Pascale M and Chulze S: Trichothecenes and
zearalenone production by Fusarium equiseti and Fusarium
semitectum species isolated from Argentinean soybean. Food
Addit Contam Part A Chem Anal Control Expo Risk Assess.
29:1436–1442. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arif M, Zaidi NW, Haq QM, Singh YP, Taj G,
Kar CS and Singh US: Morphological and comparative genomic analyses
of pathogenic and non-pathogenic Fusarium solani isolated
from Dalbergia sissoo. Mol Biol Rep. 42:1107–1122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Datta J and Lal N: Application of
molecular markers for genetic discrimination of Fusarium
wilt pathogen races affecting chickpea and pigeonpea in major
regions of India. Cell Mol Biol (Noisy-le-grand). 58:55–65.
2012.PubMed/NCBI
|
|
10
|
Short DP, O'Donnell K and Geiser DM:
Clonality, recombination, and hybridization in the
plumbing-inhabiting human pathogen Fusarium keratoplasticum
inferred from multilocus sequence typing. BMC Evol Biol. 14:912014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dubey SC, Priyanka K and Singh V:
Phylogenetic relationship between different race representative
populations of Fusarium oxysporum f. sp. ciceris in respect
of translation elongation factor-1α, β-tubulin, and internal
transcribed spacer region genes. Arch Microbiol. 196:445–452. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chang SC, Macêdo DP, Souza-Motta CM and
Oliveira NT: Use of molecular markers to compare Fusarium
verticillioides pathogenic strains isolated from plants and
humans. Genet Mol Res. 12:2863–2875. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mirete S, Patiño B, Jurado M, Vázquez C,
González-Jaén MT and Puertas M: Structural variation and dynamics
of the nuclear ribosomal intergenic spacer region in key members of
the Gibberella fujikuroi species complex. Genome.
56:205–213. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Konstantinova P and Yli-Mattila T:
IGS-RFLP analysis and development of molecular markers for
identification of Fusarium poae, Fusarium
langsethiae, Fusarium sporotrichioides and Fusarium
kyushuense. Int J Food Microbiol. 95:321–331. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Van Poucke K, Monbaliu S, Munaut F,
Heungens K, De Saeger S and Van Hove F: Genetic diversity and
mycotoxin production of Fusarium lactis species complex
isolates from sweet pepper. Int J Food Microbiol. 153:28–37. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Y, Bouras N, Yang J, Howard RJ and
Strelkov SE: Mycotoxin production by isolates of Fusarium
lactis from greenhouse sweet pepper (Capsicum annuum).
Int J Food Microbiol. 151:150–156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
O'Donnell K, Sutton DA, Fothergill A,
McCarthy D, Rinaldi MG, Brandt ME, Zhang N and Geiser DM: Molecular
phylogenetic diversity, multilocus haplotype nomenclature, and in
vitro antifungal resistance within the Fusarium solani
species complex. J Clin Microbiol. 46:2477–2490. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
O'Donnell K: Ribosomal DNA internal
transcribed spacers are highly divergent in the phytopathogenic
ascomycete Fusarium sambucinum (Gibberella
pulicaris). Curr Genet. 22:213–220. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Balajee SA, Borman AM, Brandt ME, Cano J,
Cuenca-Estrella M, Dannaoui E, Guarro J, Haase G, Kibbler CC, Meyer
W, et al: Sequence-based identification of Aspergillus,
Fusarium, and Mucorales species in the clinical
mycology laboratory: Where are we and where should we go from here?
J Clin Microbiol. 47:877–884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iwen PC, Hinrichs SH and Rupp ME:
Utilization of the internal transcribed spacer regions as molecular
targets to detect and identify human fungal pathogens. Med Mycol.
40:87–109. 2002. View Article : Google Scholar : PubMed/NCBI
|















