Introduction
Fibroblast growth factor 21 (FGF21) is a member of
the family of fibroblast growth factors (FGFs). Since the discovery
of the first FGF (FGF1 or FGF-a) in 1976 (1), 22 members of this family have been
identified (1). FGFs exhibit diverse
biological and physiological activities. Specifically, FGFs are
involved in metabolic regulation and cell differentiation,
proliferation and metabolism (1–6). FGF21 was
first cloned by Nishimura et al (7). In 2005, FGF21 was reported as a novel
metabolic regulator with biological activities (8). Subsequent studies revealed that FGF21 is
a key factor secreted by the liver and is a signaling molecule
involved in important metabolic regulation. Therefore, the current
study presents a summary of the molecular mechanisms underlying the
metabolic regulation of FGF21 in different types of tissue,
including liver, adipose and nervous system tissues. The findings
may serve as a reference for drug development of FGF21.
Biological actions of FGF 21
Kharitonenkov et al (8) first demonstrated the effects of FGF21 on
metabolic regulation and found that FGF21 regulates the expression
of glucose transporter 1, and promotes the intake of glucose in
3T3-L1 cells and primary lipocytes in the human body. Subsequent
experiments indicated that FGF21 significantly reduced the blood
sugar levels and body weights of mice with genetic control- or
diet-induced obesity by reducing the contents of triglycerides in
the liver and serum; however, the glucose intake of the mice
demonstrated no significant change (8–10). In
addition, FGF21 influences the insulin sensitivity of the mice. In
mice with diet-induced obesity, FGF21 may reverse hepatic steatosis
and enhance hepatic insulin sensitivity by suppressing glucose
production in the liver and increasing hepatic glycogen content,
thereby improving systemic glucose intolerance and insulin
resistance (10–12). The regulatory effects of FGF21 in
glycolipid metabolism are influenced by hepatic metabolism
(12); however, the specific
underlying mechanism remains to be elucidated. FGF21 was shown to
exert the same metabolic effects in monkeys with diabetes mellitus.
FGF21 reduced the concentration of low-density lipoprotein
cholesterol (LDL-C) and increased that of high-density lipoprotein
cholesterol (HDL-C) (13). These
findings indicate that FGF21 is significant in metabolic regulation
in rodents and non-human primates with obesity.
Receptors mediating the biological effects
of FGF21
FGF21 participates in signal transduction by
triggering FGF receptors (FGFRs) (8).
Unlike conventional FGFs, FGF21 does not directly bind FGFRs, but
exerts the biological functions by binding the accessory receptor
βKlotho and FGFRs. This conclusion has been confirmed by the
following: i) βKlotho restores ththe biological functions by
binding the accessory receptor βKlotho and FGFRs. This conclusion
has been confirmed by the following: i) βKlotho restores the
activity of FGF21 in cells in which FGF21 is ineffective (14–16); ii) the
expression of the induced receptor βKlotho increases susceptibility
to FGF21 (17); and iii)
downregulation of βKlotho mediated by small-interfering RNA weakens
the activity of FGF21. Therefore, the receptor of FGF21 consists of
the following two components in terms of structure: FGFR and
βKlotho (15). Neither βKlotho nor
FGFR activate the signaling of FGF21, and FGF21 only acts when the
two are combined.
The N- and C-termini of FGF21 are closely associated
with the bioactivity of FGF21; after the C-terminus combines with
the transmembrane protein of βKlotho, the N-terminus binds with
FGFR. Subsequent to forming a stable FGF21/βKlotho/FGFR complex,
the associated downstream signaling molecules are activated to
exert their biological effects (18,19).
Previous studies have confirmed that FGFR1c is a key receptor of
FGF21, and that other receptors mediating FGF21 signaling include
FGFR2 and FGFR4 (14,18). βKlotho is abundant in various types of
metabolically active tissue, including the liver, heart, white
adipose tissue (WAT), brown adipose tissue (BAT), and the central
nervous system (20). FGFRs are widely
expressed in various tissue and the selective expression of βKlotho
determines the tissue-specific biological functions of FGF21
(21).
Regulatory effects of FGF21 in various
tissues of the body
Effects of FGF21 in the liver
FGF21 is expressed in the liver, adipose tissue,
central nervous system, and other tissues (20). Under normal physiological us system,
and other tissues (20). Under normal
physiological conditions, FGF21 in the blood circulation is derived
from hepatic secretion (22). In the
liver, long-time fasting may induce the expression of FGF21,
although gain-of-function experimental results indicated that FGF21
itself also causes various effects of fasting; specifically, FGF21
stimulates the oxidation of fatty acids and the production of
ketone bodies in the liver and inhibits lipogenesis (9,23–25). In addition, loss-of-function
experimental results showed that mice with FGF21 knockout exhibit
serious weight gains after ketogenic diets, with hepatic steatosis
and functional impairments in the synthesis of ketone bodies, as
compared with wild-type mice (26).
Transcriptional mechanisms of FGF21 in
the liver
The complex network of nuclear receptors, nutritive
stimuli and hormones involves the gene regulation of FGF21 in
different tissues, thereby influencing its effects in regulating
hunger stimuli and systemic energy balance. Badman et al
(23) and Inagaki et al
(24) found that the nuclear receptor
peroxisome proliferator-activated receptor α (PPARα) induces FGF21
expression. This findiuli and systemic energy balance. Badman et
al (23) and Inagaki et al
(24) found that the nuclear receptor
peroxisome proliferator-activated receptor α (PPARα) induces FGF21
expression. This finding indicates that various effects of PPARα in
the liver are mediated by FGF21. PPARα induces FGF21 transcription
in mice after long-time fasting, ketogenic diets, and treatment of
primary hepatocytes treated with PPARα by combining peroxisome
proliferator response elements (PPRE) transcriptional response
elements on the FGF21 promoter (23,24).
However, in the liver of mice with PPARα knockout, hunger stimuli
induced a five-fold increase in the RNA level of FGF21 (24). This finding indicates that FGF21
transcription is regulated by additional cytokines. Furthermore,
the results confirmed that FGF21 is the direct target of the
retinoic acid receptor β (RARβ). RARβ overexpression increases the
generation and secretion of FGF21 in the liver, thereby performing
a regulatory role in metabolism (27).
The NAD+-dependent histone deacetylase, sirtuin 1
alleviates the fasting-induced hepatic fat deposition in mice by
inducing FGF21 expression in the liver (28,29). Recent
gain-of-function and loss-of-function experimental results
demonstrated that the transcription of FGF21 in primary hepatocytes
in the liver is also regulated by cAMP response element binding
protein (CREB) H (30), PPARγ
coactivator1α (31) and retinoic acid
receptor-related orphan receptor α (32). In addition, the carbohydrate response
element binding protein, activated by high-carbohydrate diets,
upregulates FGF21 in the liver (33).
FGF21 is regulated by the inositol-requiring enzyme 1α-X-box
binding protein 1 signaling pathway of the unfolded protein
response (34). Thus, FGF21 exerts a
wide range of effects on the hepatic response to nutritive stimuli
and regulates energy metabolism.
Regulatory mechanisms of FGF21 in
glycolipid metabolism in the liver
FGF21 exerts significant effects on hepatic
metabolism; specifically, it refines hepatic insulin resistance and
steatosis in mice with diet-induced obesity, and regulates glycogen
synthesis and ketone body production (9,23–25). In mice injected with FGF21
extracellular signal-regulated kinase (ERK)1/2 phosphorylation is
induced in the liver and immediately regulates the expression of
early genes in the liver. FGF21 also induces the expression of
peroxisome proliferator-activated receptor gamma coactivator 1α and
its downstream genes, and enhances the oxidation of fatty acids in
the liver (25). However, FGF21
exerted no effect in the liver of isolated mice or in the primary
hepatocytes of rats, which indicates that the effects of FGF21 in
the liver are indirectly mediated by other signaling pathways of
the liver or by other tissues, such as adipose tissues (35). Lin et al (35) found that adiponectin (APN) mediates the
regulatory effects of FGF21 on systemic metabolism. The authors
also found that the capacity of FGF21 to ameliorate plasma
triglycerides, hepatic steatosis, and liver injuries disappears in
mice with obesity and diet-induced APN knockout (35). However, FGF21 may still significantly
reduce the blood sugar level of mice with APN knockout. This result
demonstrates that APN mediates the lipid-lowering effects of FGF21,
although the specific glucose-lowering mechanisms of FGF21 remain
unidentified. In addition, FGF21 and tissue-specific accessory
receptor, βKlotho are highly expressed in the liver (33). This indicates that FGF21 directly
regulates hepatic metabolism using βKlotho/FGFR. Thus, mice with
liver-specific βKlotho knockout or silencing are used in
experiments to elucidate the direct effect of FGF21 on the liver
and its relative contribution to the balance of systemic glycolipid
metabolism. Various studies have evaluated the upstream regulation
of FGF21; however, the downstream regulation mediated by FGF21 in
the liver remains to be elucidated. Therefore, this is currently a
research hotspot.
Effects of FGF21 in adipose
tissues
In vivo and in vitro experiments on
the metabolic regulation of FGF21 revealed that either the
overexpressed FGF21 transgene or the exogenous FGF21 significantly
reduce glucose and lipid levels in the blood of mice (8,10). However,
FGF21 increased the intake of glucose in mature 3T3-L1 cells and
the primary lipocytes in the human body (36). Subsequent experiments revealed that
FGF21 regulates the intake of glucose in WAT, the generation and
decomposition of adipose tissues, and lipid metabolism by mediating
the expression of associated genes (8,9,36). FGF21 induces the expression of
uncoupling protein 1 (UCP1) in WAT (37). UCP1 is a typical protein in BAT that is
involved in uncoupling respiration and thermogenesis. This
phenomenon is the so-called ‘browning’ of WAT, usually occurring in
subcutaneous WAT dominated by the sympathetic nerve; FGF21 is able
to regulate thermogenesis using this mechanism (37). In addition, FGF21 activates the
expression and secretion of APN in lipocytes by autocrine or
paracrine signaling (35,38), thereby regulating the glycolipid
metabolism in adipose tissues and further improving insulin
sensitivity. In vivo and in vitro experiment results
also showed that FGF21 stimulates the expression of genes
associated with glucose intake and thermogenesis in BAT, thereby
regulating energy metabolism (10,37,39,40).
Transcriptional regulation mechanism
of FGF21 in adipose tissues
In WAT, the nuclear receptor, PPARγ induces the
expression of FGF21 (41). FGF21 acts
in synergy with the PPARγ stimulant, rosiglitazone, thereby
promoting the differentiation of lipocytes and the intake of
glucose (17,41–43). Unlike
the hunger-induced regulation of FGF21 in the liver, the
feeding-induced regulation of FGF21 in WAT further activates the
activity of PPARγ in the form of feedforward regulation (44). The PPARγ signaling pathway is impaired
in mice with FGF21 knockout through the downregulation of genes
depending on body fat and PPARγ expression. In addition, the PPARγ
stimulant, rosiglitazone has side effects, including insulin
sensitization, weight gain and edema; however, such side effects do
not occur in mice with FGF21 knockout. These results demonstrate
that FGF21 is a major carrier of the physiological and
pharmacological effects of PPARγ in WAT (44). In addition, FGF21 is induced by cold
stimuli in BAT, and it regulates thermogenesis via activating
transcription factor 2 (ATF2)-mediated β3-adrenaline receptor
(37,39,45). FGF21
also directly regulates UCP1 transcription by increasing the
transcription factor AMP response elements and activating the
phosphorylation of CREB (46). FGF21
also increases the phosphorylation of fatty acid transport protein
3, thereby regulating mitochondrial respiration (46). In a cold stimulation experiment, FGF21
knockout mice exhibited low body temperatures, elevated serum
creatinine kinase levels and trembling (37). Thus, FGF21 regulates energy metabolism
by promoting thermogenesis in BAT.
Regulatory mechanism of FGF21 in the
glycolipid metabolism and energy metabolism in adipose tissues
In vivo loss-of-function experiment results
confirmed the significant effects of FGF21 in adipose tissues.
After the specific receptor βKlotho of FGF21 is knockout from
adipose tissues, the acute effects of FGF21 on insulin sensitivity
and glucose intake disappeared from the adipose tissues of the mice
with the gene knockout compared with those of the control mice fed
with high fat diets (40). Similarly,
in diet-induced obesity mice with selective deletion of FGFR1c in
adipose tissues, FGF21 exerts no effects on the weights or on the
levels of plasma glucose, insulin, triglycerides, or APN (47). Thus, βKlotho and FGFR mediate the
effects of FGF21 in regulating the glycolipid metabolism in adipose
tissues. The browning and thermogenesis of WAT reveal the important
regulatory effect of FGF21 on the energy metabolism and body weight
of the mice. Furthermore, the signal transmission and cross-talk
mechanisms between the liver and adipose tissues mediate the
regulatory effects of FGF21 on energy metabolism. The endocrine
signaling pathway of FGF21 in hepatocytes increases the energy
expenditure in mice by regulating the browning of WAT, thereby
enhancing the energy metabolism and reducing body weight (28). Additionally, Fisher et al
(37) found that the in vivo
and in vitro thermogenesis of FGF21 is closely associated
with peroxisome proliferator-activated receptor gamma coactivator
1α (PGC1α), the key factor regulating thermogenesis and energy
metabolism (37). In addition, FGF21
regulates the browning of WAT by elevating the level of protein
rather than gene expression of PGC1α, thereby promoting
thermogenesis (37).
Effects of FGF21 in the brain
Although it is not expressed in the brain (20), FGF21 passes through the blood brain
barrier (BBB) by simple diffusion, thereby entering the human
cerebrospinal fluid (48) and the
hypothalami of fasting mice (49), and
further activating ERK1/2 phosphorylation (50). Administering FGF21 in the cerebral
ventricles increases energy expenditure and improves the insulin
sensitivity in obese rats (51),
thereby triggering the FGF21-mediated signal interaction between
the brain and its peripheral tissues (52). Thus, FGF21 directly acts on the
brain.
Regulatory mechanisms of FGF21 in the
brain
In the hypothalamus, FGF21 affects the expression
levels of two important neuropeptides, namely, corticotrophin
releasing factor (CRF) and arginine vasopressin (AVP) (49,53–55) in the brain. FGF21 induces CRF
expression in the hypothalamic (49,53,55). Consequently, CRF increases the
secretion of CRF in the pituitary gland, thereby stimulating the
secretion of CRF. CRF-induced sympathetic excitation also passes
onto BAT (56). Notably, CRF receptor
inhibitors completely block FGF21-induced sympathetic excitation
(53). Therefore, its inducing action
on CRF is deemed as a mechanism by which FGF21 promotes
glucocorticoids, thermogenesis and weight loss. Conversely,
long-term treatment of lean mice (except for those with CRF-induced
weight loss) with FGF21 may also inhibit AVP expression in the
suprachiasmatic nucleus (SCN) (54).
In the hypothalami of hungry mice with βKlotho knockout, the mRNA
expression level of AVP is elevated. In female mice, the
downregulation of AVP may decrease the level of luteinizing hormone
secreted in the pituitary gland and reduce ovulation, thereby
inhibiting reproduction.
However, the specific site of the CRF-induced action
of FGF21 in the hypothalamus remains unknown. A recent study
demonstrated that βKlotho is expressed in the CRF neurons of the
paraventricular nucleus (PVN) (49).
At this site, FGF21 induces CRF expression by activating the
transcription factor, CREB (49). This
result is consistent with the expression of UCP1 in FGF21-induced
BAT (46). Therefore, FGF21 in the PVN
also directly or indirectly induces the actions of CRF.
Effects of FGF21 in signal
transmission and cross-talk in the brain and its peripheral
tissues
Four types of FGFRs extensively exist in the nervous
system; however, the expression of βKlotho in the nervous system is
limited (20). In the hypothalamus,
βKlotho is expressed in the SCN and PVN (49,55), in the
nucleus tractus solitarius of the back polar region and hindbrain,
and in the nodose ganglion root of the peripherals (55). These regions constitute the dorsal
vagal complex, which is a major integrated aspect of the autonomic
nervous system.
The brain mediates certain roles of FGF21 in the
systemic blood circulation system. In mice, various chronic effects
of FGF21 on the production of ketone bodies, circadian rhythms, and
the fertility of female mice require the involvement of the βKlotho
protein in the central nervous system (54,55). Further
research confirmed that FGF21 stimulates the release of CRF from
the nervous system to activate the sympathetic nervous system,
thereby causing WAT browning, fatty acid oxidation in BAT,
thermogenesis and lipolysis accompanying hepatic ketogenesis
(53). These results demonstrate why
certain effects of FGF21 disappear in the absence of βKlotho from
the nervous system of mice (53,55).
Notably, the glucose-lowering roles of the FGF21 transgene are
intact in mice with brain-specific βKlotho knockout (55), but are completely inhibited in mice
with βKlotho systemic knockout (40,57). This
result reveals that other metabolic tissues mediate the
glucose-lowering roles of FGF21.
Effects of FGF21 in other tissues
FGF21 is also expressed in the endocrine-specific
pancreatic α and β cells, and the expression of FGF21 is exposed to
chemical-induced pancreatic trauma (58). However, the physiological functions of
FGF21 in chemical-induced pancreatic trauma remain unknown
(58). In isolated islet cells, FGF21
increases the quantity of insulin and the secretion of insulin
induced by glucose stimulation, thereby inhibiting glucagon
secretion (8) and exerting protective
effects on mice with pancreatitis (58). Furthermore, studies have reported that
FGF21 modulates inflammation and damage induced by experimental
pancreatitis. FGF21-null mice develop more damage than wild-type
mice, whereas mice overexpressing human FGF21 exhibit an attenuated
phenotype (58). Further studies
identified the transcription factor, basic helix-loop-helix family
member a15 (BHLHA15; also termed, MIST1) as an upstream regulator
of FGF21 and showed that deletion of the MIST1 gene leads to a
marked reduction in pancreatic FGF21 levels by epigenetic
silencing, resulting in increased susceptibility to pancreatitis
(59). In addition, FGF21 may be
involved in enhanced islet transplant survival, which was
demonstrated in a model of streptozotocin-induced diabetes
(60). FGF21 may promote β-cell
survival, and protect isolated rat islets and insulin-producing INS
cells from glucolipotoxicity and cytokine-induced apoptosis
(61). However, human FGF21 fails to
alter insulin and glucagon secretion from islets isolated from
healthy mice (62), although FGF21
stimulates insulin secretion in ex vivo islets isolated from
diabetic mice (61). Islets from obese
diabetic db/db mouse failed to respond to FGF21, possibly as a
consequence of reduced βKlotho expression levels (63).
In addition, the dysfunction of the mitochondria in
the skeletal muscle induces the expression of FGF21 by activating
ATF4 (64,65). Thus, FGF21 is important in the
metabolic network. Fig. 1 briefly
outlines the metabolic regulations of FGF21 in different tissues
and the interactions between them.
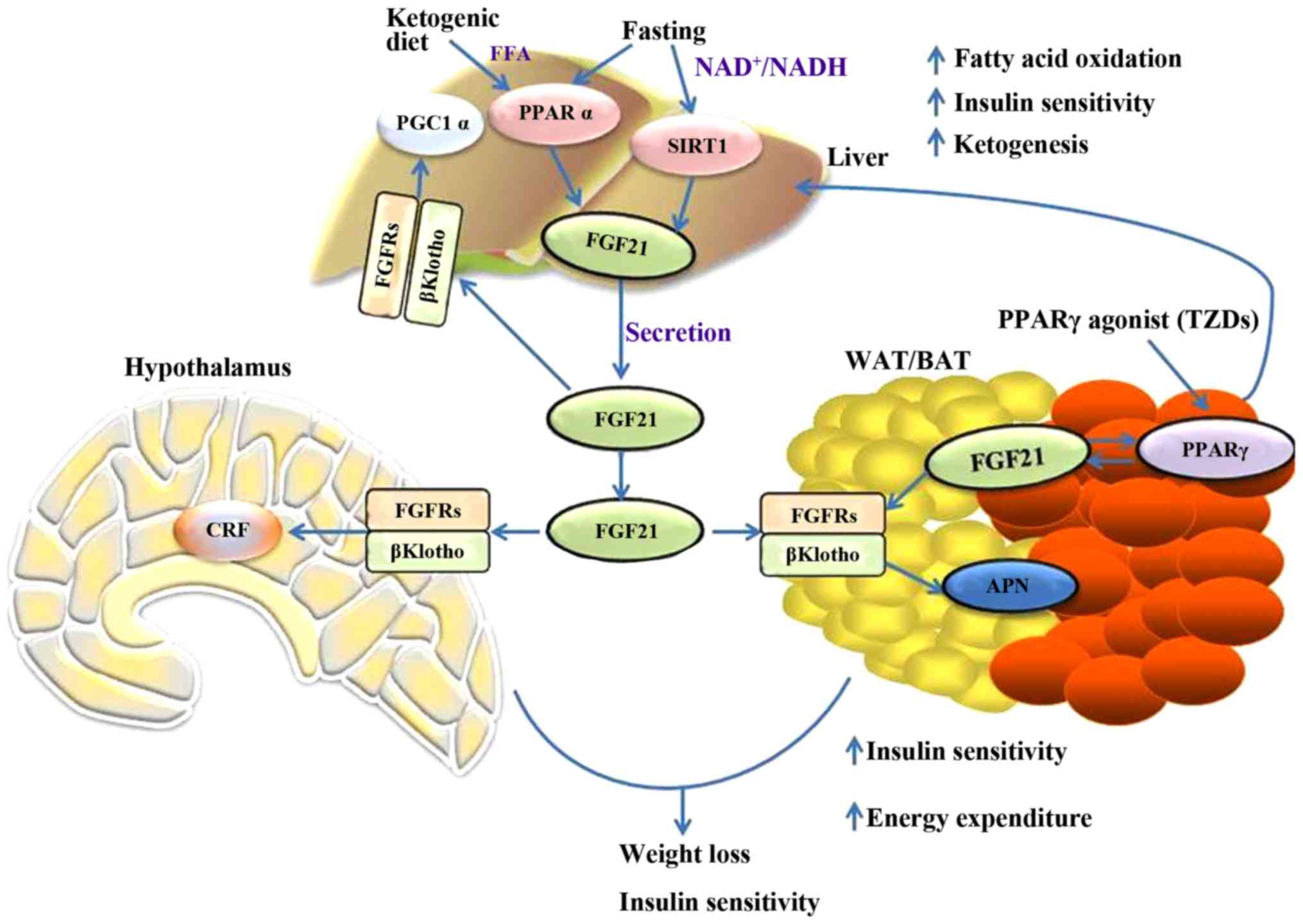 | Figure 1.Summary of the metabolic actions of
FGF21 in the liver, adipose, and nervous system tissues. FGF21,
fibroblast growth factor 21; PGC1α, peroxisome
proliferator-activated receptor gamma coactivator 1α; PPARα,
peroxisome proliferator-activated receptor α; SIRT1, sirtuin 1;
FGFR, fibroblast growth factor receptor; APN, adiponectin; CRF,
corticotrophin releasing factor; WAT, white adipose tissue; BAT,
brown adipose tissue; TZD, thiazolidinediones. |
Clinical applications of FGF21
Manifestations of FGF21 in the human
body
Similar to the findings in experiments with mice,
PPARα stimulants (such as fibrates) increase the circulating level
of FGF21 in the human body (66–68);
however, the concentration of FGF21 is significantly increased in
patients with rheumatoid arthritis after 7 days of fasting or in
patients with obesity and diets low in calories within 3 weeks
(66). These results demonstrate that
FGF21 performs similar metabolic regulation and clinical
applications in different species; however, the effects of PPARα
stimulant and fasting on FGF21 expression are relatively weaker in
the human body than in mice. Notably, neither short duration
fasting nor anorexia nervosa increases the circulating level of
FGF21 in the human body (68).
Ketogenic diets do not elevate FGF21 expression levels in the human
body (69). This discrepancy may be
caused by the low content of protein in the ketogenic diet used in
the experiment with mice (70).
Therefore, the regulatory roles of FGF21 in rodents differ from
those in the human body. Notably, certain studies detected FGF21
mRNA expression in human WAT (43,71),
although Dushay et al (69)
reported that FGF21 is not expressed in human WAT. In addition,
clinical studies revealed that the circulating levels of FGF21 in
patients with obesity or in those with accompanying diabetes
mellitus (type 2), insulin resistance, or nonalcoholic fatty liver
disease are considerably elevated (71–73). This
result may be attributed to the increased content of FGF21 in the
liver induced by diets containing high levels of fat and
carbohydrates. Similarly, the concentration of FGF21 is increased
in the plasma of rodents with obesity, and this phenomenon is
similar to hyperinsulinism and hyperleptinemia; therefore,
introducing the concept of FGF21 resistance (11,74,75). However, when the concentration of FGF21
in the blood of mice with diet-induced obesity is increased, FGF21
effectively exerts pharmacodynamic effects, with a marked increase
in insulin sensitivity. Therefore, the hypothesis on FGF21
resistance remains controversial (74,76).
FGF21 significantly reduced body weight and improved
insulin sensitivity in obese rodents and primates (76). Prolonged fasting or feeding with a
ketogenic diet results in hepatic production and secretion of
FGF21. In the liver, FGF21 increases fatty acid oxidation and
ketogenesis, and decreases hepatic lipid biosynthesis, resulting in
improved insulin sensitivity (75).
FGF21 induces production and secretion of adiponectin through PPARγ
in adipose tissue, which in turn potentiates insulin-sensitizing
effects of PPARγ and stimulates browning by inducing PGC1α
(75,76). FGF21 stimulates sympathetic nerve
activity via CRF, leading to improved whole body metabolism. These
effects of FGF21 require signaling via the liver- and
adipose-enriched coreceptor, βKlotho and one of the FGFRs, such as
FGFR1, FGFR2 or FGFR4.
Clinical trials on FGF21
The significant curative effects of FGF21 in the
animal model are of interest. However, inadequacies in its physical
and biological properties rendered the natural FGF21 protein hard
to apply to molecule drugs for clinical settings (77). The main causes include the following:
First, the FGF21 protein stored in solution is easily degraded. In
addition, FGF21 is conformationally unstable. In addition, the
large-scale expression and purification of FGF21 are difficult.
Considering the above factors, Kharitonenkov et al (78) developed a drug, LY2405319 (LY), which
avoids the biopharmaceutical defects of FGF21; the engineered drug
is an FGF21 isomer that retains the curative effects of FGF21 on
various metabolic disorders, but avoids the defects of the
wild-type FGF21 (78). For patients
with obesity and type 2 diabetes mellitus, LY is administered by
daily injection, and clinical test results indicated that injected
LY significantly improves the profiling levels of four types of
lipids (i.e., total cholesterol, LDL-C, HDL-C and triglycerides)
and normalizes the body weights of these patients. These results
are consistent with those of FGF21 or LY in Rhesus monkeys with
obesity (13).
The triglyceride level declines quickly after
fasting and LY treatment. This phenomenon is accompanied by a
reduction in apolipoprotein C III and aesult indicates that LY
regulates the oxidation level of fatty acids in the human body
(26). Furthermore, the insulin level
in the human body subsequent to fasting and LY treatment is reduced
markedly, the APNesult indicates that LY regulates the oxidation
level of fatty acids in the human body (79–81).
Furthermore, the insulin level in the human body subsequent to
fasting and LY treatment is reduced markedly, the APN level in the
plasma is elevated appreciably (79),
and the APN of high molecular weight is associated with the
treatment efficiency of type 2 diabetes mellitus. However, the
continuous glucose-lowering effect of LY on the subject is
insignificant compared with that of FGF21 in rodents and monkeys
with diabetes mellitus, which is potentially due to individual
differences and small sample sizes (8,13,82). The potential effect and glucose
regulation of exogenous FGF21 in obese patients may be elucidated
further using a large sample size, employing long-term regimens, or
treating patients with other diseases, such as impaired fasting
glucose or type 2 diabetes mellitus of various severities (79).
Conclusion
The current review primarily summarizes the
metabolic functions and regulatory mechanisms of FGF21 in the
liver, adipose, brain and other tissues. Given its consistent and
reproducible efficacy in animals, FGF21 is a candidate drug for
curing obesity and diabetes mellitus. This drug is now entering the
pre-clinical research stage. Clinical trials with a large sample
size are required to validate the potential therapeutic effects of
FGF21 on metabolic diseases. Although wild-type FGF21 may have side
effects on bone mass and fertility, population studies showed that
the content of FGF21 in the blood of healthy women positively
correlates with bone density (61).
This result suggests that the effect of FGF21 in mice with
osteoporosis may differ from that in human bones. However, the
inhibitory actions of FGF21 on fertility via the central nervous
system are eliminated by synthesizing FGF21 isomers, which could
not pass through the BBB, by genetic modification (61).
To date, the mechanism of action of FGF21 has not
been fully elucidated, and studies on the biological target organs
and molecular mechanisms of FGF21 remain incomplete. There are few
studies discussing whether FGF21 as a secretory hormone regulates
systemic metabolism in different tissues. In addition,
thiazolidinedione drugs affect the expression of FGF21 and its
receptor βKlotho (17,41,42), as well
as enhancing the sensitivity of FGF21 (17). This result indicates that these drugs
act against diabetes mellitus by partially affecting the FGF21
signaling pathway. Therefore, intensive studies on the signaling
pathway, regulatory mechanisms and biological functions of FGF21 in
different tissues are significant for the treatment of
human-associated metabolic disorders, such as diabetes mellitus and
obesity. With the number of patients with metabolic syndrome
increasing annually, the development of drugs for this disease has
been receiving considerable attention (61). With further animal experiments and
clinical trials covering the improvements in the function and
survival of β cells, FGF21 is expected to become a novel candidate
for insulin in treating diabetes mellitus. However, animal
experiments are inconsistent with clinical trials in certain
aspects, and FGF21 requires further investigation. The elevated
expression level of FGF21 in the plasma indicates that FGF21
resistance probably compensates for glucolipotoxicity and requires
an FGF21 dose exceeding the normal to exert the necessary
biological effects. Given the elevated level of FGF21 in the plasma
of patients with metabolic disorders, whether FGF21 will be
developed into a molecular marker for the early diagnosis and
assessment of associated diseases requires further
investigation.
Acknowledgements
The present study was supported by the Natural
Science Foundation of China (grant no. 81600342), the Medical
Foundation of Hui Zhou (grant no. 2015Y134), the Medical Research
Foundation of Guangdong Province (grant no. A2015620) and the
Graduate Student Research Innovation Project of Hunan Province
(grant no. CX2013B396).
References
|
1
|
Itoh N: Hormone-like (endocrine) Fgfs:
Their evolutionary history and roles in development, metabolism,
and disease. Cell Tissue Res. 342:1–11. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kelleher FC, O'Sullivan H, Smyth E, Mc
Dermott R and Viterbo A: Fibroblast growth factor receptors,
developmental corruption and malignant disease. Carcinogenesis.
34:2198–2205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Evans SJ, Choudary PV, Neal CR, Li JZ,
Vawter MP, Tomita H, Lopez JF, Thompson RC, Meng F, Stead JD, et
al: Dysregulation of the fibroblast growth factor system in major
depression. Proc Natl Acad Sci USA. 101:15506–15511. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hutley L, Shurety W, Newell F, Mc Geary R,
Pelton N, Grant J, Herington A, Cameron D, Whitehead J and Prins J:
Fibroblast growth factor 1: A key regulator of human adipogenesis.
Diabetes. 53:3097–3106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamagata H, Chen Y, Akatsu H, Kamino K,
Ito J, Yokoyama S, Yamamoto T, Kosaka K, Miki T and Kondo I:
Promoter polymorphism in fibroblast growth factor 1 gene increases
risk of definite Alzheimer's disease. Biochem Biophys Res Commun.
321:320–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
van der Walt JM, Noureddine MA, Kittappa
R, Hauser MA, Scott WK, McKay R, Zhang F, Stajich JM, Fujiwara K,
Scott BL, et al: Fibroblast growth factor 20 polymorphisms and
haplotypes strongly influence risk of Parkinson disease. Am J Hum
Genet. 74:1121–1127. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nishimura T, Nakatake Y, Konishi M and
Itoh N: Identification of a novel FGF, FGF-21, preferentially
expressed in the liver. Biochim Biophys Acta. 1492:203–206. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kharitonenkov A, Shiyanova TL, Koester A,
Ford AM, Micanovic R, Galbreath EJ, Sandusky GE, Hammond LJ, Moyers
JS, Owens RA, et al: FGF-21 as a novel metabolic regulator. J Clin
Invest. 115:1627–1635. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Coskun T, Bina HA, Schneider MA, Dunbar
JD, Hu CC, Chen Y, Moller DE and Kharitonenkov A: Fibroblast growth
factor 21 corrects obesity in mice. Endocrinology. 149:6018–6027.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu J, Lloyd DJ, Hale C, Stanislaus S, Chen
M, Sivits G, Vonderfecht S, Hecht R, Li YS, Lindberg RA, et al:
Fibroblast growth factor 21 reverses hepatic steatosis, increases
energy expenditure, and improves insulin sensitivity in dietinduced
obese mice. Diabetes. 58:250–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Berglund ED, Li CY, Bina HA, Lynes SE,
Michael MD, Shanafelt AB, Kharitonenkov A and Wasserman DH:
Fibroblast growth factor 21 controls glycemia via regulation of
hepatic glucose flux and insulin sensitivity. Endocrinology.
150:4084–4093. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu J, Stanislaus S, Chinookoswong N, Lau
YY, Hager T, Patel J, Ge H, Weiszmann J, Lu SC, Graham M, et al:
Acute glucose-lowering and insulin-sensitizing action of FGF21 in
insulin-resistant mouse models-association with liver and adipose
tissue effects. Am J Physiol Endocrinol Metab. 297:E1105–E1114.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kharitonenkov A, Wroblewski VJ, Koester A,
Chen YF, Clutinger CK, Tigno XT, Hansen BC, Shanafelt AB and Etgen
GJ: The metabolic state of diabetic monkeys is regulated by
fibroblast growth factor-21. Endocrinology. 148:774–781. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ogawa Y, Kurosu H, Yamamoto M, Nandi A,
Rosenblatt KP, Goetz R, Eliseenkova AV, Mohammadi M and Kuro-o M:
BetaKlotho is required for metabolic activity of fibroblast growth
factor 21. Proc Natl Acad Sci USA. 104:7432–7437. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kharitonenkov A, Dunbar JD, Bina HA,
Bright S, Moyers JS, Zhang C, Ding L, Micanovic R, Mehrbod SF,
Knierman MD, et al: FGF-21/FGF-21 receptor interaction and
activation is determined by betaKlotho. J Cell Physiol. 215:1–7.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki M, Uehara Y, Motomura-Matsuzaka K,
Oki J, Koyama Y, Kimura M, Asada M, Komi-Kuramochi A, Oka S and
Imamura T: betaKlotho is required for fibroblast growth factor
(FGF) 21 signaling through FGF receptor (FGFR) 1c and FGFR3c. Mol
Endocrinol. 22:1006–1014. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Moyers JS, Shiyanova TL, Mehrbod F, Dunbar
JD, Noblitt TW, Otto KA, Reifel-Miller A and Kharitonenkov A:
Molecular determinants of FGF-21 activitysynergy and cross-talk
with PPARgamma signaling. J Cell Physiol. 210:1–6. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yie J, Hecht R, Patel J, Stevens J, Wang
W, Hawkins N, Steavenson S, Smith S, Winters D, Fisher S, et al:
FGF21 N- and C-termini play different roles in receptor interaction
and activation. FEBS Lett. 583:19–24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Micanovic R, Raches DW, Dunbar JD, Driver
DA, Bina HA, Dickinson CD and Kharitonenkov A: Different roles of
N- and C- termini in the functional activity of FGF21. J Cell
Physiol. 219:227–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tacer Fon K, Bookout AL, Ding X, Kurosu H,
John GB, Wang L, Goetz R, Mohammadi M, Kuro-o M, Mangelsdorf DJ and
Kliewer SA: Research resource: Comprehensive expression atlas of
the fibroblast growth factor system in adult mouse. Mol Endocrinol.
24:2050–2064. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adams AC, Coskun T, Rovira AR, Schneider
MA, Raches DW, Micanovic R, Bina HA, Dunbar JD and Kharitonenkov A:
Fundamentals of FGF19 & FGF21 action in vitro and in vivo. PLo
S One. 7:e384382012. View Article : Google Scholar
|
|
22
|
Markan KR, Naber MC, Ameka MK, Anderegg
MD, Mangelsdorf DJ, Kliewer SA, Mohammadi M and Potthoff MJ:
Circulating FGF21 is liver derived and enhances glucose uptake
during refeeding and overfeeding. Diabetes. 63:4057–4063. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Badman MK, Pissios P, Kennedy AR, Koukos
G, Flier JS and Maratos-Flier E: Hepatic fibroblast growth factor
21 is regulated by PPARalpha and is a key mediator of hepatic lipid
metabolism in ketotic states. Cell Metab. 5:426–437. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Inagaki T, Dutchak P, Zhao G, Ding X,
Gautron L, Parameswara V, Li Y, Goetz R, Mohammadi M, Esser V, et
al: Endocrine regulation of the fasting response by
PPARalpha-mediated induction of fibroblast growth factor 21. Cell
Metab. 5:415–425. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Potthoff MJ, Inagaki T, Satapati S, Ding
X, He T, Goetz R, Mohammadi M, Finck BN, Mangelsdorf DJ, Kliewer SA
and Burgess SC: FGF21 induces PGC-1alpha and regulates carbohydrate
and fatty acid metabolism during the adaptive starvation response.
Proc Natl Acad Sci USA. 106:10853–10858. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Badman MK, Koester A, Flier JS,
Kharitonenkov A and Maratos-Flier E: Fibroblast growth factor
21-deficient mice demonstrate impaired adaptation to ketosis.
Endocrinology. 150:4931–4940. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Wong K, Walsh K, Gao B and Zang M:
Retinoic acid receptor β stimulates hepatic induction of fibroblast
growth factor 21 to promote fatty acid oxidation and control
whole-body energy homeostasis in mice. J Biol Chem.
288:10490–10504. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Y, Wong K, Giles A, Jiang J, Lee JW,
Adams AC, Kharitonenkov A, Yang Q, Gao B, Guarente L and Zang M:
Hepatic SIRT1 attenuates hepatic steatosis and controls energy
balance in mice by inducing fibroblast growth factor 21.
Gastroenterology. 146:539–549.e7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Purushotham A, Schug TT, Xu Q, Surapureddi
S, Guo X and Li X: Hepatocyte-specific deletion of SIRT1 alters
fatty acid metabolism and results in hepatic steatosis and
inflammation. Cell Metab. 9:327–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim H, Mendez R, Zheng Z, Chang L, Cai J,
Zhang R and Zhang K: Liver-enriched transcription factor CREBH
interacts with peroxisome proliferator-activated receptor α to
regulate metabolic hormone FGF21. Endocrinology. 155:769–782. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Estall JL, Ruas JL, Choi CS, Laznik D,
Badman M, Flier Maratos E, Shulman GI and Spiegelman BM: PGC-1alpha
negatively regulates hepatic FGF21 expression by modulating the
heme/Rev-Erb (alpha) axis. Proc Natl Acad Sci USA. 106:22510–22515.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Y, Solt LA and Burris TP: Regulation
of FGF21 expression and secretion by retinoic acid receptor-related
orphan receptor alpha. J Biol Chem. 285:15668–15673. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iizuka K, Takeda J and Horikawa Y: Glucose
induces FGF21 mRNA expression through ChREBP activation in rat
hepatocytes. FEBS Lett. 583:2882–2886. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shao M, Shan B, Liu Y, Deng Y, Yan C, Wu
Y, Mao T, Qiu Y, Zhou Y, Jiang S, et al: Hepatic IRE1α regulates
fasting-induced metabolic adaptive programs through the
XBP1s-PPARalpha axis signalling. Nat Commun. 5:35282014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin Z, Tian H, Lam KS, Lin S, Hoo RC,
Konishi M, Itoh N, Wang Y, Bornstein SR, Xu A and Li X: Adiponectin
mediates the metabolic effects of FGF21 on glucose homeostasis and
insulin sensitivity in mice. Cell Metab. 17:779–789. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kurosu H, Choi M, Ogawa Y, Dickson AS,
Goetz R, Eliseenkova AV, Mohammadi M, Rosenblatt KP, Kliewer SA and
Kuro-o M: Tissue-specific expression of betaKlotho and fibroblast
growth factor (FGF) receptor isoforms determines metabolic activity
of FGF19 and FGF21. J Biol Chem. 282:26687–26695. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fisher FM, Kleiner S, Douris N, Fox EC,
Mepani RJ, Verdeguer F, Wu J, Kharitonenkov A, Flier JS,
Maratos-Flier E and Spiegelman BM: FGF21 regulates PGC-1α and
browning of white adipose tissues in adaptive thermogenesis. Genes
Dev. 26:271–281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Holland WL, Adams AC, Brozinick JT, Bui
HH, Miyauchi Y, Kusminski CM, Bauer SM, Wade M, Singhal E, Cheng
CC, et al: An FGF21-adiponectin-ceramide axis controls energy
expenditure and insulin action in mice. Cell Metab. 17:790–797.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hondares E, Iglesias R, Giralt A, Gonzalez
FJ, Giralt M, Mampel T and Villarroya F: Thermogenic activation
induces FGF21 expression and release in brown adipose tissue. J
Biol Chem. 286:12983–12990. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ding X, Boney-Montoya J, Owen BM, Bookout
AL, Coate KC, Mangelsdorf DJ and Kliewer SA: βKlotho is required
for fibroblast growth factor 21 effects on growth and metabolism.
Cell Metab. 16:387–393. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muise ES, Azzolina B, Kuo DW, El-Sherbeini
M, Tan Y, Yuan X, Mu J, Thompson JR, Berger JP and Wong KK: Adipose
fibroblast growth factor 21 is up-regulated by peroxisome
proliferator-activated receptor gamma and altered metabolic states.
Mol Pharmacol. 74:403–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang H, Qiang L and Farmer SR:
Identification of a domain within peroxisome proliferator-activated
receptor gamma regulating expression of a group of genes containing
fibroblast growth factor 21 that are selectively repressed by SIRT1
in adipocytes. Mol Cell Biol. 28:188–200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang X, Yeung DC, Karpisek M, Stejskal D,
Zhou ZG, Liu F, Wong RL, Chow WS, Tso AW, Lam KS and Xu A: Serum
FGF21 levels are increased in obesity and are independently
associated with the metabolic syndrome in humans. Diabetes.
57:1246–1253. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dutchak PA, Katafuchi T, Bookout AL, Choi
JH, Yu RT, Mangelsdorf DJ and Kliewer SA: Fibroblast growth
factor-21 regulate PPARγ activity and the antidiabetic actions of
thiazolidinediones. Cell. 148:556–567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chartoumpekis DV, Habeos IG, Ziros PG,
Psyrogiannis AI, Kyriazopoulou VE and Papavassiliou AG: Brown
adipose tissue responds to cold and adrenergic stimulation by
induction of FGF21. Mol Med. 17:736–740. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu AL, Kolumam G, Stawicki S, Chen Y, Li
J, Zavala-Solorio J, Phamluong K, Feng B, Li L, Marsters S, et al:
Amelioration of type 2 diabetes by antibody-mediated activation of
fibroblast growth factor receptor 1. Sci Transl Med.
3:113ra1262011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Adams AC, Yang C, Coskun T, Cheng CC,
Gimeno RE, Luo Y and Kharitonenkov A: The breadth of FGF21's
metabolic actions are governed by FGFR1 in adipose tissue. Mol
Metab. 2:31–37. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tan BK, Hallschmid M, Adya R, Kern W,
Lehnert H and Randeva HS: Fibroblast growth factor 21 (FGF21) in
human cerebrospinal fluid: Relationship with plasma FGF21 and body
adiposity. Diabetes. 60:2758–2762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liang Q, Zhong L, Zhang J, Wang Y,
Bornstein SR, Triggle CR, Ding H, Lam KS and Xu A: FGF21 maintains
glucose homeostasis by mediating the cross talk between liver and
brain during prolonged fasting. Diabetes. 63:4064–4075. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yang C, Jin C, Li X, Wang F, Mc Keehan WL
and Luo Y: Differential specificity of endocrine FGF19 and FGF21 to
FGFR1 and FGFR4 in complex with KLB. PLoS One. 7:e338702012.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sarruf DA, Thaler JP, Morton GJ, German J,
Fischer JD, Ogimoto K and Schwartz MW: Fibroblast growth factor 21
action in the brain increases energy expenditure and insulin
sensitivity in obese rats. Diabetes. 59:1817–1824. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hsuchou H, Pan W and Kastin AJ: The
fasting polypeptide FGF21 can enter brain from blood. Peptides.
28:2382–2386. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Owen BM, Ding X, Morgan DA, Coate KC,
Bookout AL, Rahmouni K, Kliewer SA and Mangelsdorf DJ: FGF21 acts
centrally to induce sympathetic nerve activity, energy expenditure,
and weight loss. Cell Metab. 20:670–677. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Owen BM, Bookout AL, Ding X, Lin VY, Atkin
SD, Gautron L, Kliewer SA and Mangelsdorf DJ: FGF21 contributes to
neuroendocrine control of female reproduction. Nat Med.
19:1153–1156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bookout AL, de Groot MH, Owen BM, Lee S,
Gautron L, Lawrence HL, Ding X, Elmquist JK, Takahashi JS,
Mangelsdorf DJ and Kliewer SA: FGF21 regulates metabolism and
circadian behavior by acting on the nervous system. Nat Med.
19:1147–1152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Arase K, York DA, Shimizu H, Shargill N
and Bray GA: Effects of corticotropin-releasing factor on food
intake and brown adipose tissue thermogenesis in rats. Am J
Physiol. 255:e255–e259. 1988.PubMed/NCBI
|
|
57
|
Adams AC, Cheng CC, Coskun T and
Kharitonenkov A: FGF21 requires βklotho to act in vivo. PLoS One.
7:e499772012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Johnson CL, Weston JY, Chadi SA, Fazio EN,
Huff MW, Kharitonenkov A, Köester A and Pin CL: Fibroblast growth
factor 21 reduces the severity of cerulein-induced pancreatitis in
mice. Gastroenterology. 137:1795–1804. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Johnson CL, Mehmood R, Laing SW, Stepniak
CV, Kharitonenkov A and Pin CL: Silencing of the fibroblast growth
factor 21 gene is an underlying cause of acinar cell injury in mice
lacking MIST1. Am J Physiol Endocrinol Metab. 306:E916–E928. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Uonaga T, Toyoda K, Okitsu T, Zhuang X,
Yamane S, Uemoto S and Inagaki N: FGF-21 enhances islet engraftment
in mouse syngeneic islet transplantation model. Islets. 2:247–251.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kharitonenkov A and Adams AC: Inventing
new medicines: The FGF21 story. Mol Metab. 3:221–229. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xu J, Stanislaus S, Chinookoswong N, Lau
YY, Hager T, Patel J, Ge H, Weiszmann J, Lu SC, Graham M, et al:
Acute glucose-lowering and insulin-sensitizing action of FGF21 in
insulin resistant mouse models-association with liver and adipose
tissue effects. Am J Physiol Endocrinol Metab. 297:E1105–E1114.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
So WY, Cheng Q, Chen L, Evans-Molina C, Xu
A, Lam KS and Leung PS: High glucose represses β-klotho expression
and impairs fibroblast growth factor 21 action in mouse pancreatic
islets: Involvement of peroxisome proliferator-activated receptor γ
signaling. Diabetes. 62:3751–3759. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tyynismaa H, Carroll CJ, Raimundo N,
Ahola-Erkkilä S, Wenz T, Ruhanen H, Guse K, Hemminki A,
Peltola-Mjøsund KE, Tulkki V, et al: Mitochondrial myopathy induces
a starvationlike response. Hum Mol Genet. 19:3948–3958. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kim KH, Jeong YT, Oh H, Kim SH, Cho JM,
Kim YN, Kim SS, Kim DH, Hur KY, Kim HK, et al: Autophagy deficiency
leads to protection from obesity and insulin resistance by inducing
Fgf21 as a mitokine. Nat Med. 19:83–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gälman C, Lundåsen T, Kharitonenkov A,
Bina HA, Eriksson M, Hafström I, Dahlin M, Amark P, Angelin B and
Rudling M: The circulating metabolic regulator FGF21 is induced by
prolonged fasting and PPARalpha activation in man. Cell Metab.
8:169–174. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Christodoulides C, Dyson P, Sprecher D,
Tsintzas K and Karpe F: Circulating fibroblast growth factor 21 is
induced by peroxisome proliferator-activated receptor agonists but
not ketosis in man. J Clin Endocrinol Metab. 94:3594–3601. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Mráz M, Lacinová Z, Kaválková P,
Haluzíková D, Trachta P, Drápalová J, Hanušová V and Haluzík M:
Serum concentrations of fibroblast growth factor 19 in patients
with obesity and type 2 diabetes mellitus: The influence of acute
hyperinsulinemia, very-low calorie diet and PPAR-α agonist
treatment. Physiol Res. 60:627–636. 2011.PubMed/NCBI
|
|
69
|
Dushay J, Chui PC, Gopalakrishnan GS,
Varela-Rey M, Crawley M, Fisher FM, Badman MK, Martinez-Chantar ML
and Maratos-Flier E: Increased fibroblast growth factor 21 in
obesity and nonalcoholic fatty liver disease. Gastroenterology.
139:456–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Laeger T, Henagan TM, Albarado DC, Redman
LM, Bray GA, Noland RC, Münzberg H, Hutson SM, Gettys TW, Schwartz
MW and Morrison CD: FGF21 is an endocrine signal of protein
restriction. J Clin Invest. 124:3913–3922. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Mraz M, Bartlova M, Lacinova Z, Michalsky
D, Kasalicky M, Haluzikova D, Matoulek M, Dostalova I, Humenanska V
and Haluzik M: Serum concentrations and tissue expression of a
novel endocrine regulator fibroblast growth factor-21 in patients
with type 2 diabetes and obesity. Clin Endocrinol (Oxf).
71:369–375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Chen WW, Li L, Yang GY, Li K, Qi XY, Zhu
W, Tang Y, Liu H and Boden G: Circulating FGF-21 levels in normal
subjects and in newly diagnose patients with type 2 diabetes
mellitus. Exp Clin Endocrinol Diabetes. 116:65–68. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chavez AO, Molina-Carrion M, Abdul-Ghani
MA, Folli F, Defronzo RA and Tripathy D: Circulating fibroblast
growth factor-21 is elevated in impaired glucose tolerance and type
2 diabetes and correlates with muscle and hepatic insulin
resistance. Diabetes Care. 32:1542–1546. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fisher FM, Chui PC, Antonellis PJ, Bina
HA, Kharitonenkov A, Flier JS and Maratos-Flier E: Obesity is a
fibroblast growth factor 21 (FGF21) resistant state. Diabetes.
59:2781–2789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lundasen T, Hunt MC, Nilsson LM, Sanyal S,
Angelin B, Alexson SE and Rudling M: PPARalpha is a key regulator
of hepatic FGF21. Biochem Biophys Res Commun. 360:437–440. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hale C, Chen MM, Stanislaus S,
Chinookoswong N, Hager T, Wang M, Véniant MM and Xu J: Lack of
overt FGF21 resistance in two mouse models of obesity and insulin
resistance. Endocrinology. 153:69–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Maruthur NM: The growing prevalence of
type 2 diabetes: Increased incidence or improved survival? Curr
Diab Rep. 13:786–794. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kharitonenkov A, Beals JM, Micanovic R,
Strifler BA, Rathnachalam R, Wroblewski VJ, Li S, Koester A, Ford
AM, Coskun T, et al: Rational design of a fibroblast growth factor
21-based clinical candidate, LY2405319. PLoS One. 8:e585752013.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Gaich G, Chien JY, Fu H, Glass LC, Deeg
MA, Holland WL, Kharitonenkov A, Bumol T, Schilske HK and Moller
DE: The effects of LY2405319, an FGF21 analog, in obese human
subjects with type 2 diabetes. Cell Metab. 18:333–340. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fisher FM, Estall JL, Adams AC, Antonellis
PJ, Bina HA, Flier JS, Kharitonenkov A, Spiegelman BM and
Maratos-Flier E: Integrated regulation of hepatic metabolism by
fibroblast growth factor 21 (FGF21) in vivo. Endocrinology.
152:2996–3004. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Mai K, Bobbert T, Groth C, Assmann A,
Meinus S, Kraatz J, Andres J, Arafat AM, Pfeiffer AF, Möhlig M and
Spranger J: Physiological modulation of circulating FGF21:
Relevance of free fatty acids and insulin. Am J Physiol Endocrinol
Metab. 299:E126–E130. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Adams AC, Halstead CA, Hansen BC, Irizarry
AR, Martin JA, Myers SR, Reynolds VL, Smith HW, Wroblewski VJ and
Kharitonenkov A: LY2405319, an engineered FGF21 variant, improves
the metabolic status of diabetic monkeys. PLoS One. 8:e657632013.
View Article : Google Scholar : PubMed/NCBI
|














