Introduction
During pregnancy, the maternal cardiovascular system
is subjected to volume overload (1).
However, pressure overload is not observed unless the pregnancy is
complicated by gestational hypertension (1). The increase in blood volume activates
several cardiac adaptive mechanisms, which results in the
enlargement of the left and right heart chambers and an increase in
cardiac mass (1). The most evident
increase in volume overload occurs during the third trimester
(2). The changes that take place
during physiological pregnancy are a form of physiological
reversible hypertrophy. Alterations in the diastolic function and
contractility have been described previously (3,4). For
example, lack of fibrosis accompanying the hypertrophy allows the
cardiac muscle to maintain or even increase its lusitropic
capacities.
The underlying molecular mechanisms of the changes
to cardiac structure and function are not fully understood
(5). An emerging mediator involved
in the adaptive processes are microRNAs (miRNAs/miRs). Several
miRNAs have been linked to cardiovascular diseases, and even
proposed as cardiovascular biomarkers (6). Furthermore, miRNAs have been shown to
be involved in the adaptation of the cardiovascular system to
exercise, another situation which where physiological hypertrophy
is observed (7).
Studies of the role of miRNAs in cardiovascular
alterations associated with pregnancy is limited primarily to
preeclampsia (8,9). Additionally, the potential involvement
of miRNAs in peripartum cardiomyopathy is the subject of only a
limited number of published studies (10,11). As
miRNAs regulate gene expression under specific conditions
influencing cellular activity, it was hypothesized that they may
serve a crucial role in cardiac adaptation to pregnancy-related
volume overload (12). MicroRNAs
associated with stimulation or inhibition of cardiac hypertrophy
and fibrosis were chosen for further inspection.
Detailed information regarding the expression of
cardiac miRNAs in pregnancy may improve our understanding of the
physiological adaptations which take place to accommodate volume
overload, and additionally may serve as a reference for various
pathological conditions, for example dilated cardiomyopathy. The
aim of the present study was to analyze the profile of miRNAs
associated with cardiac hypertrophy and fibrosis during the period
of maximum volume overload. To identify potential
pregnancy-specific changes in the expression of cardiac miRNAs, the
results obtained in pregnant women were compared with those from
non-pregnant healthy controls.
The overall hypothesis of the present study was that
there is a difference in the expression of cardiac miRNAs between
pregnant and non-pregnant women. The specific hypotheses were: i)
prohypertrophic and antifibrotic miRNAs would exhibit increased
expression in the third trimester of physiologic pregnancy; ii)
antihypertrophic and profibrotic miRNAs would exhibit decreased
expression in the third trimester of physiological pregnancy; iii)
the Δ cycle quantification (ΔCq) values of prohypertrophic miRNAs
would be positively correlated with the thickness of the left
ventricle walls; and iv) the ΔCq values of profibrotic miRNAs would
be correlated with diastolic dysfunction parameters.
Materials and methods
Study population and protocol
In the present case-controlled study, conducted
between October 2014 and June 2017, healthy women >18 years of
age with singleton pregnancies and gestational age corresponding to
the third trimester were included. The median age of pregnant women
was 31 years (range 20-41 years) and of the non-pregnant controls
was 28 years (range 21-43 years). Women with a history of
cardiovascular diseases or interventions, gestational hypertension,
or preeclampsia or eclampsia in the current pregnancy were excluded
from the study. The results obtained in the group of pregnant women
were compared with those obtained in healthy young non-pregnant
female volunteers. Volunteers were enlisted by disseminating
information about the study through posters distributed in the
Institute of Mother and Child and announcements during classes with
nursing students. Both pregnant women and the controls underwent an
echocardiographic examination to assess cardiac morphology, valve
function, systolic and diastolic function. In the present study, a
focus was placed on select echocardiographic variables describing
cardiac morphology and function. For cardiac morphology, these were
diastolic diameters of intraventricular septum, posterior wall and
left ventricle. For systole, these were systolic ejection fraction
and maximal blood flow velocity of the aortic valve. Additionally,
diastolic function (peak velocity of early diastolic transmitral
flow to late transmitral flow ratio, early diastolic velocity of
medial and lateral part of mitral annulus) was assessed. The
protocol used in the present study was approved by the Local
Bioethics Committee at the Institute of Mother and Child, and
written informed consent was obtained from all participants. The
study was performed in accordance with the guidelines described in
the Declaration of Helsinki (13).
Sample preparation and RNA
extraction
Upon collection, whole blood samples were
centrifuged at 25˚C for 20 min at 2,000 x g, and obtained sera were
stored at -80˚C until further analysis. Total RNA containing a
fraction of miRNA was extracted from 400 µl of each serum sample
using a mirVana PARIS kit (Ambion; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol.
Determination of miRNA expression
Expression of miRNAs was analyzed using Custom
TaqMan Array miRNA Cards (Applied Biosystems; Thermo Fisher
Scientific, Inc.). First, selected miRNAs were used to synthesize
cDNA using TaqMan MicroRNA Reverse Transcription kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Briefly, 6 ng isolated
RNA in a final volume of 3 µl was added to the reverse
transcription (RT) reaction mix consisting of 6 µl custom RT primer
pool, 0.3 µl dNTPs with dTTP (100 mM), 3 µl MultiScribe Reverse
Transcriptase (50 U/µl), 1.5 µl 10x RT buffer, 0.19 µl RNase
inhibitor (20 U/µl) (Applied Biosystems; Thermo Fisher Scientific,
Inc) and 1.01 µl nuclease-free water (Ambion; Thermo Fisher
Scientific, Inc.). cDNA was synthesized according to the
manufacturer's protocol (Applied Biosystems; Thermo Fisher
Scientific, Inc). Subsequently, the preamplification reaction was
performed using TaqMan PreAmp Master mix and 2X custom PreAmp
primer pool (Applied Biosystems; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Briefly, 22.5 µl of the
reaction mix was added to 2.5 µl of each product from the reverse
transcription, and then, the reaction was amplified using
thermocycling conditions specified in the manufacturer's protocol
(Applied Biosystems; Thermo Fisher Scientific, Inc). After the
reaction was complete, each PreAmp product was diluted 8-fold with
0.1X TE buffer (pH 8.0) (Ambion; Thermo Fisher Scientific, Inc.) to
a final volume of 200 µl. To prepare the qPCR reaction mix, 1.13 µl
diluted PreAmp product was added to the reaction mix consisting of
56.25 µl TaqMan Universal MasterMix II, No AmpErase UNG (2X)
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and 55.12 µl
nuclease-free water (Ambion; Thermo Fisher Scientific, Inc.), and
mixed. Subsequently, 100 µl of the complete qPCR reaction mix was
loaded into a single port on the TaqMan Array miRNA card,
centrifuged (2x1 min, 331 x g at 25˚C) and mechanically sealed.
Each qPCR reaction was performed in triplicate using a ViiA7
Real-Time PCR system using thermocycling conditions specified by
the manufacturer (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The Cq cut-off for the detection was 40 cycles. The results
of qPCR were analyzed using automatic baseline and manual threshold
set at 0.2 for all samples. U6 small nuclear RNA was used as the
endogenous control. Relative expression of analyzed miRNAs was
calculated using the 2-ΔΔCq method (14). The expression levels of the following
24 miRNAs were analyzed: miR-1 (assay ID 000385), miR-15b (assay ID
000390), miR-17-5 (assay ID 000393), miR-21 (assay ID 000397),
miR-22 (assay ID 002301), miR-26a (assay ID 000404), miR-27b (assay
ID 000409), miR-29-a (assay ID 002112), miR-29c (assay ID 000415),
miR-30c (assay ID 00419), miR-34a (assay ID 000425), miR-101 (assay
ID 002253), miR-124 (assay ID 001182), miR-133a (assay ID 000458),
miR-146a (assay ID 000468), miR-191 (assay ID 002299), miR-195
(assay ID 000494), miR-199a-3p (assay ID 002304), miR-199b (assay
ID 000500), miR-208a-5p (assay ID 462036_mat), miR-210 (assay ID
000512), miR-222 (Assay ID 000525), miR-328 (assay ID 00543) and
miR-1249 (assay ID 002868). Assay ID numbers refer to the catalogue
numbers of TaqMan Real-time PCR assays (Thermo Fisher Scientific
Inc.).
Analysis of miRNA results
miRNA expression is presented as frequencies,
(number of subjects in whom the expression of a given miRNA
exceeded the detection limit divided by the sample size) normalized
to the U6 ΔCq values, and as the fold change difference in miRNA
expression between pregnant and non-pregnant women.
First, the expression of miRNAs was measured as Cq
values. In brief the Cq value was the number of cycles of
amplification of the selected miRNA that allowed for their
detection. Then, the ΔCq values were obtained by subtracting U6
from the Cq value of the analyzed miRNA for each participant. The
distributions of U6 in both groups were normal, and the
between-group differences in U6 expression did not differ
significantly (mean 31.94 for non-pregnant controls vs. 31.13 for
pregnant women, P=0.09). Additionally, in a subgroup of patients,
an external spike, cel-39 (exogenous control molecule), was used.
The median value of cel-39 in the pregnancy group was 20.26±2.06
(range 18.7-33.5) and for the control group, 20.4±1.6 (range
19.6-24.5).
ΔCq should be interpreted as a marker of expression
levels. The expression of each miRNA is inversely proportional to
its ΔCq value. The lower the ΔCq values, the higher the expression
levels of a given miRNA. Conversely, larger ΔCq values should be
interpreted as lower expression levels of miRNAs.
Whenever the ΔCq value for a given miRNA differed
significantly between the study groups, the fold change was
calculated. First the ΔΔCq value was obtained by subtracting the
control groups median ΔCq value from the pregnancy groups median
ΔCq value of the analyzed miRNA. Then, fold change was calculated
using 2-ΔΔCq formula (14). The fold change is a measure of the
difference in the expression levels of a given compound between two
groups.
Statistical analysis
As the distributions of most Cq values was skewed,
non-parametric tests were used for statistical analysis.
Statistical characteristics of continuous variables are presented
as medians and ranges/interquartile ranges (IQR), except for the
hemodynamic and echocardiographic parameters, which are presented
as the mean ± standard deviation, as they were normally distributed
across both groups. Distributions of categorical variables are
presented as numbers (percentages). Statistical significances of
between-group comparisons in the non-parametric variables were
verified using a Mann-Whitney U-test. Distributions of nominal
variables were compared using a χ2 test. Relationships
between the expression levels of the selected miRNAs and
echocardiographic parameters were analyzed using Spearman's rank
correlation coefficient analysis. All statistical calculations were
performed using theSAS/STAT version 14.3 (SAS Institute, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Characteristics of the selected
miRNAs
The analyzed cardiac miRNAs were categorized into
three groups. The first group consisted of miRNAs associated with
cardiac hypertrophy (miR-1, miR-17-5, miR-22, miR-34a, miR-124,
miR-133a, miR-195, miR-199a-3p, miR-199b, miR-210, miR-222 and
miR-1249). The second consisted of miRNAs associated with cardiac
hypertrophy and fibrosis (miR-15b, miR-21, miR-26a, miR-29-a,
miR-29c, miR-30c, miR-101, miR-146a, miR-191, miR-208a-5p and
miR-328). The third group included only one miRNA, miR-27b, a
marker of proper systolic function.
First, the expression levels of the selected cardiac
miRNAs were analyzed as dichotomous variables (present vs. absent)
and quantified based on the ΔCq values. Then, the between-group
differences in the expression levels of the selected miRNAs in
pregnant women and non-pregnant controls were determined, with the
fold-change values calculated whenever the between-group difference
turned out to be statistically significant.
Results
Group characteristics
The results obtained in a group of 61 pregnant women
were compared with those of 19 non-pregnant controls.
The median age of pregnant women was significantly
higher compared with non-pregnant controls [31 years (IQR 27-35)
vs. 28 years (IQR 22-32), P=0.02]. The study groups did not differ
significantly in terms of their (pre-pregnancy) body mass index
values (22.04 kg/m2 in pregnant women vs. 22.48
kg/m2 in the controls, P=0.77). A total of 36 (59%)
women from the pregnant group were primigravidas. Mean gestational
age at the time of the study was 32.8±2.6 weeks. No significant
between-group differences were found in terms of the number of past
pregnancies (P=0.8) (Table I).
 | Table IHemodynamic and echocardiographic
parameters. |
Table I
Hemodynamic and echocardiographic
parameters.
| Parameter | Pregnancy
groupa | Control
groupa | P-value |
|---|
| Systolic blood
pressure, mmHg | 115±10 | 120±11 | 0.16 |
| Diastolic blood
pressure, mmHg | 70.6±7 | 74.5±6 | 0.049b |
| Heart rate, beats per
minute | 87.7±11.6 | 76.6±16 | 0.0003d |
| Ejection fraction,
% | 62.2±4.7 | 65.9±3.6 | 0.0015c |
| Left ventricular
diameter at end-diastole, mm | 45.9±3.6 | 43.9±5.1 | 0.14 |
| Interventricular
septum thickness at end-diastole, mm | 8.7±1.4 | 7.9±1.1 | 0.028b |
| Left ventricular
posterior wall thickness at end-diastole, mm | 8.6±1.2 | 7.7±0.9 | 0.005c |
| E/A | 1.37±0.32 | 1.6±0.34 | 0.0084c |
| E/E'med | 5.86±2.5 | 6.17±1.1 | 0.19 |
| E/E'lat | 5.02±1.9 | 4.82±1.2 | 0.94 |
| Ao Vmax, m/s | 1.42±0.19 | 1.22±0.09 |
<0.0001d |
The study groups differed with regards to a few
hemodynamic and echocardiographic indices (Table I). A few study participants presented
with cardiovascular risk factors and comorbidities. The list of
concomitant diseases present in the controls included diabetes
mellitus (n=1), hypercholesterolemia (n=1), hypothyroidism (n=1)
and polycystic ovary syndrome (n=1). In turn, the group of pregnant
women included five women with gestational diabetes mellitus
without a past history of diabetes, four with hypercholesterolemia,
five with hypothyroidism and another five with asthma. The study
groups did not differ significantly in the frequency of physical
activity (P=0.61). The proportions of smokers in both groups were
low and did not differ significantly (3.5% in pregnant women vs.
5.3% in the controls; P=0.72).
Expression of cardiac miRNAs in the
third trimester
Amongst the miRNAs associated with cardiac
hypertrophy, pregnant women expressed miR-17-5 (58/61, 95%), miR-22
(22/50, 44%), miR133a (17/61, 28%), miR-195 (40/61, 66%), miR-199a
(60/61, 98%), miR-199b (2/61, 3%), miR-210 (22/61, 36%), miR-222
(58/61, 95%) and miR-1249 (17/61, 28%). The expression of miR-1 was
found in only one pregnant woman (1.67%), and none of the pregnant
women expressed miR-34a and miR-124.
All pregnant women expressed miRNAs associated with
cardiac hypertrophy and fibrosis (miR-21, miR-26a, miR-30c,
miR-146a and miR-191). Additionally, the other miRNAs from this
group, miR-15b (58/61, 95%), mir-29a (57/61, 93%), miR-29c (60/61,
98%), miR-101 (40/61, 66%) and miR-328 (58/61, 95%), were found to
be expressed in a considerable proportion of pregnant women. In
contrast, none of the study participants expressed miR-208a.
Expression of the only miRNA from the third group,
miR-27b, was observed in 41 out of 61 (67%) pregnant women.
Median ΔCq values for the selected
cardiac miRNAs in the third trimester
Median ΔCq values for all miRNAs, the expression of
which was observed in >50% of the study participants, are shown
in Table II.
| Table IIMedian ΔCq values for cardiac miRNAs
expressed in more than half of the study participants during the
third trimester. |
Table II
Median ΔCq values for cardiac miRNAs
expressed in more than half of the study participants during the
third trimester.
| Group | miRNA | Median ΔCq
values | Interquartile
range | Range |
|---|
| Hypertrophy | miR-17-5 | 1.68 | 0.73-2.82 | -1.2-6.4 |
| | miR-199a | 0.736 | 0.04-1.74 | -2.5-5.5 |
| | miR-222 | -0.17 | -2.79 | -4.4-4.0 |
| Hypertrophy and
fibrosis | miR-21 | 8.6 | 7.73-9.45 | 4.6-12.7 |
| | miR-26a | -2.17 | -1.89 | -6.3-2.9 |
| | miR-30c | 0.2 | -1.89 | -3.6-3.8 |
| | miR-146a | -2.51 | -1.66 | -5.4-0.9 |
| | miR-191 | -2.26 | -1.84 | -5.7-1.6 |
| | miR-29a | 1.49 | 0.4-2.44 | -2.2-5.3 |
| | miR-29c | 0.53 | -3.05 | -2.5-8.5 |
| | miR-101 | 4.59 | 2.7-7.75 | -2.0-10.8 |
| | miR-15b | 1.92 | 1.15-2.75 | -1.6-8.2 |
| | miR-328 | 1.58 | 0.87-3.17 | -2.8-8.5 |
| Systolic
function | miR-27 | 3.09 | 2.0-6.35 | -0.5-12.5 |
Comparison with the control group
Pregnant women and non-pregnant controls did not
differ significantly in terms of the expression frequencies of all
analyzed miRNAs (Table III).
However, several statistically significant between-group
differences were found with regards to ΔCq values, suggesting that
pregnant women differed from non-pregnant controls in terms of the
expression levels of certain miRNAs (Table IV). The between-group comparisons of
the ΔCq values were performed solely for the miRNAs that were
expressed in >30% of the study subjects. The expression levels
of miRNAs which were significant differentially expressed between
pregnant and non-pregnant women or at least a trend towards
differential expression (P<0.1) was found, stratified according
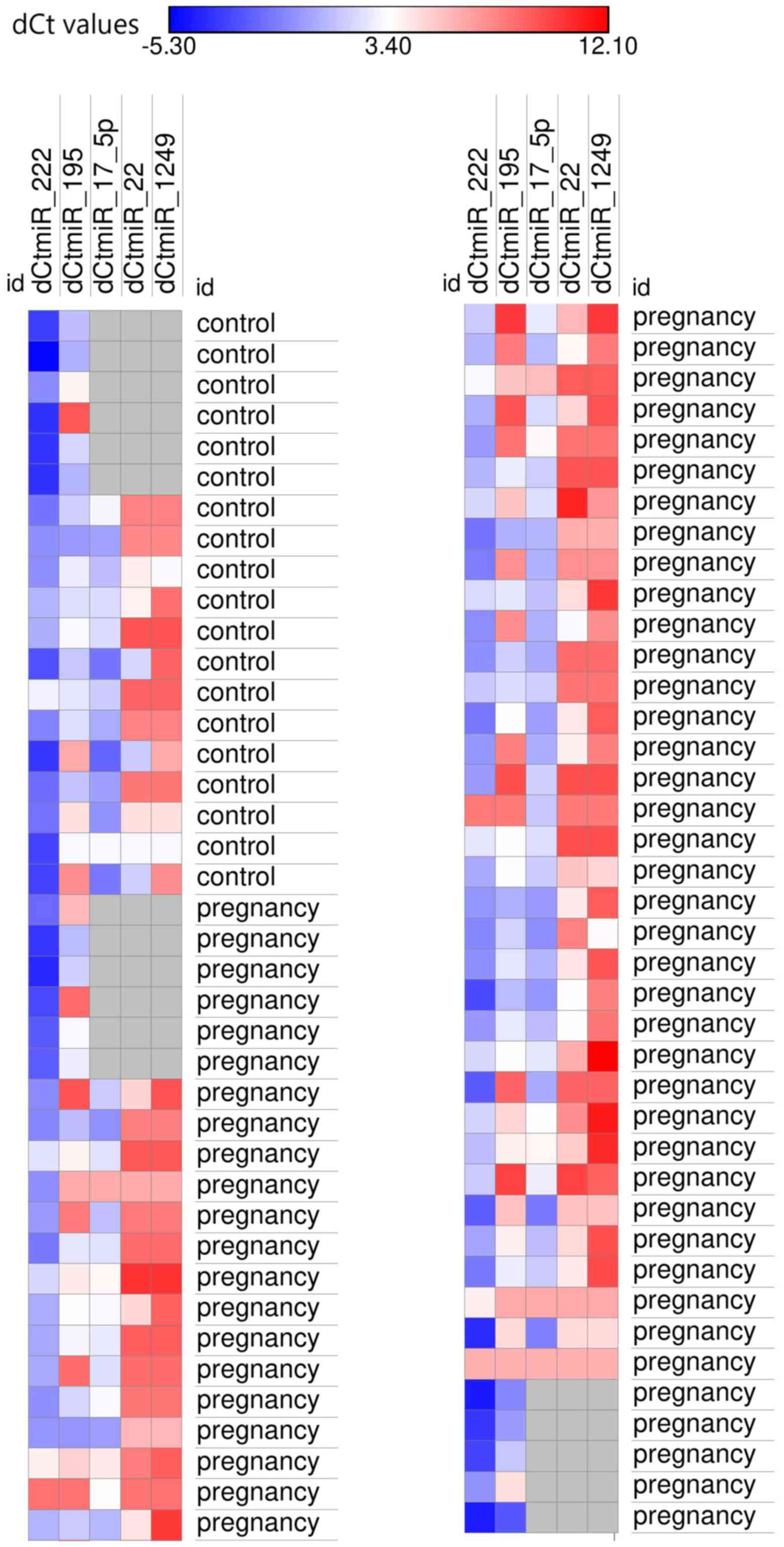
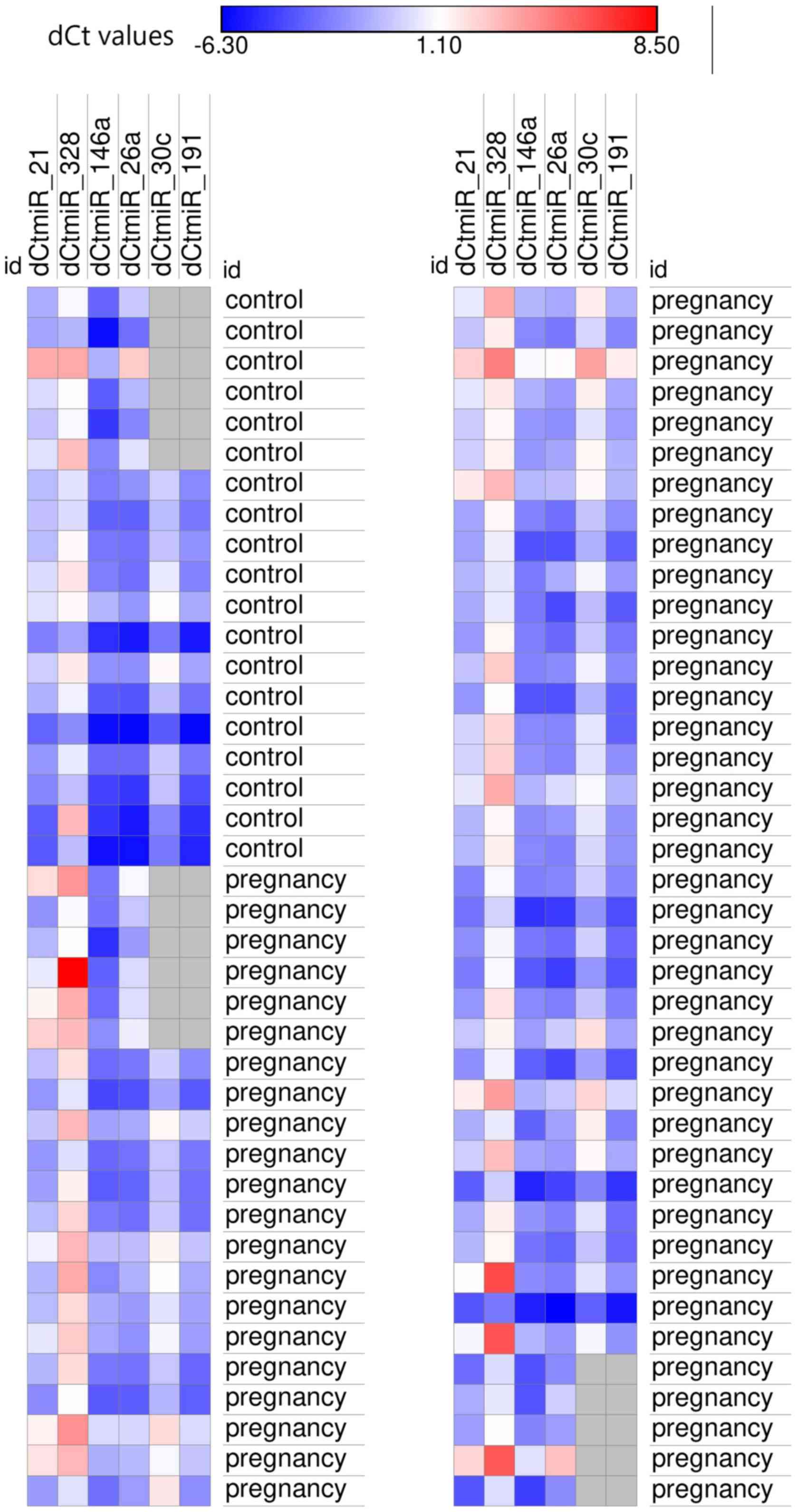
to their function, are presented in Figs. 1 and 2.
| Table IIIDifferences in the expression
frequencies of selected microRNAs between healthy pregnant women
and non-pregnant controls. |
Table III
Differences in the expression
frequencies of selected microRNAs between healthy pregnant women
and non-pregnant controls.
| miR | Pregnant women, n
(%) | Control group, n
(%) | P-value |
|---|
| miR-1 | 1/61(1.6) | 1/19 (5.3) | 0.4 |
| miR-17-5 | 58/61 (95.1) | 18/19 (94.7) | 1.0 |
| miR-22 | 33/61 (54.1) | 11/19 (57.9) | 0.8 |
| miR-34a | 0/61 (0) | 1/19 (5.3) | 0.2 |
| miR-124a | 11/61(18) | 6/19 (31.6) | 0.2 |
| miR-133a | 17/61 (27.9) | 5/19 (26.3) | 0.9 |
| miR-195 | 40/61 (65.6) | 14/19 (73.7) | 0.5 |
| miR-199a-3p | 60/61 (98.4) | 18/19 (94.7) | 0.42 |
| miR-199b | 2/61 (3.3) | 0/19 (0) | 1.0 |
| miR-210 | 22/61(36) | 6/19 (31.6) | 0.7 |
| miR-222 | 58/61 (95.1) | 19/19(100) | 1.0 |
| miR-1249 | 17/61 (27.9) | 7/19 (36.8) | 0.5 |
| miR-15b | 58/61 (95.1) | 16/19 (84.2) | 0.1 |
| miR-21 | 61/61(100) | 19/19(100) | 1.0 |
| miR-26a | 61/61(100) | 19/19(100) | 1.0 |
| miR-29a | 57/61 (93.4) | 15/19(79) | 0.1 |
| miR-29c | 60/61 (98.4) | 17/19 (89.5) | 0.1 |
| miR-30c | 61/61(100) | 19/19(100) | 1.0 |
| miR-101 | 35/61 (57.4) | 10/19 (52.6) | 0.7 |
| miR-146a | 61/61(100) | 19/19(100) | 1.0 |
| miR-191 | 61/61(100) | 19/19(100) | 1.0 |
| miR-208a-5p | 0/61 (0) | 0/19 (0) | 1.0 |
| miR-328 | 58/61 (95.1) | 18/19 (94.7) | 1.0 |
| miR-27b | 41/61 (67.2) | 9/19 (47.4) | 0.1 |
| Table IVDifferences in the expression levels
of selected miRNAs between healthy pregnant women and non-pregnant
controls. |
Table IV
Differences in the expression levels
of selected miRNAs between healthy pregnant women and non-pregnant
controls.
| | Pregnant women | Control group | |
|---|
| miRNA | Median ΔCq
(IQR) | Range of ΔCq
values | Median ΔCq
(IQR) | Range of ΔCq
values | P-value |
|---|
| miR-17-5 | 1.68 (0.8-3.0) | (-1.68)-4.96 | 0.56
(-0.3-2.2) | -1.18-6.35 | 0.056 |
| miR-22 | 6.31 (4.8-8.3) | 3.18-10.85 | 4.53 (3.2-7.7) | 1.62-9.28 | 0.08 |
| miR-195 | 3.79 (2.2-7.2) | (-2.32)-10.15 | 2.3 (1.4-3.5) | -0.08-9.17 | 0.046a |
| miR-199a-3p | 0.74
(0.05-1.8) | (-2.54)-7.96 | 0.33
(-1.4-1.3) | -3.78-4.12 | 0.26 |
| miR-210 | 7.7 (5.9-9.0) | 1.34-12.38 | 7.27 (3.9-8.2) | -0.28-5.92 | 0.96 |
| miR-222 | -0.17
(-1.2-1.6) | (-4.4)-8.19 | -1.58
[-3.3-(-0.5)] | -5.2-2.9 | 0.0038b |
| miR-1249 | 8.45 (7.7-9.3) | 3.54-12.06 | 7.66 (6.3-8.3) | 3.2-9.3 | 0.02a |
| miR-15b | 1.92 (1.2-2.8) | (-1.6)-8.2 | 1.14
(-0.3-3.4) | -1.83-7.66 | 0.44 |
| miR-21 | 8.6 (7.7-9.4) | 4.63-12.7 | 7.66 (6.4-8.3) | 2.06-9.28 | 0.0095b |
| miR-26a | -2.17
[(-3.1-(-1.2)] | (-6.26)-2.9 | -3.09
[-4.2-(-2.0)] | -6.1-2.65 | 0.069 |
| miR-29a | 1.49 (0.4-2.4) | (-2.24)-9.83 | 1.74 (0.8-3.1) | -1.42-7.7 | 0.37 |
| miR-29c | 0.53
(-0.4-2.7) | (-2.53)-8.51 | 0.68
(-1.3-6.3) | -2.75-9.3 | 0.62 |
| miR-30c | 0.2 (-0.6-1.2) | (-3.56)-3.8 | -0.68
[-2.4-(-0.3)] | -3.7-1.23 | 0.02a |
| miR-101 | 4.6 (2.8-7.9) | (-2.05)-10.83 | 4.37 (1.9-7.6) | 0.25-9.17 | 0.49 |
| miR-146a | -2.51
[-3.4-(-1.6)] | (-5.35)-0.89 | -3.45
[-4.7-(-2.7)] | -5.96-(-1.07) | 0.01a |
| miR-191 | -2.26
[-3.3-(-1.5)] | (-5.7)-1.63 | -2.82
[-4.8-(-2.3)] | -6.13-(-1.37) | 0.052 |
| miR-328 | 1.58 (0.9-3.2) | (-2.79)-8.46 | 0.85
(-0.3-1.5) | -2.23-3.63 | 0.01a |
| miR-27b | 3.1 (2.0-6.4) | (-0.49)-12.51 | 4.36 (0.9-7.9) | -0.65-11.33 | 1 |
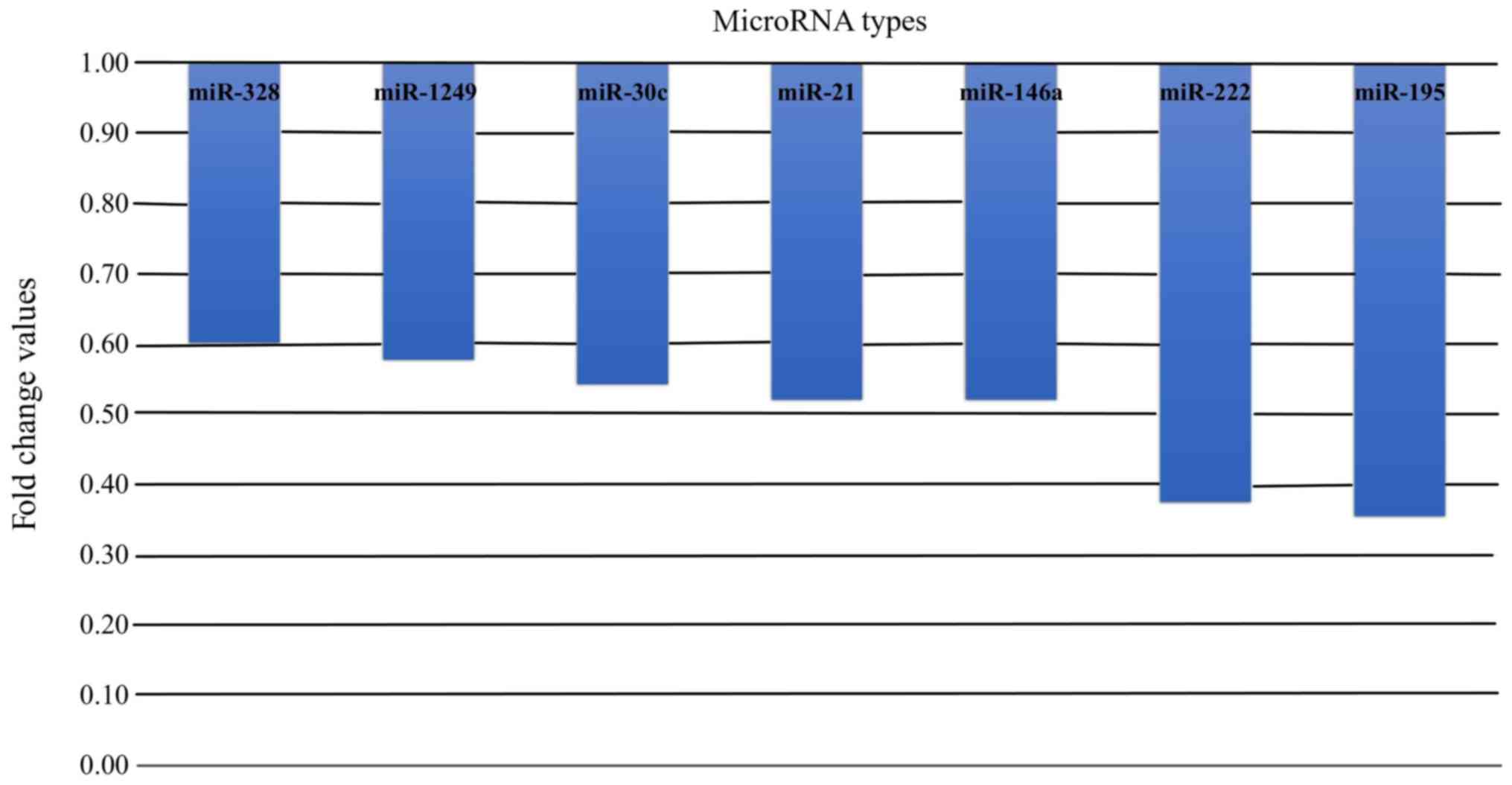
Fold-change
Among miRNAs with statistically significant
differences in the expression levels in healthy pregnant women and
non-pregnant controls, all the miRNAs, regardless of their
function, exhibited lower expression levels in pregnant women. The
fold-changes in the expression levels of these miRNAs in pregnant
women are shown in Fig. 3.
Correlations between select miRNAs and
echocardiographic parameters
Correlations with echocardiographic indices were
analyzed only if at least a trend towards a significant difference
in ΔCq values for a given miRNA were observed between pregnant
women and non-pregnant controls (P<0.1). These results are
presented in Table V).
| Table VCorrelations between
echocardiographic parameters and differentially expressed miRNAs
between pregnant women and the non-pregnant controls. |
Table V
Correlations between
echocardiographic parameters and differentially expressed miRNAs
between pregnant women and the non-pregnant controls.
| Echocardiographic
parameters | ΔCq miR-17-5 | ΔCq miR-22 | ΔCq miR-195 | ΔCq miR-222 | ΔCq miR-1249 | ΔCq miR-21 | ΔCq miR-26a | ΔCq miR-30c | ΔCq miR-146a | ΔCq miR-191 | miR-328 |
|---|
| Ejection fraction,
R-value | 0.06 | -0.17 | 0.26a | -0.13 | -0.005 | -0.04 | -0.14 | -0.07 | -0.02 | -0.12 | -0.07 |
| P | 0.66 | 0.24 | 0.04a | 0.37 | 0.97 | 0.74 | 0.29 | 0.64 | 0.9 | 0.4 | 0.58 |
| Ao Vmax,
R-value | -0.001 | -0.09 | -0.29a | -0.04 | 0.04 | -0.17 | -0.03 | -0.009 | -0.12 | -0.01 | -0.17 |
| P | 0.99 | 0.53 | 0.02a | 0.78 | 0.78 | 0.2 | 0.77 | 0.95 | 0.35 | 0.9 | 0.18 |
| Left ventricular
diameter at end-diastole, R-value | 0.11 | 0.16 | -0.05 | 0.04 | 0.09 | 0.04 | 0.03 | 0.09 | 0.03 | 0.06 | -0.01 |
| P | 0.43 | 0.28 | 0.72 | 0.78 | 0.55 | 0.74 | 0.79 | 0.54 | 0.82 | 0.67 | 0.95 |
| Interventricular
septum thickness at end-diastole, R-value | 0.006 | -0.22 | -0.03 | -0.2 | -0.25b | -0.33a | -0.09 | -0.27b | -0.19 | -0.2 | -0.11 |
| P | 0.97 | 0.13 | 0.8 | 0.13 | 0.08b | 0.02a | 0.49 | 0.06b | 0.14 | 0.17 | 0.41 |
| Left ventricular
posterior wall thickness at end-diastole, R-value | 0.16 | -0.1 | 0.02 | 0.28a | 0.05 | 0.13 | 0.07 | 0.15 | 0.2 | 0.11 | 0.002 |
| P | 0.28 | 0.5 | 0.9 | 0.03a | 0.73 | 0.37 | 0.58 | 0.29 | 0.11 | 0.46 | 0.99 |
| E/A, R-value | -0.24b | 0.007 | -0.19 | -0.24b | 0.22 | -0.11 | 0.06 | 0.03 | -0.21 | -0.004 | -0.22b |
| P | 0.09b | 0.96 | 0.15 | 0.07b | 0.12 | 0.41 | 0.64 | 0.84 | 0.11 | 0.98 | -0.09b |
| E/E'med,
R-value | 0.3b | 0.1 | 0.04 | 0.26 | 0.33a | 0.23 | 0.15 | 0.3b | 0.25b | 0.34a | 0.21 |
| P | 0.07b | 0.53 | 0.79 | 0.11 | 0.04a | 0.11 | 0.3 | 0.06b | 0.08b | 0.03a | 0.14 |
| E/E'lat,
R-value | 0.39a | 0.13 | 0.03 | 0.21 | 0.14 | 0.15 | 0.11 | 0.25 | 0.12 | 0.27 | 0.15 |
| P | 0.02a | 0.44 | 0.83 | 0.19 | 0.38 | 0.29 | 0.45 | 0.13 | 0.42 | 0.1 | 0.29 |
Morphology
Significant correlations and noticeable trends were
found between interventricular septum thickness and left
ventricular posterior wall thickness at end-diastole and the
expression levels of certain miRNAs. The intraventricular septum
thickness at end-diastole was inversely correlated with the ΔCq
value of miR-21. Furthermore, inversely correlated trends were
found between echocardiographic parameters and ΔCq values for
miR-1249 (correlation with intraventricular septum thickness) and
miR-30c (correlation with intraventricular septum thickness and
peak velocity of early diastolic transmitral flow to peak velocity
of early diastolic mitral annular motion at the septal side). In
turn, the left ventricular posterior wall thickness at end-diastole
correlated positively with ΔCq for miR-222 (Table V).
Systolic function
ΔCq for miR-195 was positively correlated with left
ventricular ejection fraction and showed an inverse correlation
with peak aortic velocity (Table
V).
Diastolic function
A number of significant correlations were found
between the expression levels of various miRNAs and parameters of
diastolic function. Interestingly, except for miR-17-5, miRNAs
which were correlated with parameters describing mitral annular
motion at the septal side did not show significant associations
with the parameters of mitral annular motion at the lateral side
and vice versa (Table V).
Correlations in non-pregnant
controls
In non-pregnant controls, the expression levels of
analyzed miRNAs did not correlate significantly with either
ejection fraction or peak aortic velocity, and the profiles of
correlations with the left ventricular posterior wall thickness,
interventricular septum thickness and parameters of diastolic
function differed compared with the pregnant women. Regarding
cardiac morphology, a negative correlation was observed between
posterior wall thickness and miR-222 (r=-0.8), miR-191 (r=-0.6) and
a positive correlation was observed between intraventricular wall
thickness and miR-22 (r=0.7), miR-17-5 (r=0.6), miR-191 (r=0.6),
miR-30c (r=0.6). In the aspect of diastolic function a negative
correlation was observed between peak velocity of early diastolic
transmitral flow to late transmitral flow ratio and miR-29c
(r=-0.5). miR-29c was negatively correlated with peak velocity of
early diastolic mitral annular motion at the septal side (r=-0.6).
There were no correlations between miR concentration and analyzed
systolic function parameters (data not shown).
Discussion
Relatively little is known regarding the role of
cardiovascular miRNAs in physiological myocardial hypertrophy
during pregnancy. In the present study, a detailed analysis of
cardiovascular miRNAs expressed during the third trimester of
pregnancy was performed, when pregnancy-associated volume overload
is at its highest.
During the design of the present study a difference
in cardiac miRNA expression between pregnant and non-pregnant women
was hypothesized. Although there were no differences in the
specific miRNAs expressed, the expression levels of miR-195,
miR-222, miR-1249, miR-21, miR-30c, miR-146a and miR-328 were lower
in pregnant women. The similarities in the qualitative composition
of the assessed miRNAs suggested that the mechanisms controlling
cardiomyocytes and extracellular matrix composition in these two
groups may be essentially the same; however, the differences in the
miRNA expression levels highlight their likely influence on
pregnancy related remodeling. Importantly, despite a volume
overload typical for physiological pregnancy, the expression levels
of miRNAs associated with myocardial damage, such as miR-208, were
not altered.
Neither the prohypertrophic, nor the antifibrotic
microRNAs exhibited increased expression in the third trimester of
physiological pregnancy. Among the prohypertrophic miRNAs, miR-30c
(15), miR-195(16) and miR-328(15) were downregulated compared with the
healthy controls. Interestingly, miR-222, an antifibrotic miRNA
which is inversely correlated with the degree of myocardial
fibrosis through inhibition of transforming growth factor-β
(17), was downregulated during
pregnancy. The decrease in miR-328 expression in pregnancy, first
reported in our previous study (18), has now been confirmed in a larger
cohort.
A hypothesis of the present study was that
expression of antihypertrophic and profibrotic miRNAs would be
decreased in the third trimester of physiological pregnancy.
However, expression of the studied antihypertrophic miRNAs were
similar in both groups. Among the profibrotic miRNAs, a significant
difference was observed in expression of miR-21. In myocardial
infarction models, miR-21 stimulated collagen synthesis (19,20).
Thus, it is hypothesized that the decrease in miR-21 expression may
protect the myocardium of pregnant women against fibrosis resulting
from volume overload in pregnancy.
Additionally a decrease in miR-146a expression was
observed, and this miRNA possesses both an antimetabolic function,
suppressing mitochondrial metabolism (21) and a profibrotic role (22). The decreased expression observed in
pregnant women may be an adaptive mechanism, allowing
cardiomyocytes to increase mitochondrial metabolism. Furthermore,
miR-146a is the only miRNA associated with peripartum
cardiomyopathy (21). In this
disease, miR-146a released form endothelial cells following
stimulation with 16 kDa N-terminal fragments of prolactin is
absorbed by cardiomyocytes, inhibiting their metabolic activity
(21). This mechanism may also
underlie the decrease in myocardial contractility observed in
peripartum cardiomyopathy (10).
To analyze potential associations between miRNAs and
echocardiographic indices, a series of statistical analyses was
performed. Echocardiographic data collected during the present
study were consistent with published evidence; pregnant women in
the third trimester possessed thicker cardiac walls, higher peak
aortic velocity, lower left ventricular ejection fraction and
altered parameters of diastolic function. These changes seem to be
consequences of volume overload in pregnancy that secures a fetus'
demand for nutrients delivered via maternal circulation.
The differences between correlations observed in
pregnant women and non-pregnant controls suggest that the analyzed
miRNAs may serve a role in myocardial remodeling during pregnancy,
and their function in pregnant women may differ than during
physiological conditions. A correlation between ΔCq values of
prohypertrophic miRNAs and left ventricle wall thickness was
hypothesized. Additionally, a dependence between profibrotic miRNAs
and diastolic left ventricle function parameters was also
hypothesized. The results suggested that hypertrophy of the
interventricular septum and free cardiac wall (represented by
posterior wall thickness) in pregnancy was achieved via different
underlying mechanisms. Interventricular septum thickness was
positively correlated with the expression of miR-21, whereas an
inverse correlation was found between the thickness of the
posterior wall and miR-222 expression. This observation was
surprising as miR-222 is considered a vital miRNA required for
proliferation and growth of cardiomyocytes in response to physical
exercise (23,24). In human and animal exercise models
miR-222 increases myosin heavy chain α/β ratio. In animal models
administration of miR-222 antagonist resulted in complete
abrogation of heart size enlargement in response to intensive
exercise (24). Based on the results
of the present and the aforementioned previous studies, adaption
mechanisms to exercise and pregnancy overload may have different
molecular basis, and the differences may be due the alternative
models of overload-in pregnancy the volume overload is constant and
stably increasing over several weeks, whereas during exercise,
overload appears in intervals and the increase is more varied and
short-lasting.
Analysis of miRNA expression and hemodynamic
parameters revealed an inverse correlation between the expression
of miR-195 and left ventricular ejection fraction, along with a
positive correlation between miR-195 expression levels and peak
aortic velocity. In contrast, none of the miRNAs were associated
with the assessed hemodynamic indices in non-pregnant women.
The correlations between the diastolic parameters
and expression of miR-17-5p seem to be of particular import. Higher
expression levels of miR-17-5p were observed to be associated with
improved relaxation, as shown by higher values of the E to A wave
ratio and lower estimated left ventricular filling pressures, shown
by lower values of the E to E'med and E to E'lat ratios (see
Table V for detailed results).
Experimental mouse models of myocardial infarction demonstrated
that knockdown of miR-17-5p expression decreased infarction area
and the area of fibrosis and stimulated endothelial growth
(25). Therefore, decreased
expression of miR-17-5p may serve a regulatory role in diastolic
function adaptation in pregnancy. Furthermore, there was a tendency
towards positive correlations between left ventricular myocardial
relaxation (E/A ratio) and the expression levels of miR-222 and
miR-328.
The present study has some limitations. Age
differences between studied groups needs to be taken into account
when interpreting the results. Age is a factor that can affect
miRNA expression levels. Although, the difference in age between
the groups was statistically significant, the numerical difference
was only 3 years. Available data suggests that the effect of age on
miRNA expression levels requires decades (26). Furthermore, there were no
correlations between expression of any of the miRNAs assessed with
age in any of the groups. Therefore, it is unlikely that age had a
significant impact on the observed differences in miRNA expression
levels. Another limitation was that the present study was a case
control study. Any differences in demographics could not be avoided
due to the design of the study.
The relatively low expression levels of the studied
miRNAs with a significant number of obtained Cq values >30
constitutes a limitation as well. This may have been a result of
low RNA input. In order to ensure the highest quality of results,
the RNA input was unified across all participants. The lowest
quantity of RNA obtained from the samples from all 80 participants
was 6 ng. Median Cq values of U6 (internal normalization) and
studied miRNA compared with median Cq values of cel-39 (external
normalization) highlights the fact that there the absolute
expression levels of the studied miRNAs was very low in both
groups, and not the result of a low RNA input.
In conclusion, while the profiles of cardiac miRNAs
expressed in healthy pregnant women and healthy non-pregnant
controls were similar, these two groups differed in terms of miRNA
expression levels of select miRNAs. In the third trimester of
physiological pregnancy, downregulation of miR-17-5p, miR-21,
miR-30c, miR-146a, miR-195, miR-222 and miR-328 expression was
observed. The differences in the correlations of echocardiographic
indices with miRNAs in pregnant and non-pregnant women suggests
that miRNAs may regulate both the structure and function of the
heart in pregnant women, influencing cardiac muscle thickness, as
well as systolic and diastolic function.
Acknowledgements
We would like to thank Professor Katarzyna
Szamotulska, Dr Leszek Soluch and Dr Katarzyna Markiewicz
(Institute of Mother and Child) for their assistance.
Funding
The present study was supported by a grant from the
Polish National Center of Science (grant no. NCN
2013/11/N/NZ5/03388).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors contributed to the conception and design
of the study and writing of the manuscript. ES, AB, KP, GS and AF
recruited the participants, obtained informed consent and were
responsible for the collection blood samples. AZ and MM performed
the microRNA assays. ES assisted during microRNA measurements. ES
managed the database. ES, AZ, GO, TM, MM and AF interpreted the
results of statistical analysis. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The protocol used in the present study was approved
by the Local Bioethics Committee at the Institute of Mother and
Child, and written informed consent was obtained from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sanghavi M and Rutherford JD:
Cardiovascular physiology of pregnancy. Circulation. 130:1003–1008.
2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Indira Devi C and Srikanth S:
Cardiovascular changes during pregnancy, labour and puerperium. Int
J Sci Res. 4:555–561. 2015.
|
|
3
|
Umar S, Nadadur R, Iorga A, Amjedi M,
Matori H and Eghbali M: Cardiac structural and hemodynamic changes
associated with physiological heart hypertrophy of pregnancy are
reversed postpartum. J Appl Physiol. 113:1253–1259. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fok WY, Chan LY, Wong JT, Yu CM and Lau
TK: Left ventricular diastolic function during normal pregnancy:
Assessment by spectral tissue Doppler imaging. Ultrasound Obstet
Gynecol. 28:789–793. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
5
|
Li J, Umar S, Amjedi M, Iorga A, Sharma S,
Nadadur RD, Regitz-Zagrosek V and Eghbali M: New frontiers in heart
hypertrophy during pregnancy. Am J Cardiovasc Dis. 2:192–207.
2012.PubMed/NCBI
|
|
6
|
Condorelli G, Latronico MVG and Cavarretta
E: microRNAs in cardiovascular diseases: Current knowledge and the
road ahead. J Am Coll Cardiol. 63:2177–2187. 2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Silva GJJ, Bye A, El Azzouzi H and Wisløff
U: MicroRNAs as important regulators of exercise adaptation. Prog
Cardiovasc Dis. 60:130–151. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Mohseni Z, Spaanderman MEA, Oben J, Calore
M, Derksen E, Al-Nasiry S, de Windt LJ and Ghossein-Doha C: Cardiac
remodelling and pre-eclampsia: An overview of overlapping miRNAs.
Ultrasound Obstet Gynecol. 52:310–317. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li J, Wu G, Cao Y and Hou Z: Roles of
miR-210 in the pathogenesis of pre-eclampsia. Arch Med Sci.
15:183–190. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Halkein J, Tabruyn SP, Ricke-Hoch M,
Haghikia A, Nguyen NQ, Scherr M, Castermans K, Malvaux L, Lambert
V, Thiry M, et al: MicroRNA-146a is a therapeutic target and
biomarker for peripartum cardiomyopathy. J Clin Invest.
123:2143–2154. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Yang Y, Rodriguez JE and Kitsis RN: A
microRNA links prolactin to peripartum cardiomyopathy. J Clin
Invest. 123:1925–1927. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
12
|
O'Brien J, Hayder H, Zayed Y and Peng C:
Overview of MicroRNA biogenesis, mechanisms of actions, and
circulation. Front Endocrinol (Lausanne). 9(402)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
World Medical Association. World Medical
Association Declaration of Helsinki: Ethical principles for medical
research involving human subjects. JAMA. 310:2191–2194.
2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Li C, Li X, Gao X, Zhang R, Zhang Y, Liang
H, Xu C, Du W, Zhang Y, Liu X, et al: MicroRNA-328 as a regulator
of cardiac hypertrophy. Int J Cardiol. 173:268–276. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
van Rooij E, Sutherland LB, Liu N,
Williams AH, McAnally J, Gerard RD, Richardson JA and Olson EN: A
signature pattern of stress-responsive microRNAs that can evoke
cardiac hypertrophy and heart failure. Proc Natl Acad Sci USA.
103:18255–18260. 2006.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Verjans R, Peters T, Beaumont FJ, van
Leeuwen R, van Herwaarden T, Verhesen W, Munts C, Bijnen M, Henkens
M, Diez J, et al: MicroRNA-221/222 family counteracts myocardial
fibrosis in pressure overload-induced heart failure. Hypertension.
71:280–288. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Szczerba E, Zajkowska A, Bochowicz A,
Pankiewicz K, Szewczyk G, Markiewicz K, Opolski G, Maciejewski T,
Małecki M and Fijałkowska A: Rise in antifibrotic and decrease in
profibrotic microRNA protect the heart against fibrosis during
pregnancy: A preliminary study. Adv Clin Exp Med. 27:867–872.
2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Liang H, Zhang C, Ban T, Liu Y, Mei L,
Piao X, Zhao D, Lu Y, Chu W and Yang B: A novel reciprocal loop
between microRNA-21 and TGFβRIII is involved in cardiac fibrosis.
Int J Biochem Cell Biol. 44:2152–2160. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
You XY, Huang JH, Liu B, Liu SJ, Zhong Y
and Liu SM: HMGA1 is a new target of miR195 involving isoprenaline
induced cardiomyocyte hypertrophy. Biochemistry (Moscow).
79:538–544. 2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Heggermont WA, Papageorgiou AP, Quaegebeur
A, Deckx S, Carai P, Verhesen W, Eelen G, Schoors S, van Leeuwen R,
Alekseev S, et al: Inhibition of microRNA-146a and overexpression
of its target dihydrolipoyl succinyltransferase protect against
pressure overload-induced cardiac hypertrophy and dysfunction.
Circulation. 136:747–761. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Feng B, Chen S, McArthur K, Wu Y, Sen S,
Ding Q, Feldman RD and Chakrabarti S: miR-146a-mediated
extracellular matrix protein production in chronic diabetes
complications. Diabetes. 60:2975–2984. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tao L, Bei Y, Zhang H, Xiao J and Li X:
Exercise for the heart: Signaling pathways. Oncotarget.
6:20773–20784. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Liu X, Xiao J, Zhu H, Wei X, Platt C,
Damilano F, Xiao C, Bezzerides V, Boström P, Che L, et al: miR-222
is necessary for exercise-induced cardiac growth and protects
against pathological cardiac remodeling. Cell Metab. 21:584–95.
2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Yang S, Fan T, Hu Q, Xu W, Yang J, Xu C,
Zhang B, Chen J and Jiang H: Downregulation of microRNA-17-5p
improves cardiac function after myocardial infarction via
attenuation of apoptosis in endothelial cells. Mol Genet Genomics.
293:883–894. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Ong J, Woldhuis RR, Boudewijn IM, van den
Berg A, Kluiver J, Kok K, Terpstra MM, Guryev V, de Vries M,
Vermeulen CJ, et al: Age-related gene and miRNA expression changes
in airways of healthy individuals. Sci Rep. 9(3765)2019.PubMed/NCBI View Article : Google Scholar
|