1. Introduction
It is well known that the general population may be
exposed to ‘naturally occurring asbestos’ (NOA) (1-4).
The term NOA refers to the mineral as a natural component of soils
or rocks as opposed to asbestos in commercial products, mining or
processing operations. NOA may be released as fibers into the air
by human activities or natural weathering processes that represent
a risk for human exposure (3).
A fiber is defined as a particle (length/diameter
ratio 3:1) with certain characteristics that make it respirable,
penetrating into the alveolar level and participating in gaseous
exchanges (5). In vitro,
in vivo and ex vivo studies have demonstrated that
the dimension, surface property, shape, cristallinity, chemical
composition, physical durability, exposure route, duration of
exposure, dose (6) and genetic
background of the host exposed (7)
are all determinants for the biological activities of a certain
fibrous element. Fiber dimensions are important because only fibers
with diameter <0.4 µm and length <10 µm are respirable to the
distal alveolar space. Dose is a crucial determinant for triggering
inflammation. Indeed, high doses over short periods promote an
acute neutrophil-predominant inflammation, whereas low doses over
prolonged exposure periods promote alveolar macrophage
(AM)-predominant chronic inflammation (7). In general, fibers greater than 20 µm in
length are associated with asbestosis, and fibers greater than 10
µm in length are the most carcinogenic (5).
Carcinogenic mineral fibers are divided into
asbestos and asbestiform fibers (8,9). The
term ‘asbestos’ is used to identify silicate minerals belonging to
two families: Amphiboles (amosite, anthophyllite, actinolite,
crocidolite, tremolite, anthracite) and serpentine (crysotile)
(6,10). Amphiboles are straight, rod-like
fibers, whereas serpentines are curvilinear fibers. Asbestos has a
significant industrial importance for its characteristics, its
abundance and its low cost. Asbestos has sound-absorbing and
sound-proofing properties, thermal stability at high temperatures,
good mechanical resistance, good resistance to chemical and
biological agents, and for this high versatility it has been widely
used in different areas, especially in the building industry
(11).
Exposure to asbestos fibers can cause several
diseases such as asbestosis, lung and bronchus cancer, malignant
mesothelioma (MM) of the pleura, peritoneum, pericardium and tunica
vaginalis testis, neoplasms of the ovary, larynx and trachea
carcinoma (11). There is evidence
that the inhalation of asbestos fibers can provoke two types of
inter-connected pathogenetic processes: Chronic inflammation and
carcinogenesis, involving the lung after inhalation and deposition
of asbestos fibers (12). Therefore,
it has been established that cancer frequently arises in areas of
chronic inflammation (13). Many
lines of evidence have highlighted the ability of asbestos fibers
to: Interfere with the mitotic apparatus; stimulate host cell
proliferation; induce genetic and epigenetic alterations; induce
cytotoxicity and fibrosis; produce oxidative stress by at least
three sources including fiber surface reactivity, release from
inflammatory cells especially AMs, and mitochondrial-derived ROS
release from inflammatory and other target cells such as lung
epithelial cells and mesothelial cells (7). The ROS production results in DNA
damage, release of inflammatory cytokines and growth factors that
collectively contribute to fiber pathogenicity (12,14) and
H2O2 production in mediating asbestos
pulmonary toxicity (7).
Several studies have reported a high incidence of MM
due to asbestos exposure in: Finland (15), California, USA (16), China (17), New Caledonia (18), Corsica (19), Cyprus (20) and Greece (21). Yet, in many cases, it has been
discovered that the cause of these MM cases has not been asbestos
but asbestiform fibers.
The term ‘asbestiform fiber’ is commonly used to
indicate erionite, winchite, magnesio-riebeckite, richterite, Libby
asbestos, antigorite and fluoro-edenite (FE) fibers. Erionite is a
mineral belonging to the zeolite family (22). Several studies have reported a high
incidence of MM due to erionite exposure in rural regions of
Turkey, Central Anatolia (23-27).
Furthermore, individuals exposed to erionite may develop
interstitial fibrosis and additional pulmonary pathology impacting
lung function and patient survival (28).
Clark and Nye counties, in southern Nevada, USA,
have shown a significantly high incidence of MM due to carcinogenic
fibers including erionite, winchite, magnesio-riebeckite, and
richterite (29). These are the same
fibrous minerals present in Libby, Montana, USA, where they have
been related to MM and other asbestos-related diseases (30).
Antigorite is a silicate mineral very similar in
chemical composition to chrysotile and its asbestiform variant is
present in serpentinite rocks associated with MM (31). Antigorite is found in the Western
Alps (Piemonte, Italy) (32,33), in North America, Australia-Oceania,
and Rowland Flat in South Australia (34,35).
Fluoro-edenite (FE), the amphibole of Biancavilla
(Sicily, Italy), is a silicate mineral belonging to the amphibole
family (36). This silicate mineral
has been identified in the lavic products of Monte Calvario from
stone quarries located in the southeast of Biancavilla (37), a small town of the Etnean volcanic
complex, in Sicily. This silicate mineral presents some
characteristics similar to the asbestos group (38,39); in
particular it presents the same morphological and compositional
aspect of the two fibrous phases tremolite and actinolite. The
mineralization process led to the development of large prismatic
crystals embedded in the matrix, small acicular crystals that line
cavities or also fibrous and asbestiform (37). The salient feature, which nonetheless
distinguishes the FE of Biancavilla not only from other fibrous
minerals, but also from all the other known amphiboles, is the very
anomalous composition characterized by high sodium, aluminum and
fluorine contents, in comparison to other known oncogenic minerals
(40).
Epidemiological studies have indeed confirmed that
FE fibers have shown similar effects to those already reported
after exposure to asbestos fibers (8,41-43)
including cell necrosis with release of high mobility group protein
B1 and activation of the Nalp3 inflammasone, leading to chronic
inflammation, DNA damage and carcinogenesis (44).
Several studies have reported a high incidence of MM
in Italy due to FE exposure in Biancavilla (45-48)
concerning the time window 1980-2009. All of the data suggest that
a mode of exposure to FE fibers is related to environmental
contamination, rather than specific occupational activities
(45). In fact, the stone material
from the quarry of Monte Calvario has been used locally for about
50 years for building purposes (8,49,50) and
none of the residents diagnosed with MM have been significantly
exposed to asbestos during their professional lives (12).
Diseases related to erionite and FE fibers present
with characteristics similar to asbestos-related pathologies. The
underlying modes of action of asbestosis, lung cancer and MM seem
to be different in regards to the fiber type, lung clearance, and
genetics. Several lines of evidence have led to the classification
of asbestos, erionite, and FE as Group 1 human carcinogens (51;
IARC, 1987).
Therefore, NOA represents an important environmental
concern. Thus, asbestos and asbestiform fibers continue to cause a
high health concern due to the long latency period of related
diseases.
2. Literature search methodology
This systematic review was carried out in accordance
with PICo criteria (52). The
review/research question was defined, using PICo criteria, by
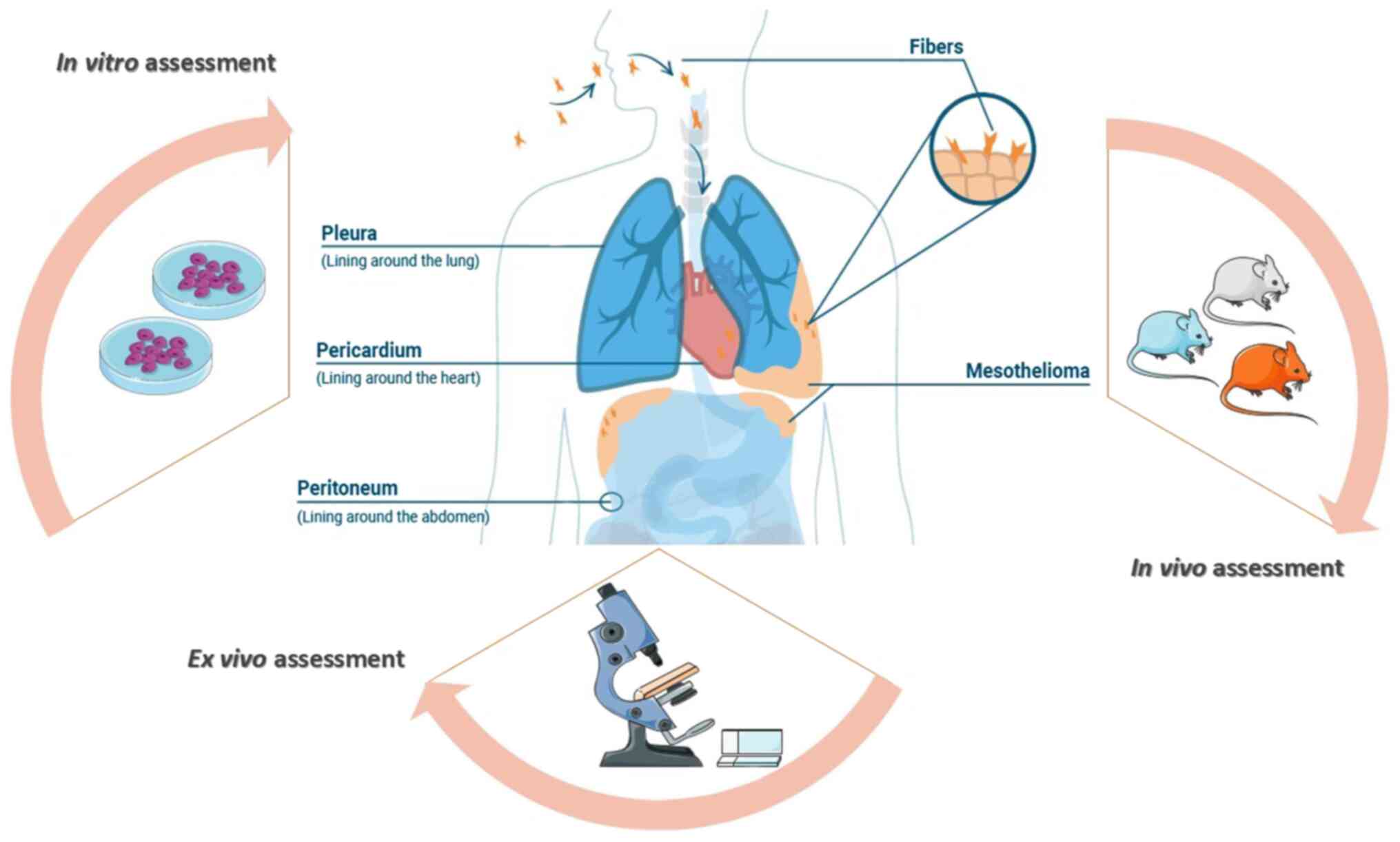
identifying: Population, interest, and context of research. What
are the in vitro, in vivo and ex vivo studies
correlated with MM due to FE exposure in Biancavilla? (Fig. 1). The research was performed by using
the following search term: ‘Fluoro-edenite fibers’. A search of the
research manuscripts suitable for inclusion in this systematic
review, was carried out and the research papers of significance
were collected and reviewed. The main topics and alternate terms
from our PICo question that were used for the search were:
Fluoro-edenite exposure, Fluoro-edenite, Fluoro-edenite fibers,
Fluoro-edenite fibres, Biancavilla, Biancavilla's exposure,
Malignant mesothelioma. The English language was used as a limit to
our search. SCOPUS and Medline (using PubMed as the search engine)
databases were used to search relevant research articles available
from March 4 to August 4, 2020 (Fig.
1).
Inclusion and exclusion criteria
The following inclusion criterion was adopted:
Experimental studies that assessed the effects of FE fiber exposure
in vitro, in vivo and ex vivo models. The
following exclusion criteria were applied: i) Scientific articles
that were not published in the English language; ii) review or
conference abstracts or letters to the editor; iii) experimental
studies that did not concern in vitro, in vivo and
ex vivo models. For duplicate studies, the article
containing further detailed information was solely included.
Quality assessment and data
extraction
Two reviewers (VF and CL) retrieved articles
independently. The title, abstract and full text of each
potentially pertinent study were reviewed. Any divergence on the
eligibility of the studies was determined by debate. The following
information was extracted from all qualified papers: Authors, year
of publication, and study characteristics.
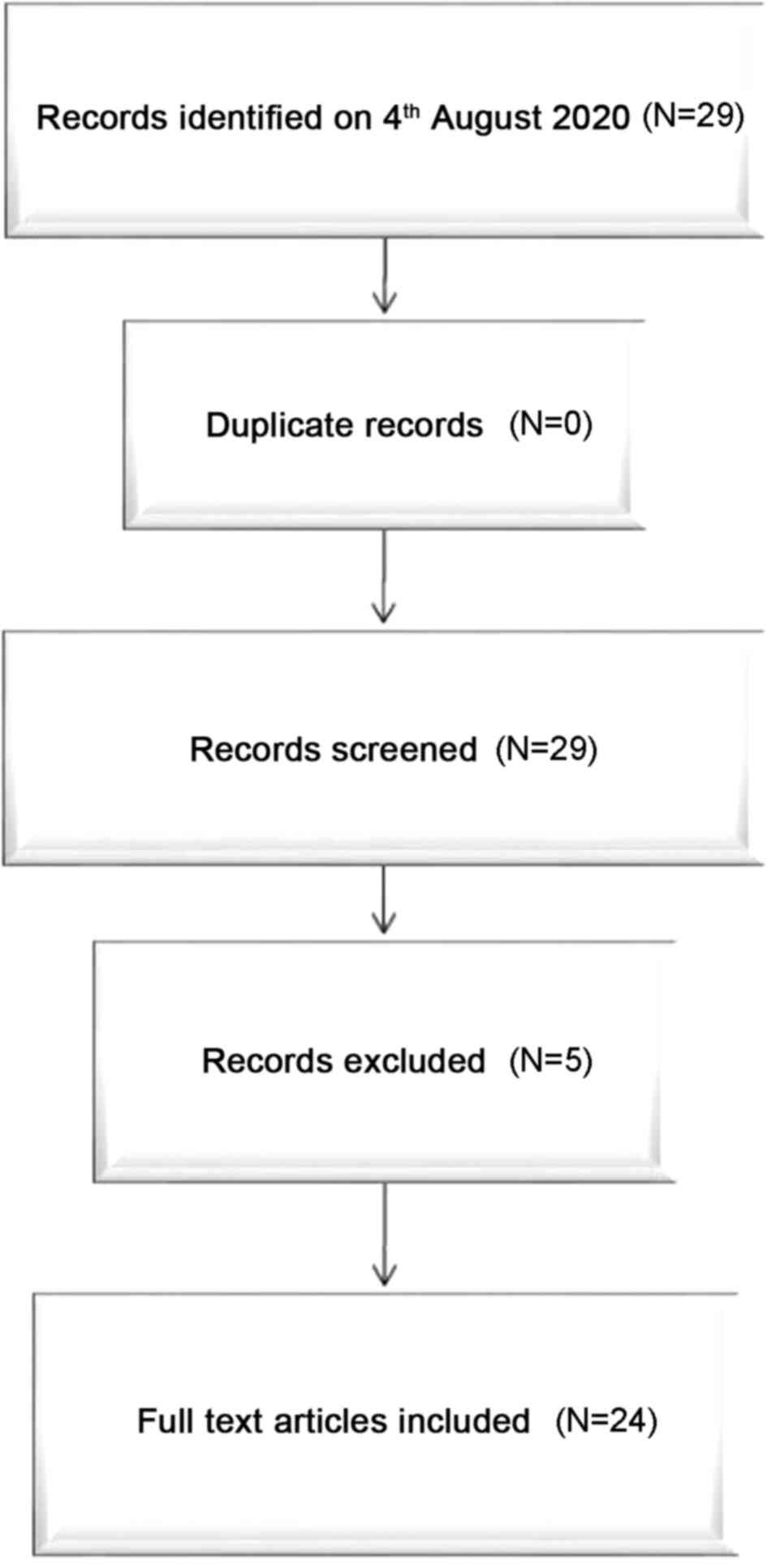
After a free search for scientific literature by
reviewers, a total of 29 documents were collected. In conclusion, 5
studies were disqualified after review of the manuscript. A total
of 24 studies satisfied the inclusion criteria and were included in
this systematic review. A flow-chart depicting the choice of
studies is documented in Fig. 2. The
information concerning authors, year of publication, and
characteristics of included studies have been included in Table I.
 | Table ICharacteristics of the studies
included in the systematic review. |
Table I
Characteristics of the studies
included in the systematic review.
| | (Refs.) | Samples |
|---|
| In vitro
N=11 | Travaglione et
al (12) | A549 cells |
| | Cardile et
al (60) | J774 cells |
| | Cardile et
al (64) | A549 cells |
| | | J774 cells |
| | | Human lung
fibroblasts |
| | Travaglione et
al (56) | A549 cells |
| | Cardile et
al (32) | MeT-5A cells |
| | | J774 cells |
| | Pugnaloni et
al (54) | MeT-5A cells |
| | | A549 cells |
| | | J774 cells |
| | Loreto et al
(63) | A549 cells |
| | Musumeci et
al (69) | MeT-5A cells |
| | | A549 cells |
| | Rapisarda et
al (74) | MeT-5A cells |
| | Rapisarda et
al (76) | Human lung
fibroblasts |
| | Filetti et
al (77) | MeT-5A cells JU77
cells |
| In vivo
N=2 | Soffritti et
al (36) | Sprague-Dawley
rats |
| | Belpoggi et
al (88) | Sprague-Dawley
rats |
| Ex vivo
N=11 | DeNardo et
al (82) | Lung tissue of
sheep |
| | Martinez et
al (43) | Lung tissue of
sheep |
| | Loreto et al
(83) | Lung tissue of
sheep |
| | Musumeci et
al (84) | Lung tissue of
sheep |
| | Musumeci et
al (92) | Lung tissue of
sheep |
| | Musumeci et
al (85) | Lung tissue of
sheep |
| | Loreto et al
(86) | Lung and lymph
nodes of sheep |
| | Rapisarda et
al (93) | Tracheobronchial
lymph nodes of sheep |
| | Ledda et al
(87) | Tracheobronchial
lymph nodes of sheep |
| | Angelico et
al (94) | Human MM
tissue |
| | Caltabiano et
al (44) | Human MM
tissue |
3. In vitro studies concerned with MM
due to FE exposure
Various studies have been conducted to investigate
the effects of FE fibers on several cell lines which are commonly
used to evaluate the cytotoxicity of various silica dusts (53): A549 (human pulmonary epithelial
cancer cells), MeT-5A (human pleural mesothelial cells), J774
(mouse alveolar monocyte-macrophage cells), JU77 (human MM cells),
and human lung fibroblasts. Epithelial cells are involved in
proinflammatory effects and they are the cells of origin of
bronchogenic carcinoma. In addition, the transformation of
mesothelial cells leads to mesothelioma, thus they are suitable to
determine the direct effects of fibers. Alveolar macrophages are
the first defense mechanism against particulates and fibers
entering the lower respiratory tract (54); and fibroblasts represents a cell type
of central regulatory potential in lung diseases (55).
Prismatic vs. fibrous FE
Several studies (12,56) have
reported the cellular effects of prismatic and fibrous FE by using
the A549 cell line. These cells, upon contact with prismatic FE,
develop actin-rich protrusions from the plasma membrane, namely
ruffles and filopodia, that allow the capture and internalization
of material into the cytoplasm of epithelial cells. This
phagocytic-like behavior of cells exposed to prismatic FE occurs
only after the cells have reached the confluent state. The
organization of the actin cytoskeleton, which represents one of the
key target for a huge number of toxicants, remains well organized
both in control and in treated A549 cells. In contrast with the
results obtained with prismatic FE (12), the fibrous mineral provokes dramatic
changes in the actin network of A549 cells (56). In particular, the actin stress fibers
completely disappeared and actin-rich membrane ruffles arise from
the cell surface following FE fiber exposure. In addition to these
effects, the results obtained show that fibrous FE promote
multinucleation, cell spreading and a dramatic increase in cell
size, but without interfering with the passage of the resulting
multinucleated cells through the cell cycle and without condemning
cells to death.
Cytokines and growth factors derived from alveolar
macrophages are implicated as mediators of asbestos-induced
patho-physiological responses (57).
Indeed, inflammation also characterizes the response of epithelial
cells to external danger, which produce an array of mediators
transmitting cellular signals (58).
Interleukin (IL)-6 and IL-8 concentrations, that are respectively a
multifunctional cytokine with immunoregulatory and proinflammatory
effects (56) and a chemotactic
cytokine involved in the recruitment of polymorphonuclear
granulocytes to the site of injury (59), have been determined by Travaglione
et al (12,56) after treatment of A549 cells with
prismatic and fibrous FE. The results revealed that although
prismatic FE interferred with cell physiology, by reducing the
proliferation rate and increasing the release of the
proinflammatory cytokine IL-6, this did not perturb the cell cycle
and there was no evidence of any particular effects correlated to
cellular transformation (12). On
the contrary, in the case of fibrous FE exposure, an increase in
both IL-6 and IL-8 secretion in the surnatant, in a time
dependent-manner, was demonstrated (56). Cytokines, produced by epithelial
cells in response to a cellular damage, activate neutrophils and
macrophages which accumulate in the injured area. The recruited
cells then produce ROS (60) and
additional cytokines in an attempt to remove the unsafe agent. With
chronic inflammation, tissue fibrosis can occur concomitantly with
an enhanced risk of cancer development, arising from enhanced ROS
production leading to DNA mutations and enhanced signal
transduction that may lead to activation of oncogenes.
Therefore, these research studies, which
demonstrated a pro-mesotheliomatogenic effect of fibrous but not
prismatic FE in lung epithelial cells, highlight a differential
cell response ensued by prismatic or fibrous FE.
Fibrous FE vs. crocidolite and
tremolite
In lung epithelial cancer A549 cells, the behavior
of FE fibers is similar to crocidolite (56), whose link with chronic inflammation
and lung cancer is renowned (61,62).
Fibrous FE causes actin reorganization and multinucleation
accompanied by a significant increase in cell size. While these
effects became evident after 48 h of FE exposure, in the case of
crocidolite exposure the effects are observable at 24 h (56). Loreto et al (63) demonstrated that also FE 27 (70% of
Fe3+), FE 19 (50% Fe3+) and tremolite fibers
are able to promote the formation of multinucleated cells, but the
phenomenom is greater in cells following crocidolite exposure
(56,63).
Furthermore, the incubation with fibrous FE and
crocidolite causes a comparable decrease in the number of viable
A549 cells (56), J774 cells and
human lung fibroblasts (64). Pӓӓkkӧ
et al (65) associated the
observed reduction in A549 growth to the induction of the apoptotic
process. The results reported that crocidolite is able to cause
apoptosis in A549 cells, associating this phenomenom to the nuclear
accumulation of the p53 protein, a transcription factor mainly
involved in cell cycle arrest and apoptosis after DNA damage. In
contrast, Travaglione et al (56) demonstrated that both fibrous FE and
crocidolite were not able to influence the expression of p53 and of
pro-Bax and anti-apoptotic Bcl-2 and Bcl-XL proteins.
Furthermore, neither fibrous FE nor crocidolite were able to induce
alterations in the passage of A549 cells throughout the cell cycle;
the percentages of cells in the G0/G1, S, G2/M phases did not vary
between the control and cells exposed to the fibers.
In order to investigate the proinflammatory
potential of FE 19 and FE 27, tremolite, and crocidolite fibers,
the secretion of IL-8(56), IL-6,
IL-1β and TNF-α (63) were
determined in A549 cells. All fibers increased the release of
proinflammatory cytokines in a time-dependent manner, but
crocidolite was found to promote a more consistent secretion of the
analyzed cytokines (59,63); on the contrary, tremolite induced a
reduced release of the same (63).
Among cytokines, it has been demonstrated that IL-1β and tumor
necrosis factor (TNF)-α act through the phosphoinositide-specific
phospholipase C (PLC) pathway to activate protein kinase C (PKC)
(66), essential for many cellular
functions such as the processes of secretion, differentiation,
proliferation and cell growth (67).
However, it is known that asbestos stimulates ROS generation by
interaction with cellular membranes or through protein tyrosine
kinase (PTK), PLC and PKC pathway activation in a dose-response
manner (68). Loreto et al
(63) aimed to ascertain whether
exposure to FE 27 and FE 19 fibers may induce cytokine increase
related to PLC activation in A549 cells comparing these effects
with those of tremolite and crocidolite. The results revealed that
all fibers induced PLC expression in lung epithelial cells
following exposure to 50 µg/ml for 48 h but with a different level
of expression induced by each fiber. Tremolite showed the highest
level of expression of isoform PLC-β1 while FE 19 showed the
highest level of expression of isoform PLC-γ1. FE 27 showed values
of expression of both the comparable isoforms, while crocidolite
fibers induced lower expression of PLC. The possible induction of
oxidative stress was also examined by Cardile et al
(64) by evaluating the
intracellular ROS production, the amount of nitrite/nitrate and the
expression of inducible nitric oxide synthase (iNOS). In this case,
the cell lines exposed to FE and crocidolite included A549, J774
and human lung fibroblasts. The results indicated that the increase
in ROS generation was directly proportional to fiber concentration
and exposure time. Moreover, in all experimental cultures,
NO• synthesis and iNOS expression increased after
crocidolite fiber but not after FE fiber exposure. Cell
cytotoxicity was also evaluated assessing lactic dehydrogenase
(LDH) release in A549, J774, human lung fibroblasts exposed to FE
and crocidolite. The presence of LDH in culture medium is a marker
of membrane breakdown. The results revealed that LDH release was
significantly increased when cell cultures, in particular A549
cells, were exposed to 50 µg/ml FE or crocidolite for at least 48
h, or the when the fiber concentration was increased to 100 µg/ml
for 24 h (64).
The sensitivity to mineral fibers of A549 cells has
also been compared with pleural mesothelial MeT-5A cells (54,69). The
data demonstrated the critical role of epithelial and mesothelial
cross-talk in FE fiber exposure and that these are able to induce
functional modifications in a variety of parameters with crucial
roles in cell cycle control, cancer development and progression.
Pugnaloni et al (54)
investigated the distribution of polymerized actin in A549 and
MeT-5A cells exposed to FE fibers. Furthermore, the effects of FE
exposure on the synthesis of vascular endothelial growth factor
(VEGF), β-catenin (54) and
retinoblastoma (Rb) (69), three
critical steps of epithelial cell activation pathways, have been
investigated. The results showed greater viability in A549 than in
MeT-5A cells exposed to FE fibers (54,69) with
consequent actin staining more irregular and granular than in the
respective controls, suggesting that FE induces a dyregulated
assembly of actin (54). The
pathogenicity of FE fibers was demonstrated by the almost total
arrest of cell movement at 48 h in both cell lines and by the
flocculation of F-actin molecules. After a 48-h incubation, VEGF
and β-catenin expression were evident in both cell lines (54), suggesting that FE fibers do not exert
a primary toxic action inducing rapid cell death, but induce an
abnormal cellular status with upregulated cell activities and a
risk of cell transformation (54,70-72).
In contrast to expectations, A549 and MeT-5A cells exposed to FE 19
fibers exhibited no change in Rb level, but overexpressed
phospho-retinoblastoma (pRb) (69).
The initial status of Rb changes with the cell cycle and it is
regulated by the activity of cyclins D1, D2, D3 and CDK4 or CDK6
complexes in mid-G1 phase (73). Rb
promotes cell cycle arrest and a return to the G1 phase; it has a
central role in most instances of apoptosis, while pRb functions as
a checkpoint in the G1 phase promoting cell growth. Since cyclin D1
is a sensor of cell division signals, it has been evaluated whether
its expression is correlated with pRb expression; indeed, there was
found a positive relationship in a dose-response manner in both
cell lines (69). Furthermore, FE
reduced the p27 expression both in A549 cells (69) and in MeT-5A cells (69,74), a
tumor-suppressor gene due to its function as cell cycle regulator,
that in cancer is often inactivated (75). The downregulation of p27 is
associated with stathmin-1 upregulation in cancer, conferring an
aggressive phenotype to cancer cells (74). In addition, fibulin-3 (Fb-3)
overexpression may be responsible for the malignant transformation
of MeT-5A cells after exposure to FE fibers (74). Fb-3 overexpression reflects a
defensive response of the tissues after exogenous stimuli as FE
exposure (76).
Functional in vitro experiments performed on
MeT-5A and JU77 cells have been carried out in order to test the
carcinogenetic effects and epigenetic modulation induced by FE
exposure (77). The results showed
that MeT-5A cells were more sensitive to FE fibers compared to JU77
tumor cells. The in silico analyses revealed a set of miRNAs
strictly involved in MM and these have been used as in vitro
experimental targets. The in vitro results showed that the
expression levels of hsa-miR-323a-3p vared significantly in both
supernatant- and cell-derived miRNAs derived from treated and
untreated cells. Secreted and cellular hsa-miR-101-3p in MeT-5A
cells exposed to FE fibers and JU77 cells showed different trends
of expression. In regards to hsa-miR-20b-5p, there was no
differential expression between secreted and cellular
hsa-miR-20b-5p. This miRNA has shown a significant upregulation in
JU77 cells vs. control and treated MeT-5A. Certainly, translational
analyses will be performed on a subset of patients chronically
exposed to FE fibers to further verify the clinical role of such
miRNAs in high-risk individuals and their possible use as
biomarkers of FE exposure or MM early onset (77).
The monocyte-macrophage J774 cells are more
sensitive to FE than MeT-5A cells, suggesting that the primary site
of the inflammatory response induced by mineral fibers could be the
macrophage rather than the lung epithelium (78). The greatest sensitivity has been
demonstrated in terms of heat shock protein 70 (Hsp70) induction,
that was found to stimulate the formation of ROS and
NO•. The form 19 was found to have a markedly strong
effect on NO• biosynthesis while the form 27 had a
stimulatory effect on ROS generation in J774 cells, in contrast to
MeT-5A cells. In contrast, detection of LDH release, which is a
marker of cell necrosis, was found to be amplified in MeT-5A cells
compared to J774 cell line (60,64,78). In
general, tremolite is less effective than FE in producing more
biological alterations, while it is inactive at 5 µg/ml. In
opposition, at higher concentrations FE 19 is stronger than the
other particulate (78).
Alveolar macrophages have an important role in the
fibrotic process involved in silicosis and in other lung diseases
(79). They are mediators in the
interaction between inhaled particulates and different types of
cells, by the release of a variety of inflammatory and
growth-mediating factors (80).
Cyclooxygenase-2 (COX-2) catalyses the conversion of arachinodic
acid to prostaglandins (PGs) and it is mainly induced in response
to proinflammatory stimuli, cytokines, growth factors and mitogens.
It is known that PGs have an important role in cancer pathogenesis
(81); therefore, Pugnaloni et
al (54) demonstrated a
time-dependent COX-2 overexpression and a PGE2 increase
in monocyte-macrophage J774 cells exposed to FE. PEG2
derived from COX-2 was found to be involved in solid tumor
pathogenesis through inhibition of apoptosis, facilitation of tumor
cell invasiveness and promotion of angiogenesis (54).
4. In vivo studies concerned with MM
due to FE exposure
It is important to underline that FE fibers were
identified in the lungs of a housewife of Biancavilla, who died
subsequently to a diagnosis of MM (45), in lung tissue (43,82-86)
and lymph nodes (86,87) of sheep living in the Biancavilla
area. Indeed, several studies have been performed to evaluate the
relationship between FE and MM and to prove the biopersistence of
these fibers in tissue.
Induced exposure of prismatic vs.
fibrous FE
Different authors (36,88) have
tested the same concentration of two mineral forms, prismatic and
fibrous FE, with two administration methods, single intraperitoneal
and intrapleural injection, on groups of Sprague-Dawley rats, to
acquire more information on the potential relationship between
exposure to FE and MM. The results are concordant and in line with
previous preliminary data (64), and
there is evident confirmation regarding a mesotheliomatogenic
potential of FE fibers. In contrast, prismatic FE failed to induce
a mesotheliomatogenic response in the totality of the animals, in
accordance to the controls (36,88).
The intraperitoneal injection of fibrous FE caused
effects much stronger than those observable by intrapleural
administration (36,88). In particular, 82.5% of the deceased
rats treated by intraperitoneal injection died because of
mesothelioma; only 14.3% of the deceased rats treated by
intrapleural injection died due to mesothelioma induced by fibrous
FE. The peritoneal tumors involved the abdominal cavity, with
whitish and yellowish tissue on the surface of all the organs;
furthermore a serosal effusion was present in almost all cases.
Instead, the pleural tumors involved the visceral and/or parietal
pleura and in 80% of the cases the diaphragm was largely involved
with subsequent extension of the cancer into the peritoneal cavity
(36).
Prismatic FE did not provoke mesothelioma in animal
models. On the contrary, fibrous FE was found to cause a strong
mesotheliomatogenic effect on the peritoneum and a milder extent on
the pleura.
5. Ex vivo studies concerned with MM
due to FE exposure
Environmental exposure of fibrous
FE
Sheep lung is comparable to human lung in
architecture, volume, and respiratory parameters (89); therefore, it is a suitable model for
toxicological studies concerning exposure to environmental
pollutants (89), such as asbestos
(20,90,91) or
FE fibers (43,83-87,92).
One of the first experimental study (43) that analyzed sheep exposed to FE
fibers present in the surrounding environment of Biancavilla,
demonstrated that the first pathological event seems to involve the
alveolar epithelium, resulting in classic honeycombing (43,83), and
subsequently the interstitial matrix (43,83,84).
Matrix metalloproteinase (MMP)-13 is mainly overexpressed in
fibroblasts and epithelial cells, while immunopositivity of
TNF-related apoptosis-inducing ligand (TRAIL) and its receptor
death receptor 5 (DR5) are detected on alveolar surfaces and in the
vascular stroma. The triggering event at the level of type I
pneumocytes seems to damage the cytoplasmic membrane, resulting in
loss of cell elements and exposure of underlying capillaries, and
eventually in a series of reactions including macrophage
activation, possible release of growth factors, metaplasic
reconstruction of lung alveoli, and fibrosis (43). Loreto et al (83) demonstrated epithelial and
interstitial Bax overexpression and negative Bcl-2
immunoexpression. pRb overexpression was also detected in
FE-exposed sheep lung, in particular in alveolar epithelium and the
interstitium, while Rb expression was absent (84). Immunopositivity for TRAIL and MMP-13
receptor (43), the changes in Bax
and Bcl-2(83), and the altered
balance between Rb and pRb expression (84) can be considered a programmed response
to protect the organism against uncontrolled cell proliferation,
suggesting that apoptosis may be activated by FE fibers. A
significant increase in the expression of CD68-positive
macrophages, tryptase-positive mast cells, as well as a significant
increase in microvascular density evaluated as CD31-positive areas
in lung tissue of sheep exposed to FE fibers have been
demonstrated. These data confirm the important role played by tumor
microenvironment components in favor of angiogenesis in MM induced
by FE exposure (85). Musumeci et
al (92) investigated
N-cadherin, ADAM-10 and aquaporin-1 (AQP1) expression in the lung
tissue of sheep exposed to FE fibers, showing different patterns of
immunolabeling. N-cadherin and ADAM-10 were more expressed in
FE-exposed lung tissue, when compared with the control. On the
contrary, AQP1 was more highly expressed in non-exposed lung
tissue. These results suggest that N-cadherin, ADAM-10 and AQP1 are
probably involved in different pathological processes induced by FE
fiber exposure. The cellular and molecular toxicity mechanisms and
the cellular response to FE fibers are still not well known, but
these results highlight that molecules involved in carcinogenesis
and in the inflammatory process participate in the network of
events induced by exposure to FE fibers.
Loreto et al (86) demonstrated overexpression of
MacroH2A.1, at the protein level, in lung epithelial cells and in
lymph nodes of sheep exposed to FE fibers. The data suggest an
involvement of MacroH2A.1 in the cellular response triggered by
direct exposure to FE. The immunoreactions were detected in the
areas where fibers were embedded and localized, in the lung, to the
pulmonary and bronchial epithelium and not to the fibrotic
interstitium. This significant research (86) seems to show a clear association
between exposure to FE fibers and MacroH2A.1 expression view to
identifying, in the future, a targeting epigenetics for cancer
therapy.
Several studies have examined the lymph node
draining pulmonary lobes of sheep grazing around Biancavilla
(87,93). The results show a greater size of
lymph nodes with signs of anthracosis. At the paracortical level,
they show lymph-follicle hyperplasia with wide reactive corea and
several macrophages containing grey-brownish particulate
interspersed with elements with a fibril structure, forming
nodules. Similar findings were detected in some peribronchiolar
areas of the lung parenchyma. The FE fiber dimensions found in
digested lymph nodes of sheep were similar to those found in the
lung of a housewife from Biancavilla who died of MM as described by
Paoletti et al (45). Indeed,
sheep can be a biological indicator of environmental pollution by
FE fibers, measuring these fibers in lymph node draining pulmonary
lobes (87,93).
Several immunohistochemical investigations were
conducted to demonstrate the implication of different compounds in
MM due to FE fibers exposure and to investigate their potential
role as diagnostic and prognostic markers. Angelico et al
(94) demonstrated the prognostic
role of AQP1 in FE-induced MM. In fact, the immunohistochemical
overexpression of AQP1 was found to be associated with an increased
median overall survival. Caltabiano et al (44) found high immunoexpression of Fb-3 in
neoplastic cells with nuclear and cytoplasmic localization,
demonstrating the implication of Fb-3 in MM due to FE exposure.
Fb-3 could therefore have a potential role as a diagnostic and
prognostic marker.
6. Discussion
The prevention of pathologies related to exposure
to carcinogenic fibers such as asbestos and fluoro-edenite (FE),
also includes the reduction of these fibers in the environment.
Generally, this can be achieved in three ways: Reclamation,
encapsulation and confinement. These interventions tend to
eliminate airborne fibers to avoid exposure that can cause various
diseases including cancer.
The observation of a significant incidence of MM,
subsequently linked to the inhalation of FE fibers of Biancavilla,
has been reported by epidemiological studies (45-48).
Previous cross-sectional studies conducted on subjects exposed to
FE fibers confirm the in vitro, in vivo and ex
vivo data. In fact, it has been demonstrated that exposure to
FE fibers may induce autoimmunity (95,96), and
the involvement of the inflammosome (41).
This malignant cancer is a highly aggressive
neoplasm of the serosal membranes lining the pleural cavity
(97). Only 5% of MM patients are
diagnosed at an early stage (98)
and the median survival is approximately 6-12 months (97,99).
Moreover, current treatment for MM, which is based on surgery and
standard chemotherapy, has a modest effect on the overall survival
(OS), which remains dismal (4).
The diagnosis of MM is always challenging as MM may
appear in patients up to 30-40 years after exposure to carcinogenic
fibers; the clinical and imaging signs of MM are non-specific; and
a definitive diagnosis, which relies on histology, can sometimes be
very difficult to achieve, even with the use of
immunohistochemistry. To date, no single marker or panel of soluble
biomarkers is available for a clear diagnosis of MM (100).
Many biomarkers have been proposed for screening
and diagnosis of MM in subjects exposed, such as calretinin,
cytokeratin 5 (CK5), podoplanin, mesothelin, osteopontin,
hyaluronic acid, Fb-3(44), VEGF
(101), AQP1(94), high mobility group box 1 (HMGB-1)
(102), and MacroH2A.1(86). Mesothelin is the only Food and Drug
Administration (FDA)-approved biomarker for MM (103-105),
but with limitations (106). In
fact, the poor sensitivity of mesothelin clearly limits the added
value to the diagnosis of MM (100).
Some studies have been conducted to understand the
link between common genetic variations in the molecular pathways
and cancer risk with the final goal to develop novel therapeutic
targets. Lim et al (107)
reported mutations in SMO and SUFU and a novel multi-exonic
deletion in PTCH1 in MM cell lines and tumors. These data suggest
that aberrant activation of the Hedgehog (HH) signaling in MM is
unlikely to be driven by mutations in the majority of tumors but
instead activated through autocrine signaling (107,108).
This pathway may represent a novel therapeutic target in MM for
recently developed HH pathway inhibitors.
Several studies have demonstrated that microRNAs
(miRNAs) may be used as valuable non-invasive diagnostic and
prognostic biomarkers for various human diseases, including cancers
(42,109,110).
In the clinical setting, circulating cell-free miRNAs and fecal
miRNAs are the main forms of RNA used as diagnostic biomarkers
(111). In particular, a recent
review of the literature by Ledda et al (112) indicates a list of miRNAs
potentially involved in MM. Potential miRNA biomarkers for this
malignant neoplasm include the following: miRNA-126-3p,
miRNA-625-3p, miRNA-103a-3p, miRNA-16-5p, miRNA-143-3p,
miRNA-145-5p, miRNA-192-5p, miRNA-193a-3p, miRNA-200b-3p,
miRNA-203a-3p, and miRNA-652-3p. The scientific community has
revealed that several miRNAs are involved in deregulation and in
all molecular mechanisms associated with MM development (113,114)
and constantly updates the miRNAs which can be associated with MM
early diagnosis and prognosis.
In recent studies, several bioinformatics
approaches to identify selected miRNAs highly deregulated in cancer
samples when compared with normal control have been developed
(77,115-118).
The in silico study of the expression of certain miRNAs
represents an effort in the field of biomarker discovery because in
this way it is possible to analyze the data coming from multiple
studies of miRNA profiling; in this way it is possible to have a
large series of samples useful to obtain truthful expression data
concerning miRNAs with a potential diagnostic and prognostic role
in cancer. In addition, the development of new high-sensitivity
technologies and the analysis of liquid biopsy samples and
circulating tumor DNA are paving the way to new non-invasive
validation studies aimed to discover new promising diagnostic and
prognostic biomarkers for several pathologies, including MM
(119-121).
Therefore, future studies will be conducted to
understand the link between common genetic variations in the
molecular pathways and cancer risk with the final goal to develop
novel therapeutic targets. Research is needed in order to
computationally select putative miRNAs involved in the development
and progression of lung cancer or MM and to be validated in
correspective in vitro tumor models and in a subset of
patients chronically exposed to FE. It could also be particularly
helpful to study and subsequently use a combination of several
protein and molecular markers to improve diagnostic accuracy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All information included in this review has been
documented by relevant references.
Authors' contributions
Conceptualization of the review was accomplished by
VF and CL. The research methodology was designed and conducted by
VF, EV and GB. Validation of the research data was conducted by VF
and CL. Formal analysis was carried out by VF. Investigation of the
data was carried out by VF, EV and GB. Data curation was conducted
by VF, MPH and SC. Writing-original draft preparation was carried
out by VF and CL. Writing-review and editing was accomplished by
VF, MPH, SC and AS. Supervision was conducted by CL. All authors
read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors report that they have no competing
interests.
References
|
1
|
Hansen J, De Klerk NH, Eccles JL, Musk AW
and Hobbs MS: Malignant mesothelioma after environmental exposure
to blue asbestos. Int J Cancer. 54:578–581. 1993.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kanarek MS: Mesothelioma from chrysotile
asbestos: Update. Ann Epidemiol. 21:688–697. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bayram M, Dongel I, Bakan ND, Yalççn H,
Cevit R, Dumortier P and Nemery B: High risk of malignant
mesothelioma and pleural plaques in subjects born close to
ophiolites. Chest. 143:164–171. 2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Carbone M, Kanodia S, Chao A, Miller A,
Wali A, Weissman D, Adjei A, Baumann F, Boffetta P, Buck B, et al:
Consensus report of the 2015 weinman international conference on
mesothelioma. J Thorac Oncol. 11:1246–1262. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Case BW, Abraham JL, Meeker G, Pooley FD
and Pinkerton KE: Applying definitions of ‘asbestos’ to
environmental and ‘low-dose’ exposure levels and health effects,
particularly malignant mesothelioma. J Toxicol Environ Health B
Crit Rev. 14:3–39. 2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Huang SX, Jaurand MC, Kamp DW, Whysner J
and Hei TK: Role of mutagenicity in asbestos fiber-induced
carcinogenicity and other diseases. J Toxicol Environ Health B Crit
Rev. 14:179–245. 2011.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liu G, Cheresh P and Kamp DW: Molecular
basis of asbestos-induced lung disease. Annu Rev Pathol. 8:161–187.
2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Ledda C, Pomara C, Bracci M, Mangano D,
Ricceri V, Musumeci A, Ferrante M, Musumeci G, Loreto C, Fenga C,
et al: Natural carcinogenic fiber and pleural plaques assessment in
a general population: A cross-sectional study. Environ Res.
150:23–29. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Falzone L, Marconi A, Loreto C, Franco S,
Spandidos DA and Libra M: Occupational exposure to carcinogens:
Benzene, pesticides and fibers (Review). Mol Med Rep. 14:4467–4474.
2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Gangemi S, Rapisarda V, Minciullo PL, Di
Pasquale G, Lombardo G, Valentino M and Fenga C: Circulating levels
of interleukin-18 in asbestos-exposed workers. Toxicol Ind Health.
21:125–129. 2005.
|
|
11
|
Aitio A, Cantor KP, Attfield MD, Demers
PA, Fowler BA, Grandjean P, Fubini B, Hartwig A, Gérin M, et al: A
review of human carcinogens: Arsenic, metals, fibres and dusts. Vol
100 C. IARC, Lyon, pp219-224, 2012.
|
|
12
|
Travaglione S, Bruni B, Falzano I,
Paoletti I and Fiorentini C: Effects of the new-identified
amphibole FE in lung epithelial cells. Toxicol In Vitro.
17:547–552. 2003.
|
|
13
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Admit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E
and Ben-Neriah Y: NF-kappaB function as a tumor promoter in
inflammation-associated cancer. Nature. 431:461–466.
2004.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Tokokuni S: Mechanisms of asbestos-induced
carcinogenesis. Nagoya J Med Sci. 71:1–10. 2009.PubMed/NCBI
|
|
15
|
Koskinen K, Rinne JP, Zitting A,
Tossavainen A, Kivekäs J, Reijula K, Roto P and Huuskonen MS:
Screening for asbestos-induced diseases in Finland. Am J Ind Med.
30:241–251. 1996.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pan XL, Day HW, Wang W, Beckett LA and
Schenker MB: Residential proximity to naturally occurring asbestos
and mesothelioma risk in California. Am J Respir Crit Care Med.
172:1019–1025. 2005.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Luo S, Liu X, Mu S, Tsai SP and Wen CP:
Asbestos related diseases from environmental exposure to
crocidolite in Da-yao, China. I. Review of exposure and
epidemiological data. Occup Environ Med. 60:35–41. 2003.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Baumann F, Maurizot P, Mangeas M, Ambrosi
JP, Douwes J and Robineau B: Pleural mesothelioma in New Caledonia:
Associations with environmental risk factors. Environ Health
Perspect. 119:695–700. 2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Rey F, Boutin C, Steinbauer J, Viallat JR,
Alessandroni P, Jutisz P, Di Giambattista D, Billon-Galland MA,
Hereng P, Dumortier P, et al: Environmental pleural plaques in an
asbestos exposed population of northeast Corsica. Eur Respir J.
6:978–982. 1993.PubMed/NCBI
|
|
20
|
McConnochie K, Simonato L, Mavrides P,
Christofides P, Pooley FD and Wagner JC: Mesothelioma in Cyprus:
The role of tremolite. Thorax. 42:342–347. 1987.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Constantopoulos SH: Environmental
mesothelioma associated with tremolite asbestos: Lessons from the
experiences of Turkey, Greece, Corsica, New Caledonia and Cyprus.
Regul Toxicol Pharmacol. 52 (1 Suppl):S110–S115. 2008.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Baris YI, Artvinli M, Sahin AA, Sebastien
P and Gaudichet A: Diffuse lung fibrosis due fibrous zeolite
(erionite) exposure. Eur J Respir Dis. 70:122–125. 1987.PubMed/NCBI
|
|
23
|
Yazicioglu S, Ilçayto R, Balci K, Sayli BS
and Yorulmaz B: Pļeural calcification, pleural mesotheliomas, and
bronchial cancers caused by tremolite dust. Thorax. 35:564–569.
1980.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dumortier P, Coplü L, de Maertelaer V,
Emri S, Baris I and De Vuyst P: Assessment of environmental
asbestos exposure in Turkey by bronchoalveolar lavage. Am J Respir
Crit Care Med. 158:1815–1824. 1998.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Senyigit A, Dalgic A, Kavak O and
Tanrikulu AC: Determination of environmental exposure to asbestos
(tremolite) and mesothelioma risks in the southeastern region of
Turkey. Arch Environ Health. 59:658–662. 2004.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Metintas M, Metintas S, Hillerdal G, Ucgun
I, Erginel S, Alatas F and Yildirim H: Non malignant pleural
lesions due to environmental exposure to asbestos: A field-based,
cross-sectional study. Eur Respir J. 26:875–880. 2005.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Döngel I, Bayram M, Bakan ND, Yalçın H and
Gültürk S: Is living close to ophiolites related to asbestos
related diseases? Cross-sectional study. Respir Med. 107:870–874.
2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kliment CR, Clemens K and Oury TD: North
American erionite-associated mesothelioma with pleural plaques and
pulmonary fibrosis: A case report. Int J Clin Exp Pathol.
2:407–410. 2009.PubMed/NCBI
|
|
29
|
Baumann F, Buck BJ, Metcalf RV, McLaurin
BT, Merkler DJ and Carbone M: The presence of asbestos in the
natural environment is likely related to mesothelioma in young
individuals and women from southern Nevada. J Thorac Oncol.
10:731–737. 2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Konen T, Johnson JE, Lindgren P and
Williams A: Cancer incidence and mortality associated with
non-occupational and low dose exposure to Libby vermiculite in
Minnesota. Environ Res. 175:449–456. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Baur X: Review on the adverse health
effects of asbestiform antigorite, a non-regulated asbestiform
serpentine mineral. Am J Ind Med. 61:625–630. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Cardile V, Lombardo L, Belluso E, Panico
A, Capella S and Balazy M: Toxicity and carcinogenicity mechanisms
of fibrous antigorite. Int J Environ Res Public Health. 4:1–9.
2007.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Groppo C and Compagnoni R: Ubiquitous
fibrous antigorite veins from the Lanzo Ultramafic Massif, Internal
Western Alps (Italy): Characterisation and genetic conditions. Per
Mineral. 76:169–181. 2007.
|
|
34
|
FitzGerald J, Eggleton R and Keeling J:
Antigorite from Rowland flat, South Australia: Asbestiform
character. Eur J Mineral. 22:525–533. 2010.
|
|
35
|
Fitzgerald SM and Harty EA: Antigorite: Is
it the forgotten asbestos? Prof Saf. 59:43–48. 2014.
|
|
36
|
Soffritti M, Minardi F, Bua L, Degli
Esposti D and Belpoggi F: First experimental evidence of peritoneal
and pleural mesotheliomas induced by FE fibres present in Etnean
volcanic material from Biancavilla (Sicily, Italy). Eur J Oncol.
9:169–715. 2004.
|
|
37
|
Gianfagna A and Oberti R: Fluoro-edenite
from Biancavilla (Catania, Sicily, Italy): Crystal chemistry of a
new amphibole end-member. Am Mineral. 86:1489–1493. 2001.
|
|
38
|
Biggeri A, Pasetto R, Belli S, Bruno C, Di
Maria G, Mastrantonio M, Trinca S, Uccelli R and Comba P: Mortality
from chronic obstructive pulmonary disease and pleural mesothelioma
in an area contaminated by natural fiber (fluoro-edenite). Scand J
Work Environ Health. 30:249–252. 2004.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Comba P, Gianfagna A and Paoletti L:
Pleural mesothelioma cases in Biancavilla are related to a new
fluoro-edenite fibrous amphibole. Arch Environ Health. 58:229–232.
2003.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Gianfagna A, Ballirano P, Bellatreccia F,
Bruni B, Paoletti L and Oberti R: Characterization of amphibole
fibres linked to mesothelioma in the area of Biancavilla, Eastern
Sicily, Italy. Mineral Mag. 67:1221–1229. 2003.
|
|
41
|
Ledda C, Costa C, Matera S, Puglisi B,
Costanzo V, Bracci M, Fenga C, Rapisarda V and Loreto C:
Immunomodulatory effects in workers exposed to naturally occurring
asbestos fibers. Mol Med Rep. 15:3372–3378. 2017.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Ledda C and Rapisarda V: Malignant pleural
mesothelioma: The need to move from research to clinical practice.
Arch Med Res. 47(407)2016.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Martinez G, Loreto C, Rapisarda V,
Musumeci G, Valentino M and Carnazza ML: Effects of exposure to
fluoro-edenite fibre pollution on the respiratory system: An in
vivo model. Histol Histopathol. 21:595–601. 2006.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Caltabiano R, Loreto C, Vitale E, Matera
S, Miozzi E, Migliore M, Angelico G, Tumino R, Ledda C and
Rapisarda V: Fibulin-3 immunoexpression in malignant mesothelioma
due to fluoro-edenite: A preliminary report. Future Oncol. 14 (6
Suppl):S53–S57. 2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Paoletti L, Batisti D, Bruno C, Di Paola
M, Gianfagna A, Mastrantonio M, Nesti M and Comba P: Unusually high
incidence of malignant pleural mesothelioma in a town of eastern
Sicily: An epidemiological and environmental study. Arch Environ
Health. 55:392–398. 2000.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Di Paola M, Mastrantonio M, Carboni M,
Belli S, Grignoli M, Comba P and Nesti M: Mortality from malignant
pleural neoplasms in Italy in the years 1988-1992. Rapporti
ISTISAN, 96/40. Istituto Superiore di Sanità, Rome, 1996.
|
|
47
|
Fazzo L, De Santis M, Minelli G, Bruno C,
Zona A, Marinaccio A, Conti S and Comba P: Pleural mesothelioma
mortality and asbestos exposure mapping in Italy. Am J Industr Med.
55:11–24. 2012.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Fazzo L, Minelli G, De Santis M, Bruno C,
Zona A, Marinaccio A, Conti S, Pirastu R and Comba P: Mesothelioma
mortality surveillance and asbestos exposure tracking in Italy. Ann
Ist Super Sanita. 48:300–310. 2012.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Rapisarda V, Ledda C, Ricceri V, Arena F,
Musumeci A, Marconi A, Fago L, Bracci M, Santarelli L and Ferrante
M: Detection of pleural plaques in workers exposed to inhalation of
natural fluoro-edenite fibres. Oncol Lett. 9:2046–2052.
2015.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ledda C, Loreto C, Matera S, Massimino N,
Cannizzaro E, Musumeci A, Migliore M, Fenga C, Pomara C and
Rapisarda V: Early effects of fluoro-edenite: Correlation between
IL-18 serum levels and pleural and parenchymal abnormalities.
Future Oncol. 12 (23 Suppl):S59–S62. 2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Grosse Y, Loomis D, Guyton KZ,
Lauby-Secretan B, El Ghissassi F, Bouvard V, Benbrahim-Tallaa L,
Guha N, Scoccianti C, Mattock H and Straif K: Carcinogenicity of
fluoro-edenite, silicon carbide fibres and whiskers, and carbon
nanotubes. Lancet Oncol. 15:1427–1428. 2014.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Miller SA: PICO worksheet and search
strategy. National Center for Dental Hygiene Research, 2001.
|
|
53
|
Fenoglio I, Croce A, Di Renzo F, Tiozzo R
and Fubini B: Pure-silica zeolites (Porosils) as model solids for
the evaluation of the physicochemical features determining silica
toxicity to macrophages. Chem Res Toxicol. 13:489–500.
2000.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Pugnaloni A, Lucarini G, Giantomassi F,
Lombardo L, Capella S, Belluso E, Zizzi A, Panico AM, Biagini G and
Cardile V: In vitro study of biofunctional indicators after
exposure to asbestos-like fluoro-edenite fibres. Cell Mol Biol
(Noisy-le-grand). 53 (Suppl):OL965–OL980. 2007.PubMed/NCBI
|
|
55
|
Tremblay GM, Jordana M, Gauldie J and
Särnstrand B: Fibroblasts as effector cells in fibrosis. In:
Pulmonary fibrosis. Phan SH and Thrall RS (eds). Marcel Dekker
Inc., New York, NY, pp541-577, 1995.
|
|
56
|
Travaglione S, Bruni BM, Falzano L,
Filippini P, Fabbri A, Paoletti L and Fiorentini C: Multinucleation
and pro-inflammatory cytokine release promoted by fibrous
fluoro-edenite in lung epithelial A549 cells. Toxicol in Vitro.
20:841–850. 2006.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Li XY, Lamb D and Donaldson K: The
production of TNF-alpha and IL-1-like activity by bronchoalveolar
leucocytes after intratracheal instillation of crocidolite
asbestos. Int J Exp Pathol. 74:403–410. 1993.PubMed/NCBI
|
|
58
|
Trimblin C, BeruBe K, Churg A, Driscoll K,
Gordon T, Hemenway D, Walsh E, Cummins AB, Vacek P and Mossman B:
Ambient particulate matter causes activation of the c-jun
kinase/stress-activated protein kinase cascade and DNA synthesis in
lung epithelial cells. Cancer Res. 58:4543–4547. 1998.PubMed/NCBI
|
|
59
|
Hedges S, Svensson M, Agace W and Svanborg
C: Cytokines induce an epithelial cell cytokine response. Adv Exp
Med Biol. 371A:189–193. 1995.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Cardile V, Proietti L, Panico A and
Lombardo L: Nitric oxide production in fluoro-edenite treated mouse
monocyte-macrophage cultures. Oncol Rep. 12:1209–1215.
2004.PubMed/NCBI
|
|
61
|
Hedenborg M and Klockars M: Production of
reactive oxygen metabolites induced by asbestos fibres in human
polymorphonuclear leucocytes. J Clin Pathol. 40:1189–1193.
1987.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Hamilton RF, Iyer LL and Holian A:
Asbestos induces apoptosis in human alveolar macrophages. Am J
Physiol. 271:L813–L819. 1996.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Loreto C, Carnazza ML, Cardile V, Libra M,
Lombardo L, Malaponte G, Martinez G, Musumeci G, Papa V and Cocco
L: Mineral fiber-mediated activation of phosphoinositide-specific
phospholipase c in human bronchoalveolar carcinoma-derived alveolar
epithelial A549 cells. Int J Oncol. 34:371–376. 2009.PubMed/NCBI
|
|
64
|
Cardile V, Renis M, Scifo C, Lombardo L,
Gulino R, Mancari B and Panico A: Behaviour of the new asbestos
amphibole fluor-edenite in different lung cell systems. Int J
Biochem Cell Biol. 36:849–860. 2004.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Pӓӓkkӧ P, Rämet M, Vähäkangas K, Korpela
N, Soini Y, Turunen S, Jaworska M and Gillissen A: Crocidolite
asbestos causes an induction of p53 and apoptosis in cultured A-549
lung carcinoma cells. Apoptosis. 3:203–212. 1998.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Yang CM, Luo SF, Wang CC, Chiu CT, Chien
CS, Lin CC and Hsiao LD: Tumour necrosis factor-alpha- and
interleukin-1beta-stimulated cell proliferation through activation
of mitogen-activated protein kinase in canine tracheal smooth
muscle cells. Br J Pharmac. 130:891–899. 2000.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Rana Rs and Hokin LE: Role of
phosphoinositides in trans-membrane signaling. Physiol Rev.
70:115–164. 1990.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Lim Y, Kim SH, Kim KA, Oh MW and Lee KH:
Involvement of protein kinase C, phospholipase C, and protein
tyrosine kinase pathways in oxygen radical generation by
asbestos-stimulated alveolar macrophage. Environ Health Perspect.
105 (Suppl 5):S1325–S1327. 1997.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Musumeci G, Cardile V, Fenga C, Caggia S
and Loreto C: Mineral fibre toxicity: Expression of retinoblastoma
(Rb) and phospho-retinoblastoma (pRb) protein in alveolar
epithelial and mesothelial cell lines exposed to fluoro-edenite
fibres. Cell Biol Toxicol. 27:217–225. 2011.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Beachy PA, Karhadkar SS and Berman DM:
Tissue repair and stem cell renewal in carcinogenesis. Nature.
432:324–331. 2004.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Chen S, Guttrige DC, You Z, Zhang Z,
Fribley A, Mayo MW, Kitajewski J and Wang CY: Wnt-1 signaling
inhibits apoptosis by activating beta-catenin/T cell
factor-mediated transcription. J Cell Biol. 152:87–96.
2001.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Zheng R, Yano S, Matsumori Y, Nakataki E,
Muguruma H, Yoshizumi M and Sone S: SRC tyrosine kinase inhibitor,
m475271, suppresses subcutaneous growth and production of lung
metastasis via inhibition of proliferation, invasion, and
vascularization of human lung adenocarcinoma cells. Clin Exp
Metastasis. 22:195–204. 2005.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Yamanouchi H, Furihata M, Fujita JJ,
Murakami H, Yoshinouchi T, Takahara J and Ohtsuki Y: Expression of
cyclin E and cyclin D1 in non-small cell lung cancers. Lung Cancer.
31:3–8. 2001.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Rapisarda V, Salemi R, Marconi A, Loreto
C, Graziano AC, Cardile V, Basile MS, Candido S, Falzone L,
Spandidos DA, et al: Fluoro-edenite induces fibulin-3
overexpression in non-malignant human mesothelial cells. Oncol
Lett. 12:3363–3367. 2016.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Chu IM, Hengst L and Slingerland JM: The
Cdk inhibitor p27 in human cancer: Prognostic potential and
relevance to anticancer therapy. Nat Rev Cancer. 8:253–267.
2008.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Rapisarda V, Caltabiano R, Musumeci G,
Castrogiovanni P, Ferrante M, Ledda C, Lombardo C, Graziano ACE,
Cardile V and Loreto C: Analysis of fibulin-3 after exposure to
asbestos-like fibers. Environ Res. 156:381–387. 2017.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Filetti V, Falzone L, Rapisarda V,
Caltabiano R, Graziano ACE, Ledda C and Loreto C: Modulation of
microRNA expression levels after naturally occurring asbestiform
fibers exposure as a diagnostic biomarker of mesothelial neoplastic
transformation. Ecotoxicol Environ Saf. 198(110640)2020.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Cardile V, Lombardo L, Belluso E, Panico
A, Renis M, Gianfagna A and Balazy M: Fluoro-edenite fibers induce
expression of Hsp70 and inflammatory response. Int J Environ Res
Public Health. 4:195–202. 2007.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Shukla A, Gulumian M, Hei TK, Kamp D,
Rahman Q and Mossman B: Multiple roles of oxidants in the
pathogenesis of asbestos-induced diseases. Free radical Biol Med.
34:1117–1129. 2003.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Holian A, Kelley K and Hamilton RF Jr:
Mechanisms associated with human alveolar macrophage stimulation by
particulates. Environ Health Perspect. 102 (Suppl 10):S69–S74.
1994.PubMed/NCBI View Article : Google Scholar
|
|
81
|
De Witt DL: Prostaglandin endoperoxide
synthase: Regulation of enzyme expression. Biochim Byophys Acta.
1083:121–134. 1991.PubMed/NCBI View Article : Google Scholar
|
|
82
|
DeNardo P, Bruni B, Paoletti L, Pasetto R
and Sirianni B: Pulmonary fibre burden in sheep living in the
Biancavilla area (Sicily): Preliminary results. Sci Total Environ.
325:51–58. 2004.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Loreto C, Rapisarda V, Carnazza ML,
Musumeci G, Valentino M, Fenga C and Martinez G: Fluoro-edenite
fibres induce lung cell apoptosis: An in vivo study. Histol
Histopathol. 23:319–326. 2008.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Musumeci G, Loreto C, Cardile V, Carnazza
ML and Martinez G: Immunohistochemical expression of retinoblastoma
and phospho-retinoblastoma protein in sheep lung exposed to
fluoro-edenite fibers. Anat Sci Int. 85:74–78. 2010.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Musumeci G, Loreto C, Giunta S, Rapisarda
V, Szychlinska MA, Imbesi R, Castorina A, Annese T, Castorina S,
Castrogiovanni P and Ribatti D: Angiogenesis correlates with
macrophage and mast cell infiltration in lung tissue of animals
exposed to fluoro-edenite fibers. Exp Cell Res. 346:91–98.
2016.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Loreto C, Lombardo C, Caltabiano R, Ledda
C, Hagnas M, Filetti V and Rapisarda V: An in vivo
immunohistochemical study on MacroH2A.1 in lung and lymph-node
tissues exposed to an asbestiform fiber. Curr Mol Med: Feb 20, 2020
(Epub ahead of print).
|
|
87
|
Ledda C, Loreto C, Pomara C, Rapisarda G,
Fiore M, Ferrante M, Bracci M, Santarelli L, Fenga C and Venerando
R: Sheep lymph-nodes as a biological indicator of environmental
exposure to fluoro-edenite. Environ Res. 147:97–101.
2016.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Belpoggi F, Tibaldi E, Lauriola M, Bua L,
Falcioni L, Chiozzotto D, Manservisi F, Manservigi M and Soffritti
M: The efficacy of long-term bioassays in predicting human risks:
Mesotheliomas induced by fluoro-edenite fibres present in lava
stone from the Etna volcano in Biancavilla, Italy. Eur J Oncol.
16:185–196. 2011.
|
|
89
|
Begin R, Rola-Pleszczynski M, Sirois P,
Masse S, Nadeau D and Bureau MA: Sequential analysis of the
bronchoalveolar milieu in conscious sheep. J Appl Physiol Resp
Envir Exerc Physiol. 50:665–671. 1981.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Schlesinger RB: Clearance from the
respiratory tract. Fundam Appl Toxicol. 5:435–450. 1985.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Dumortier P, Rey F, Viallat JR, Broucke I,
Boutin C and De Vuyst P: Chrysotile and tremolite asbestos fibres
in the lungs and parietal pleura of Corsican goats. Occup Env Med.
59:643–646. 2002.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Musumeci G, Loreto C, Szychlinska MA,
Imbesi R, Rapisarda V, Aiello FC, Castorina S and Castrogiovanni P:
N-Cadherin, ADAM-10 and Aquaporin 1 expression in lung tissue
exposed to fluoro-edenite fibers: An immunohistochemical study.
Histol Histopathol. 30:987–999. 2015.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Rapisarda V, Rapisarda G, Vico CD, Gobbi
L, Loreto C and Valentino M: Monitoring of fluoro-edenite fibre
pollution through the study of sheep lymph nodes as a model of a
biological indicator. Occup Environ Med. 62(656)2005.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Angelico G, Caltabiano R, Loreto C, Ieni
A, Tuccari G, Ledda C and Rapisarda V: Immunohistochemical
expression of aquaporin-1 in fluoro-edenite-induced malignant
mesothelioma: A preliminary report. Int J Mol Sci.
19(685)2018.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Ledda C, Caltabiano R, Loreto C, Cinà D,
Senia P, Musumeci A, Ricceri V, Pomara C and Rapisarda V:
Prevalence of anti-nuclear autoantibodies in subjects exposed to
natural asbestiform fibers: A cross-sectional study. J
Immunotoxicol. 15:24–28. 2018.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Rapisarda V, Loreto C, Castorina S, Romano
G, Garozzo SF, Musumeci A, Migliore M, Avola R, Cinà D, Pomara C
and Ledda C: Occupational exposure to fluoro-edenite and prevalence
of anti-nuclear autoantibodies. Future Oncol. 14 (6 Suppl):S59–S62.
2018.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Baas P, Fennell D, Kerr KM, van Schil PE,
Haas RL and Peters S: ESMO Guidelines Committee. Malignant pleural
mesothelioma: ESMO clinical practice guidelines for diagnosis,
treatment and follow-up. Ann Oncol. 26 (Suppl 5):v31–v39.
2015.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Ying S, Jiang Z, He X, Yu M, Chen R, Chen
J, Ru G, Chen Y, Chen W, Zhu L, et al: Serum HMGB1 as a potential
biomarker for patients with asbestos-related diseases. Dis Markers.
2017(5756102)2017.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Tomasson K, Gudmundsson G, Briem H and
Rafnsson V: Malignant mesothelioma incidence by nation-wide cancer
registry: A population-based study. J Occup Med Toxicol.
11(37)2016.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Cui A, Jin XG, Zhai K, Tong ZH and Shi HZ:
Diagnostic values of soluble mesothelin-related peptides for
malignant pleural mesothelioma: Updated meta-analysis. BMJ Open.
4(e004145)2014.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Arnold DT and Maskell NA: Biomarkers in
mesothelioma. Ann Clin Biochem. 55:49–58. 2018.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Wu T, Zhang W, Yang G, Li H, Chen Q, Song
R and Zhao L: HMGB1 overexpression as a prognostic factor for
survival in cancer: A meta-analysis and systematic review.
Oncotarget. 7:50417–50427. 2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Creaney J, Sneddon S, Dick IM, Dare H,
Boudville N, Musk AW, Skates SJ and Robinson BW: Comparison of the
diagnostic accuracy of the MSLN gene products, mesothelin and
megakaryocyte potentiating factor, as biomarkers for mesothelioma
in pleural effusions and serum. Dis Markers. 35:119–127.
2013.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Creaney J, Dick IM, Meniawy TM, Leong SL,
Leon JS, Demelker Y, Segal A, Musk AW, Lee YC, Skates SJ, et al:
Comparison of fibulin-3 and mesothelin as markers in malignant
mesothelioma. Thorax. 69:895–902. 2014.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Chen Z, Gaudino G, Pass HI, Carbone M and
Yang H: Diagnostic and prognostic biomarkers for malignant
mesothelioma: An update. Transl Lung Cancer Res. 6:259–269.
2017.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Boudville N, Paul R, Robinson BW and
Creaney J: Mesothelin and kidney function-analysis of relationship
and implications for mesothelioma screening. Lung Cancer.
73:320–324. 2011.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Lim CB, Prêle CM, Cheah HM, Cheng YY,
Klebe S, Reid G, Watkins DN, Baltic S, Thompson PJ and Mutsaers SE:
Mutational analysis of hedgehog signaling pathway genes in human
malignant mesothelioma. PLoS One. 8(e66685)2013.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Shi Y, Moura U, Opitz I, Soltermann A,
Rehrauer H, Thies S, Weder W, Stahel RA and Felley-Bosco E: Role of
hedgehog signaling in malignant pleural mesothelioma. Clin Cancer
Res. 18:4646–4656. 2012.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Candido S, Lupo G, Pennisi M, Basile MS,
Anfuso CD, Petralia MC, Gattuso G, Vivarelli S, Spandidos DA, Libra
M and Falzone L: The analysis of miRNA expression profiling
datasets reveals inverse microRNA patterns in glioblastoma and
Alzheimer's disease. Oncol Rep. 42:911–922. 2019.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Falzone L, Romano GL, Salemi R, Bucolo C,
Tomasello B, Lupo G, Anfuso CD, Spandidos DA, Libra M and Candido
S: Prognostic significance of deregulated microRNAs in uveal
melanomas. Mol Med Rep. 19:2599–2610. 2019.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Nikolouzakis TK, Vassilopoulou L,
Fragkiadaki P, Mariolis Sapsakos T, Papadakis GZ, Spandidos DA,
Tsatsakis AM and Tsiaoussis J: Improving diagnosis, prognosis and
prediction by using biomarkers in CRC patients (Review). Oncol Rep.
39:2455–2472. 2018.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Ledda C, Senia P and Rapisarda V:
Biomarkers for early diagnosis and prognosis of malignant pleural
mesothelioma: The quest goes on. Cancers (Basel).
10(203)2018.PubMed/NCBI View Article : Google Scholar
|
|
113
|
De Santi C, Melaiu O, Bonotti A, Cascione
L, Di Leva G, Foddis R, Cristaudo A, Lucchi M, Mora M, Truini A, et
al: Deregulation of miRNAs in malignant pleural mesothelioma is
associated with prognosis and suggests an alteration of cell
metabolism. Sci Rep. 7(3140)2017.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Martínez-Rivera V, Negrete-García MC,
Ávila-Moreno F and Ortiz-Quintero B: Secreted and tissue miRNAs as
diagnosis biomarkers of malignant pleural mesothelioma. Int J Mol
Sci. 19(595)2018.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Falzone L, Candido S, Salemi R, Basile MS,
Scalisi A, McCubrey JA, Torino F, Signorelli SS and Montella M:
Computational identification of microRNAs associated to both
epithelial to mesenchymal transition and NGAL/MMP-9 pathways in
bladder cancer. Oncotarget. 7:72758–72766. 2016.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Falzone L, Scola L, Zanghì A, Biondi A, Di
Cataldo A, Libra M and Candido S: Integrated analysis of colorectal
cancer microRNA datasets: Identification of microRNAs associated
with tumor development. Aging (Albany NY). 10:1000–1014.
2018.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Hafsi S, Candido S, Maestro R, Falzone L,
Soua Z, Bonavida B, Spandidos DA and Libra M: Correlation between
the overexpression of Yin Yang 1 and the expression levels of
miRNAs in Burkitt's lymphoma: A computational study. Oncol Lett.
11:1021–1025. 2016.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Polo A, Crispo A, Cerino P, Falzone L,
Candido S, Giudice A, De Petro G, Ciliberto G, Montella M, Budillon
A and Costantini S: Environment and bladder cancer: Molecular
analysis by interaction networks. Oncotarget. 8:65240–65252.
2017.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Salemi R, Falzone L, Madonna G, Polesel J,
Cinà D, Mallardo D, Ascierto PA, Libra M and Candido S: MMP-9 as a
candidate marker of response to BRAF inhibitors in melanoma
patients with BRAFV600E mutation detected in
circulating-free DNA. Front Pharmacol. 9(856)2018.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Cavallari I, Urso L, Sharova E, Pasello G
and Ciminale V: Liquid biopsy in malignant pleural mesothelioma:
State of the art, pitfalls, and perspectives. Front Oncol.
9(740)2019.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Tuaeva NO, Falzone L, Porozov YB, Nosyrev
AE, Trukhan VM, Kovatsi L, Spandidos DA, Drakoulis N, Kalogeraki A,
Mamoulakis C, et al: Translational application of circulating DNA
in oncology: Review of the last decades achievements. Cells.
8(1251)2019.PubMed/NCBI View Article : Google Scholar
|