Introduction
Transient receptor potential channels (TRP) are
membrane-bound non-selective ion channels present in numerous
different tissues and cell types in human and animal organisms
(1). TRP channels are involved in
intracellular Ca2+ homeostasis. Currently, seven
different families are known: TRPC (C for classical or canonical),
TRPV (vanilloid), TRPM (melastatin), TRPP (polycystin), TRPML
(mucolipin), TRPN (no mechanoreceptor potential C), and TRPA
(ANKTM1) (2). All TRP channels have
six transmembrane domains and an intracellular cytoplasmic N
(amino) and C (carboxy) terminus as structural common features. The
TRPC6 ion channel is a non-selective cation channel and is
important for the calcium balance of cells. TRPC6 is, similar to
TRPC3 and TRPC7, a diacylglycerol (DAG)-sensitive TRPC channel.
This allows a receptor operated opening of the channel (ROC).
The expression of transient receptor potential
channel 6 (TRPC6) was investigated using real-time PCR in rat
tissues and it was revealed that the highest level of expression
was found especially in the lungs, cerebrum and ovaries (1). Further investigations provided
evidence of the occurrence of TRPC6 in the smooth muscle cells of
human blood vessels and their functional significance (3). In an investigation on the calcium
regulation of dystrophin-deficient skeletal muscle of mice, with
the help of antibody-based immunofluorescence staining, TRPC6 was
found in the sarcolemma of muscle fibers and to a lesser extent in
the cytoplasm (4). The authors of
this study concluded that TRPC6 may contribute to the influx of
calcium. A previous study revealed that a mutation in the TRPC6
channel is associated with focal segmental glomerulosclerosis,
which ultimately leads to kidney failure through the death of
podocytes (5). Further associations
with regard to TRPC6 and pathophysiological conditions have been
described for idiopathic pulmonary hypertension, in which the
channels lead to the progression of the disease due to increased
calcium influx (6,7), but also for malignant hyperthermia, in
which TRPC6 was shown to be involved in the pathophysiological
process in a mouse model (8). The
latter study reported that muscle fibers of mice expressing a
distinct mutation (RYR1-p.G2435R) are hypersensitive to activators
of TRPC3/6 and have increased resting intracellular calcium and
sodium (8). This hypersensitivity
can be stopped using agents which inhibit TRPC3/6 indicating that
TRPC channels play a crucial role in causing intracellular
overloads of calcium and sodium in malignant
hyperthermia-susceptible muscle.
The TRPC6 gene is expressed in numerous peripheral
tissues, such as in the lungs, heart, muscles, gastrointestinal
tract and central nervous system (9,10)
while expression in skeletal muscle was comparable to that of the
cerebellum (9). In the present
study for the first time, to the best of our knowledge, TRPC6 was
described in human skeletal muscle. However, data in previous
studies (9,10) were related to TRPC6 gene expression,
but not to the detection of the protein. Humans are mammals,
however most studies have used tissues or cells from non-human
mammals to examine human skeletal muscle tissue. Therefore, in the
present study, immunohistochemistry (IHC) was used to examine human
skeletal muscle tissue samples of cadavers from the local body
donation program and from departments of the Institute of Pathology
of Saarland University in order to establish the occurrence of
TRPC6 in human skeletal muscle tissue.
Patients and methods
Donors
For this investigation six human cadaver donors from
the local body donation program of Saarland University and two
cadavers from the departments of the Institute of Pathology of
Saarland University, were used. The bodies of two cadavers were not
preserved, while four donors were preserved according to a protocol
in a previous study by Janczyk et al (11) and two other donors using classical
formaldehyde fixation. Data of the body donors is provided in
Table I.
 | Table IList of skeletal muscle samples which
were investigated using immunohistochemistry. |
Table I
List of skeletal muscle samples which
were investigated using immunohistochemistry.
| Donor no. | Year of birth | Year of death | Preservation | Sex | Cause of death | Department | IHCa |
|---|
| 1 | 1928 | 2018 | Formalin | Female | Cardiac failure | Anatomy | 1,2,3 |
| 2 | 1949 | 2018 | Formalin | Male | Pneumonia | Anatomy | 1,3 |
| 3 | 1944 | 2021 | Protocol by Janczyk
et alb | Female | Pulmonary
embolism | Anatomy | 1,4 |
| 4 | 1945 | 2021 | Protocol by Janczyk
et alb | Male | Pneumonia | Anatomy | 1,4 |
| 5 | 1938 | 2021 | Protocol by Janczyk
et alb | Male | Prostate
carcinoma | Anatomy | 1,2,3,4 |
| 6 | 1937 | 2021 | Protocol by Janczyk
et alb | Female | Cardiac failure | Anatomy | 1,2,3,4 |
| 7 | 1947 | 2020 | None | Female | Cardiac failure | Pathology | 1,2 |
| 8 | 2000 | 2020 | None | Male | Leukemia | Pathology | 1,2 |
The present study was approved by the Permanent
Ethics Committee of the Saarland Medical Association (Ärztekammer
des Saarlandes), Saarbrücken, Germany (approval no. 163/20 for
samples obtained from the Department of Anatomy and approval no.
130/21 for the samples obtained from the Department of Pathology of
Sarlaand University, Homburg, Germany). This committee is
responsible for all medical research projects in Saarland.
Samples
Samples from the Departments of Anatomy and
Pathology of Saarland University were obtained from May to October
2021. A biopsy punch and a scalpel were used to remove the tissue
from the Musculus (M.) deltoideus, pectoralis major, trizeps
brachii and rectus femoris from the right half of the bodies of the
donors. For comparability M. pectoralis major samples from each
donor were used. Furthermore four M. rectus femoris, four M.
trizeps brachii and five M. deltoideus samples were included.
Immunostaining
After fixation in 4% buffered formaldehyde at room
temperature overnight the specimens were embedded in paraffin and
7-µm thin sections were prepared using a microtome. The specimens
were first stained with hematoxylin and eosin (H&E) at room
temperature for 5 min and assessed to determine the intact
morphology. The first step of IHC was antigen retrieval using 1%
citrate-buffered solution for 60 min at 95˚C in heating incubator.
After washing the samples twice for 2 min in PBS each time, they
were incubated with a knockout-validated antibody against TRPC6
channel antigen structures (cat. no. ACC-017; Alomone Labs). The
specificity and quality of the antibody was assessed using
peptide-blocked control samples (cat. no. BLP-CC017; Alomone Labs).
For antibody specificity, 40 µg control peptide was dissolved with
20 µl PBS and then incubated with 40 µl 1:100 diluted TRPC6 primary
antibody overnight at 7˚C, in tubes. Negative controls were
incubated with rabbit serum (from an untreated rabbit obtained from
the Institute for Biochemistry, Homburg, Germany) instead of TRPC6
antibodies. The protein concentration in both solutions was similar
(~0.01 mg/ml). Horseradish peroxidase-conjugated goat anti-rabbit
antibodies (cat. no. A10547; 1:500; Invitrogen AG; Thermo Fisher
Scientific, Inc.) were used as secondary antibodies. The sections
were incubated with the secondary antibodies for 10-12 h at room
temperature in a humidity chamber. The dilutions as aforementioned
(1:100 and 1:500) were performed with normal goat serum (cat. no.
01-6201; Invitrogen; Thermo Fisher Scientific, Inc.). The added
chromogen, diaminobenzidine (cat. no. SK-4103; Vector Laboratories,
Inc.), became visible as a brown color after the HRP substrate
catalyzed the reaction. The incubation time with the DAB substrate
was 10 min. A total of 21 samples were examined using this
procedure. Evaluation was performed using a light microscope (Nikon
Eclipse E200; Nikon Corporation). Microphotographs were obtained
using a digital camera (TrueChrome Metrics with software TCapture;
Tucsen Photonics Co., Ltd.).
Results
The results were evaluated microscopically and the
samples were categorized according to staining intensity: Strong,
weak and negative. From all the investigated samples, 19 samples
exhibited strong staining and 2 samples exhibited weak staining.
Nearly all (11 out of 12) samples that were preserved according to
a protocol in a previous study by Janczyk et al (11) showed a strong immunohistochemical
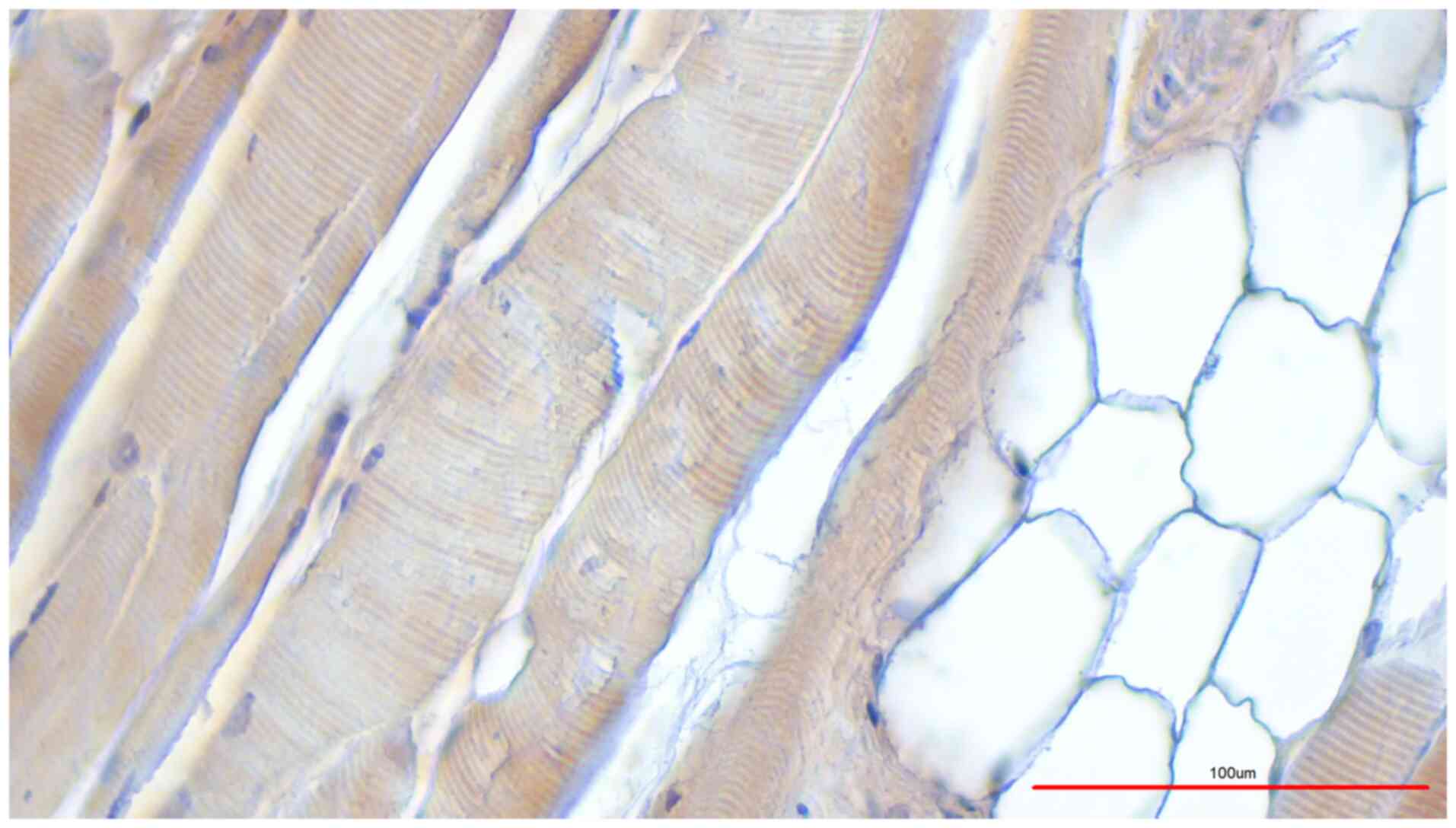
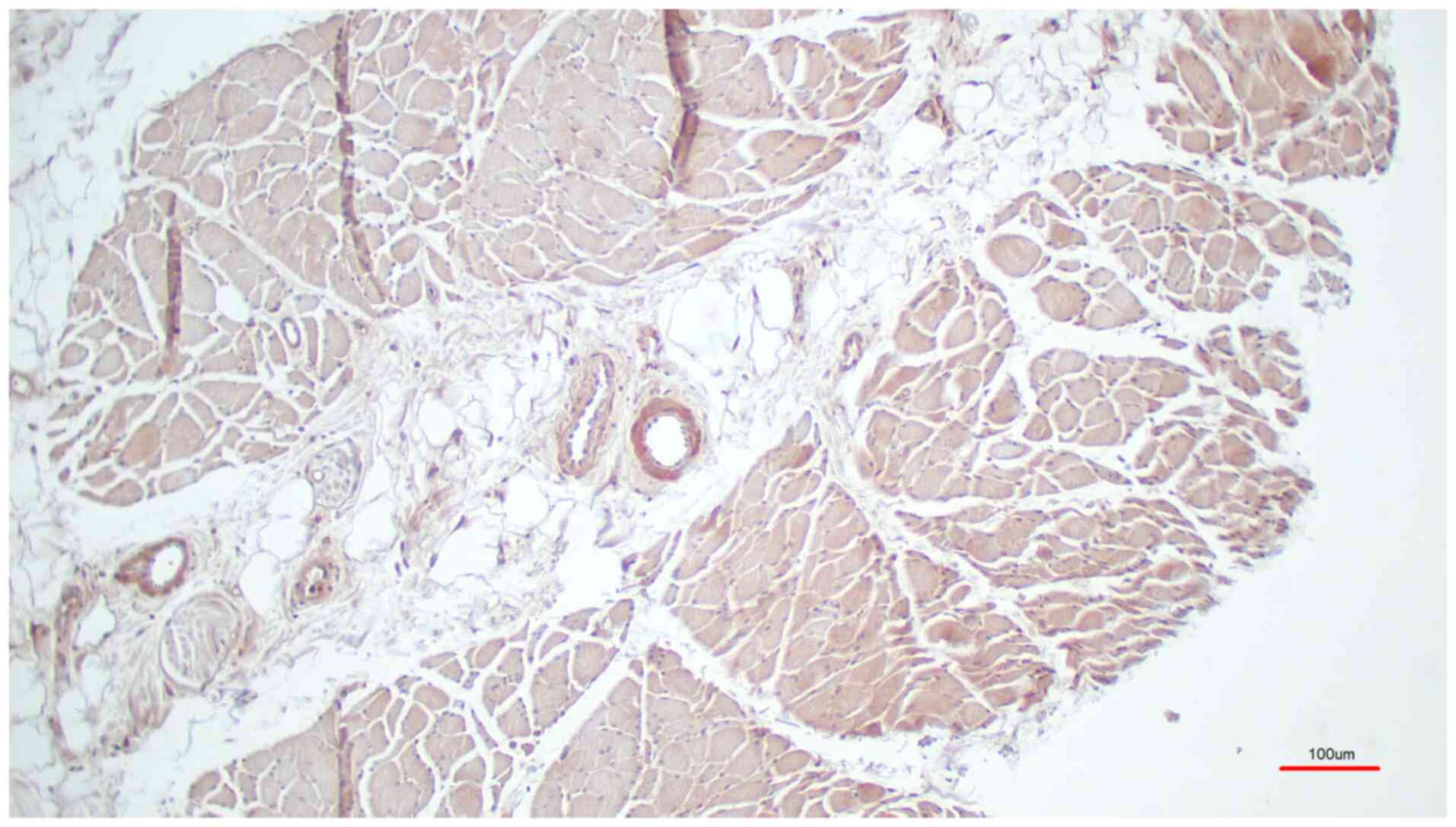
signal (Fig. 1, M. pectoralis
major), while one sample was weakly stained. The four unfixed
skeletal muscle samples consistently showed a strong
immunohistochemical signal similar to the protocol by Janczyk et
al (11). The five samples
preserved using classical formaldehyde fixation showed a strong
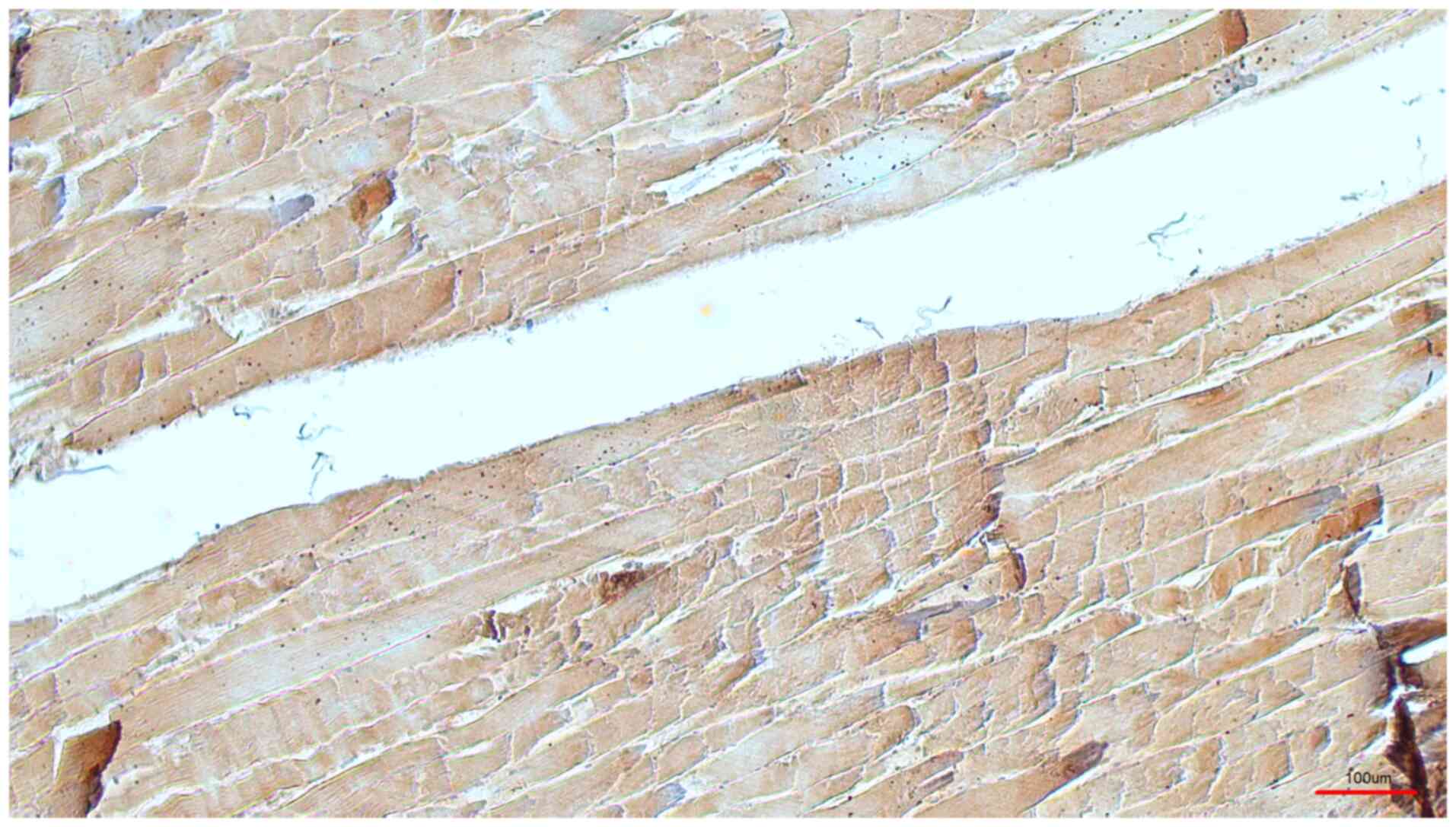
immunohistochemical signal in four cases (Fig. 2, M. triceps brachii), and a weak
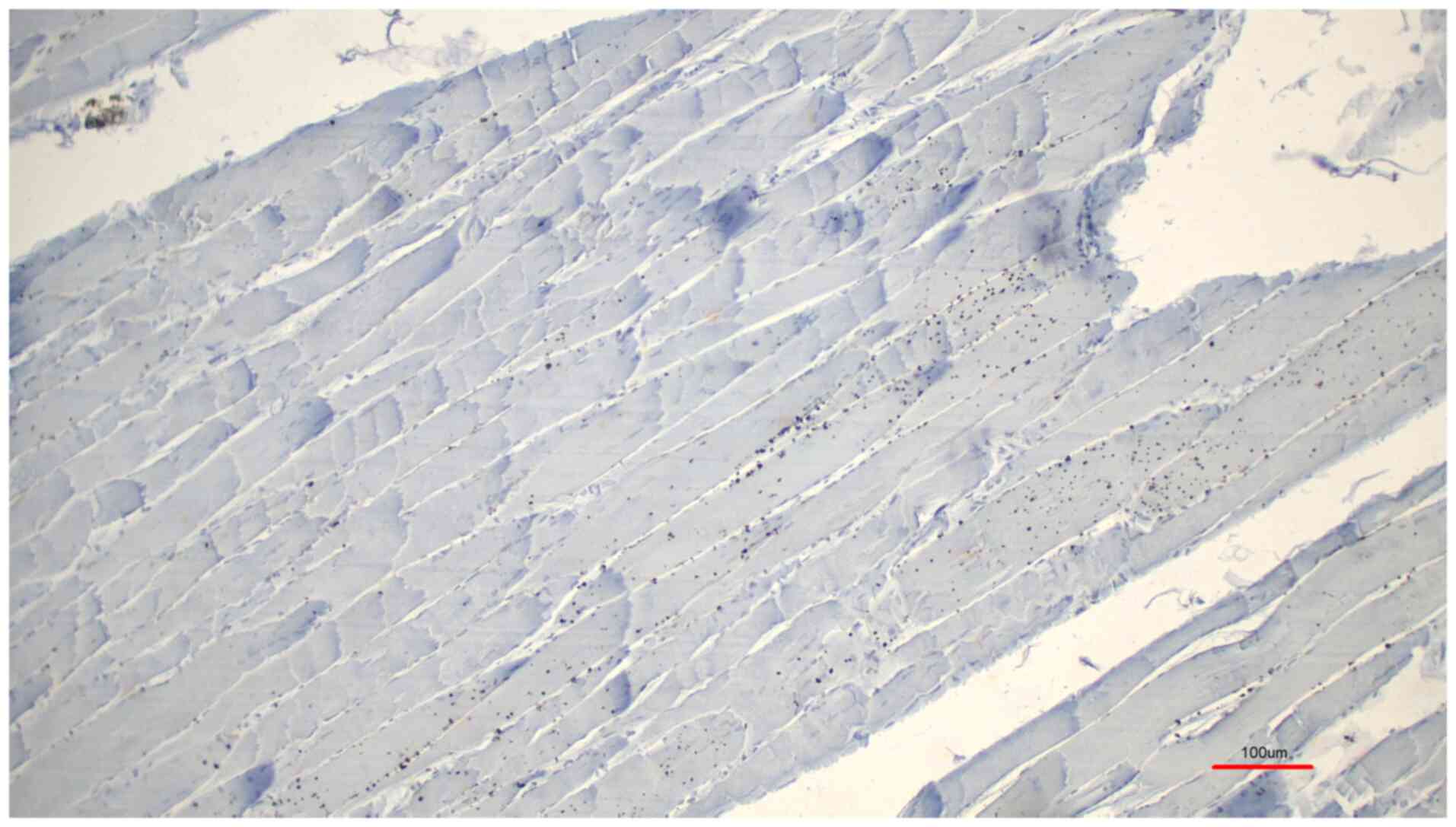
signal was detected in one case. Both controls, the normal rabbit
serum control (Fig. 3) and the
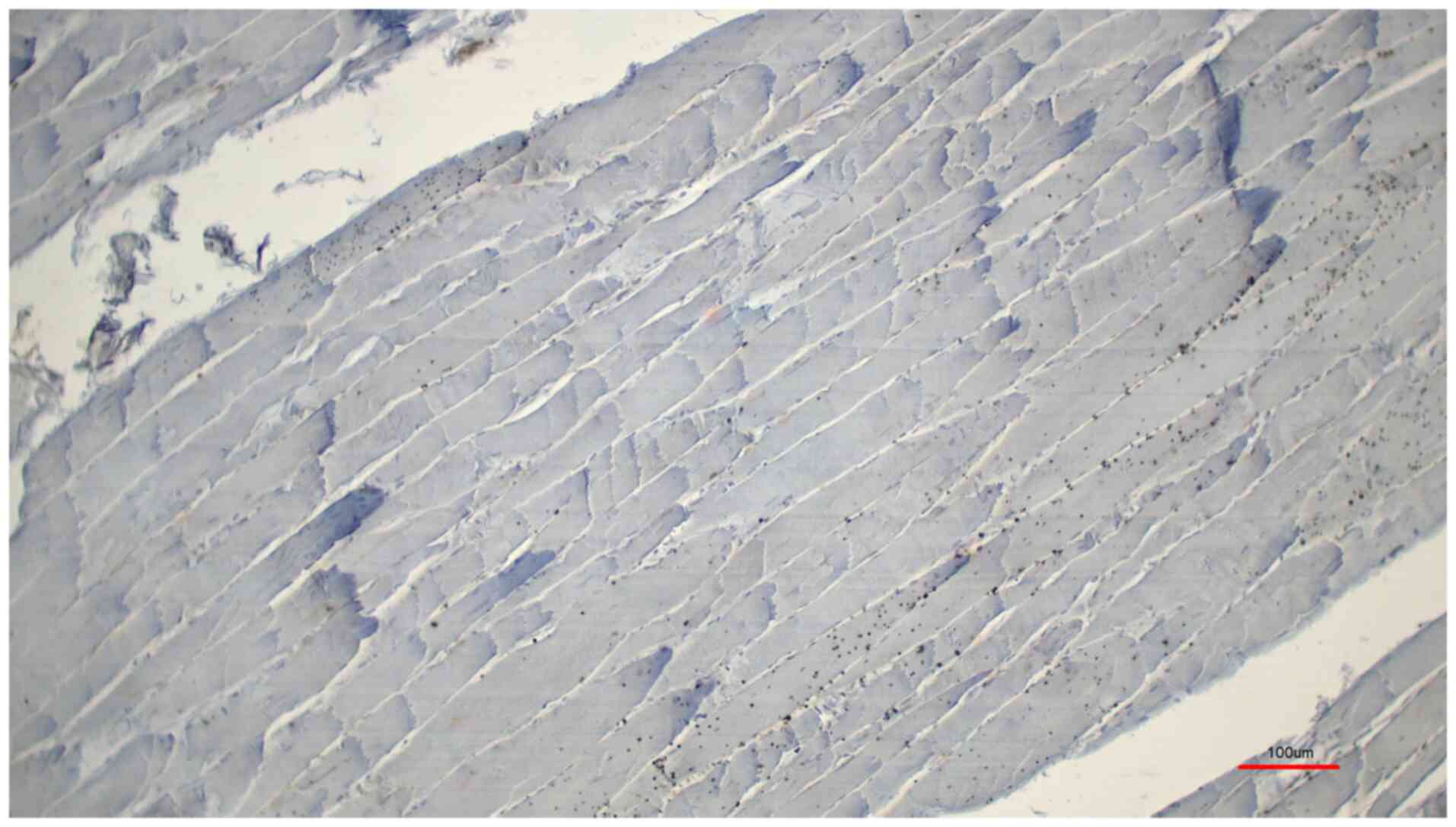
control with the peptide pre-incubation (Fig. 4), exhibited almost no
immunohistochemically-stained structures. Another representative
example of the presence of TRPC6 was revealed in a sample of M.
deltoideus, which was obtained from an autopsy of a non-preserved
cadaver (Table I, donor 8; Fig. 5, M. deltoideus). Collectively,
representative examples of TRPC6 detection in human bodies are
presented in M. deltoideus (Fig.
5), M. pectoralis major (Fig.
1) and M. triceps brachii (Fig.
2). An example of M. rectus femoris was lacking since it
appeared identical to the other skeletal muscles. Skeletal muscles
were presented from three different post mortem conditions:
Non-preserved (Fig. 5), preserved
according to the protocol by Janczyk et al (11) (Fig.
1) and preserved using classical formaldehyde fixation
(Fig. 2).
All skeletal muscle fibers were labelled with
brownish color whereas the fat cells were not labelled (Fig. 5). In summary, TRPC6 was found in the
fibers of all investigated skeletal muscles independent of whether
the tissue was freshly obtained during autopsy or after
preservation.
Discussion
The immunohistological data for TRPC6 using the
described Alomone antibody appears to be valid since corresponding
bands in the western blots were absent when tissues from knockout
mice were assessed (12). A
limitation of the present study is that it is lacking western blot
analysis because the unfixed tissue was prepared for histology.
However, western blotting was performed using the identical TRPC6
antibody as aforementioned with homogenized human vessels
containing mainly smooth muscle tissue. This was a recently
published study (13).
To investigate human tissues, the body donation
programs of anatomical institutes are of great value if the body
donors have agreed to their donations not only for teaching but
also for experimental research in an informed consent during their
lifetime.
It is necessary to preserve the bodies for different
purposes such as anatomical or surgical teaching and research. In
the present study, it was demonstrated that both a classical
preservation using formalin and a ‘soft preservation’ using pickle
salt and alcohol could conserve a protein of interest and did not
inhibit its detection using immunohistochemistry.
In summary, the present study detected TRPC6 protein
in human skeletal muscle tissue for the first time, to the best of
our knowledge. The results are consistent with a previous study, in
which, for example, skeletal muscle of the mouse displayed channels
in the sarcolemma (4). This study
assessed the mRNA expression, protein levels and localization of
TRPC3, TRPC6, TRPV4 and TRPM7. All these aforementioned channels
may mediate store-operated, stretch-activated and background
Ca2+-influx into muscle fibers. In addition, this
previous study revealed that further investigations are required to
ascertain whether TRP channels could be targets for new drugs of
Duchenne muscular dystrophy (4).
However the exact localization should be
investigated further using high-dissolving techniques such as
immune electron microscopy. The junctional sarcoplasmic reticulum
including L-tubule and T-tubule and the triads play a major role in
the electro-mechanical linkage and are areas where calcium channels
are localized (14). For the
non-junctional sarcoplasmic reticulum within striated muscles, it
has been demonstrated in 293 cells that TRPC channels interact with
the proteins junctate and inositol-1,4,5-triphosphate receptor
(15). Junctate has been
demonstrated to be associated with the calcium ion content of the
stores of the sarcoplasmic reticulum (16). Junctate has also been identified as
a calcium-sensing component of ORAI1 and stromal interaction
protein 1 (STIM1) (17,18). Notably, TRPC3 and presumably TRPC1
interact with triadic proteins such as ryanodine receptor type 1
(RyR1) and are involved in the release of calcium ions during the
excitation-contraction process (19). In this previous study RyR1- or
TRPC3-interacting triadic proteins were found by using a
co-immunoprecipitation assay after establishing potential candidate
proteins with chemical cross-linking of triadic proteins and mass
spectroscopy. The results indicated that in muscle, TRPC3 and RyR1
interact indirectly via a linker protein, TRPC1. A panel of data is
available on the physiological and presumed pathological role of
TRPC channels in smooth muscle cells (20-22).
The presented study is limited to descriptive results. In future,
functional investigations with human muscle cells or tissue is
necessary.
In view of the studies of animal skeletal muscles in
which the importance of TRPC6 and TRPC3 for calcium flux, and thus
also for the muscular contraction, was demonstrated (4,19) it
is evident that TRPC channels could play an important role. Thus,
TRPC6 and TRPC3 and most likely other TRPC channels are important
for the calcium homeostasis in human muscle fibers and may be
targets for treatment approaches in various muscular diseases.
Acknowledgements
We would like to thank Ms Irina Scheck, Mr Ronald
Dollwett, Ms Helga Meyer and Ms Barbara Michahelles-Horzella (all
from the Institute of Anatomy and Cell Biology, Saarland
University, Homberg, Germany) for the technical support.
Funding
Funding: No funding was received.
Availability of data and materials
All microphotographs from the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
All authors planned and conducted the study. DS, JA
and AG performed the experiments. DS and JA evaluated the data. DS
and TT confirm the authenticity of all the raw data. DS and TT
planned the study and wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Permanent
Ethics Committee of the Saarland Medical Association (Ärztekammer
des Saarlandes), Saarbrücken, Germany (approval no. 163/20 for
samples obtained from the Department of Anatomy and approval no.
130/21 for the samples obtained from the Department of Pathology of
Sarlaand University, Homburg, Germany).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Garcia RL and Schilling WP: Differential
expression of mammalian TRP homologues across tissues and cell
lines. Biochem Biophys Res Commun. 239:279–283. 1997.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Venkatachalam K and Montell C: TRP
channels. Annu Rev Biochem. 76:387–417. 2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gonzalez-Cobos JC and Trebak M: TRPC
channels in smooth muscle cells. Front Biosci (Landmark Ed).
15:1023–1039. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
4
|
Krüger J, Kunert-Keil C, Bisping F and
Brinkmeier H: Transient receptor potential cation channels in
normal and dystrophic mdx muscle. Neuromuscul Disord. 18:501–513.
2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Winn MP, Conlon PJ, Lynn KL, Farrington
MK, Creazzo T, Hawkins AF, Daskalakis N, Kwan SY, Ebersviller S,
Burchette JL, et al: Medicine: A mutation in the TRPC6 cation
channel causes familial focal segmental glomerulosclerosis.
Science. 308:1801–1804. 2005.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yamamura H, Suzuki Y and Yamamura A:
Pathophysiological roles of TRPC6 channels in pulmonary arterial
hypertension. Nihon Yakurigaku Zasshi. 155:230–235. 2020.PubMed/NCBI View
Article : Google Scholar : (In Japanese).
|
|
7
|
Yu Y, Fantozzi I, Remillard CV, Landsberg
JW, Kunichika N, Platoshyn O, Tigno DD, Thistlethwaite PA, Rubin LJ
and Yuan JX: Enhanced expression of transient receptor potential
channels in idiopathic pulmonary arterial hypertension. Proc Natl
Acad Sci USA. 101:13861–13866. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rafael Lopez J, Kaura V, Hopkins P, Liu X,
Uryach A, Adams J and Allen PD: Transient receptor potential cation
channels and calcium dyshomeostasis in a mouse model relevant to
malignant hyperthermia. Anesthesiology. 133:364–376.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Riccio A, Medhurst AD, Mattei C, Kelsell
RE, Calver AR, Randall AD, Benham CD and Pangalos MN: mRNA
distribution analysis of human TRPC family in CNS and peripheral
tissues. Brain Res Mol Brain Res. 109:95–104. 2002.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kunert-Keil C, Bisping F, Krüger J and
Brinkmeier H: Tissue-specific expression of TRP channel genes in
the mouse and its variation in three different mouse strains. BMC
Genomics. 7(159)2006.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Janczyk P, Weigner J, Luebke-Becker A,
Kaessmeyer S and Plendl J: Nitrite pickling salt as an alternative
to formaldehyde for embalming in veterinary anatomy - A study based
on histo- and microbiological analyses. Ann Anat. 193:71–75.
2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Dietrich A, Mederos Y, Gollasch M,
Gollasch M, Gross V, Storch U, Dubrovska G, Obst M, Yildirim E,
Salanova B, et al: Increased vascular smooth muscle contractility
in TRPC6-/- mice. Mol Cell Biol. 25:6980–6989.
2005.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Abdinghoff J, Servello D, Jacobs T,
Beckmann A and Tschernig T: Evaluation of the presence of TRPC6
channels in human vessels: A pilot study using
immunohistochemistry. Biomed Rep. 16(42)2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Perni S: The Builders of the Junction:
Roles of Junctophilin1 and Junctophilin2 in the assembly of the
sarcoplasmic reticulum-plasma membrane junctions in striated
muscle. Biomolecules. 12(109)2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Treves S, Franzini-Armstrong C, Moccagatta
L, Arnoult C, Grasso C, Schrum A, Ducreux S, Zhu MX, Mikoshiba K,
Girard T, et al: Junctate is a key element in calcium entry induced
by activation of InsP3 receptors and/or calcium store depletion. J
Cell Biol. 166:537–548. 2004.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Divet A, Paesante S, Grasso C, Cavagna D,
Tiveron C, Paolini C, Protasi F, Huchet-Cadiou C, Treves S and
Zorzato F: Increased Ca2+ storage capacity of the
skeletal muscle sarcoplasmic reticulum of transgenic mice
over-expressing membrane bound calcium binding protein junctate. J
Cell Physiol. 213:464–474. 2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Srikanth S, Jew M, Kim KD, Yee MK,
Abramson J and Gwack Y: Junctate is a Ca2+-sensing
structural component of Orai1 and stromal interaction molecule 1
(STIM1). Proc Natl Acad Sci USA. 109:8682–8687. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kiviluoto S, Decuypere JP, De Smedt H,
Missiaen L, Parys JB and Bultynck G: STIM1 as a key regulator for
Ca2+ homeostasis in skeletal-muscle development and
function. Skelet Muscle. 1(16)2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Woo JS, Kim DH, Allen PD and Lee EH:
TRPC3-interacting triadic proteins in skeletal muscle. Biochem J.
411:399–405. 2008.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Pulina MV, Zulian A, Baryshnikov SG, Linde
CI, Karashima E, Hamlyn JM, Ferrari P, Blaustein MP and Golovina
VA: Cross talk between plasma membrane Na(+)/Ca (2+) exchanger-1
and TRPC/Orai-containing channels: Key players in arterial
hypertension. Adv Exp Med Biol. 961:365–374. 2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Poburko D, Fameli N, Kuo KH and van
Breemen C: Ca2+ signaling in smooth muscle: TRPC6, NCX
and LNats in nanodomains. Channels (Austin). 2:10–12.
2008.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang J, Laurier LG, Sims SM and
Preiksaitis HG: Enhanced capacitative calcium entry and TRPC
channel gene expression in human LES smooth muscle. Am J Physiol
Gastrointest Liver Physiol. 284:G1074–G1083. 2003.PubMed/NCBI View Article : Google Scholar
|