Introduction
Apolipoprotein E (ApoE) is a constituent of
chylomicrons, and plasma very low density lipoproteins (VLDL), and
high density lipoproteins (HDL). Human ApoE-encoded by the
APOE gene-is a polymorphic protein with three common
isoforms (ApoE2, ApoE3, and ApoE4) defined by two single nucleotide
polymorphisms (rs429358 and rs7412) in the coding region of exon 4
that overlaps with a well-defined CpG island (CGI) (1). These isoforms have differing
affinities for binding to LDL receptors: ApoE3 and ApoE4 bind with
similar affinity, while ApoE2 has only 2% of this binding affinity.
ApoE3 is the most commonly expressed isoform in healthy people
(2). ApoE protein plays a key role
in lipid metabolism, including the redistribution of lipoproteins
and maintenance of cholesterol homeostasis by stabilization of
lipoprotein particles in the circulation and enhancing uptake by
the liver and other tissues. Different genetic studies have
revealed the association between APOE and its ε2/ε3/ε4
alleles with several pathological conditions and disorders,
including Alzheimer's disease (AD) (2), cardiovascular disease (3), familial dysbetalipoproteinemia
(4), metabolic syndrome and
diabetes (5). The ε4 allele has
been associated with higher total and LDL cholesterol levels than
in patients with the ε3 allele, and thus reported as the strongest
genetic factor for AD and coronary artery disease in several
studies and across several ethnic groups (1).
Furthermore, it has been reported that ε4 carriers
are less responsive to lipid lowering therapy (i.e Statins) but
more sensitive to a low fat diet and physical activity (6). A population-based study reported that
ε4/ε4 carriers had a 2.28-fold increased risk of discontinuing a
statin therapy compared to ε3/ε3 carriers (7), which has been attributed to
therapeutic inefficacy and severe side effect (8). Moreover, ε2 carriers had a better
response in TG, TC, and LDL-C reduction percentage in response to
statins treatment, compared with ε4 carriers (9). Thus, knowing the APOE genotype
may be useful in guiding the choice of treatment, in assessing the
potential side effects, and in suggesting the complementary
approaches that can be used to control metabolic variations in ε4
carriers. In addition, three common single nucleotide polymorphisms
(SNPs) rs449647 (A491-T), rs769446 (C427-T), and rs405509 (G219-T)
located in the promoter region of the APOE gene have been
reported to influence APOE transcriptional activity, most
likely through differential binding of transcription factors and
thus differential APOE gene expression (10).
Conversely, it is well known that epigenetic
mechanisms such as DNA methylation regulate the transcription of
several genes and has been linked to the development of various
diseases. Several studies investigated the regulatory mechanisms
underlying APOE gene regulation (11). Promoters and regulatory element
activities (i.e enhancers) are affected by cytosine methylation at
CpG sites in the genome (12). Gene
expression inhibition is correlated with hypermethylated promoters
and non-promoter sites that are located within enhancer regions
leading to loss of enhancer activity and consequently
transcriptional inactivation of specific genes (13). Thus, the pleiotropic roles of
APOE may lie in its unique epigenetic properties (1). To the best of our knowledge, no
previous studies have investigated the association of APOE
gene variants and methylation with the risk of dyslipidemia in type
2 diabetes (T2D) patients. Therefore, the present study aimed to
determine the allelic and genotypic frequencies of the APOE
gene polymorphisms and the three promoter variants to investigate
whether APOE genotypes influenced lipid profiles in
Palestinian T2D patients with and without dyslipidemia. Moreover,
whether DNA methylation of the APOE promoter region differed
in diabetic dyslipidemia patients compared with those without
dyslipidemia was also assessed. This genetic study may provide
additional information regarding the Palestinian population beyond
traditional risk factors.
Materials and methods
Study participants
A total of 204 unrelated hospitalized T2DM patients
were randomly recruited from Ramallah hospital between January and
April 2019. The median age of the subjects studied was 62 years
(age range 40-97 years), 125 were males and 79 were females. Among
these subjects, 96 were diagnosed with diabetic dyslipidemia and
108 T2DM subjects without dyslipidemia, and none of these patients
had been previously treated with lipid lowering drugs. Diagnosis of
T2D was based on World Health Organization criteria: Fasting blood
glucose ≥126 mg/dl and/or currently on use of antidiabetic
medication (14). Patients who were
diagnosed with type 1 diabetes were excluded from the study.
Dyslipidemia was defined as a level of total cholesterol (TC)
>200 mg/dl, triglycerides (TG) >50 mg/dl, LDL-C >130
mg/dl, HDL-C <40 mg/dl, or under medication of lipid lowering
drugs (15). The study was
conducted according to the guidelines expressed in the Declaration
of Helsinki (16), and written
informed consent was obtained from all enrolled participants. The
study procedure was approved by the local ethical committee at
Al-Quds University (East Jerusalem, Palestine; approval no.
71/REC).
Demographic, anthropometric, and
biochemical measurements
Demographic information including sex and age was
collected from the patient's medical record using a well-structured
questionnaire. Body mass index (BMI) was calculated as weight
divided by height squared (kg/m2). Blood pressure was
measured with a mercury sphygmomanometer at the time of blood
sampling. Blood samples (5 ml) were drawn after a 12-h minimum
fast. Plasma glucose, Glycated Hb (HbA1C), cholesterol, HDL
cholesterol, and triglyceride were determined using standard
methods of commercial kits. LDL cholesterol was calculated based on
the Friedewald formula (17).
DNA extraction and genotyping
Genomic DNA was extracted from whole blood (300 µl)
using a genomic QIAamp DNA purification kit according to the
manufacturer's instructions (Qiagen GmbH). The DNA concentration
was determined using a NanoDrop 1000 spectrophotometer (Thermo
Fisher Scientific, Inc.) and was stored at -20˚C until required for
further analysis. Amplicon-based next generation sequencing (NGS)
was used for genotyping as previously described (18). Briefly, three primers (two forward
and one reverse) were used in multiplex PCR to target the two SNPs
of APOE rs429358 [C/T] and rs7412 [C/T]. Based on these 2
SNPs, the APOE alleles (ε2, ε3, and ε4) were determined.
Moreover, four primers (two forward and two reverse)
were used to target the three SNPs in APOE promoter region
rs769446 [T/C], rs449647[A/T], and rs405509 [G/T]. All primers were
modified with over hanged Illumina adaptor sequences at the 5' ends
that were added to the forward
(5'-CGTCGGCAGCGTCAGATGTGTATAAGAGACA-3') and reverse primers
(5'-GTCTCGTGGGCTCGGAGATGTGTATAAGAGA-3'). The primer sequences and
the final length of target sequences are shown in Table I. The reaction was carried out using
3 µl of the extracted DNA in a final volume of 25 µl, which
contained 12.5 µl PCRBIO HS Taq Mix Red (PCR Biosystems, Ltd.), 8.5
µl double distilled water (dH2O) and 0.2 µM of each
primer. The amplification conditions were as follows: Initial
denaturation at 95˚C for 5 min followed by 32 cycles of 95˚C for 30
sec, 65˚C for 30 sec, and 72˚C for 40 sec, with a final extension
step of 72˚C for 6 min. The PCR products of all samples were
visualized on 1.5% agarose gel, cleaned using the Agencourt AMPure
XP system (X1, cat. no. A63881; Beckman Coulter Genomics), and
eluted in 25 µl elution buffer. DNA library preparation and
barcoding were performed using a Nextera XT Index Kit (Illumina,
Inc.) as described previously (18). Samples were deep sequenced on a
NextSeq 500/550 machine using the 150-cycle Mid Output Kit
(Illumina, Inc.). The obtained DNA sequences were analyzed using
the Galaxy program (https://usegalaxy.org/). A total of 10 virtual probe
sequences (four for the two APOE gene variants and six for
the three APOE promoter variants) were used to identify the
targeted polymorphisms (Table I).
The genotypes were determined, based on the calculated ratio
between the read counts for wild type and mutant alleles, for all
SNPs (APOE ε and promoter polymorphisms) in each sample. All
genotyping personnel were blinded to the clinical data.
 | Table ISequences of the primers and virtual
probes used in PCR and sequence analysis. |
Table I
Sequences of the primers and virtual
probes used in PCR and sequence analysis.
| SNP | Primer name | Primer sequence,
5'-3'a | PCR target/bp |
|---|
| rs429358T/C and
rs7412C/T | APOEF1 |
TCCAAGGAGCTGCAGGCGGCGCA | APOE
gene/120 and 218 bp |
| | APOEF2 |
AGAGCACCGAGGAGCTG | |
| | APOER |
GCCCCGGCCTGGTACACTGCCA | |
| rs449647 T/A,
rs769446 C/T and rs405509 A/C | ProAPOEF1 |
CACGCCTGGCTAACTTTTGT | APOE
Promoter/248 and 225 bp |
| | ProAPOEF2 |
AAGGACAGGGTCAGGAAAGG | |
| | ProAPOER1 |
TCCTGGATCCCAGAAAGAAA | |
| | ProAPOER2 |
AGGTGGGGCATAGAGGTCTT | |
| Methylation
primers | APOEPIF |
GAGGGGTTATTTTTAGGAGTAT | APOE
promoter/324 bp |
| | APOEPIR |
TCCCAATCCTAAAATTCAAATT | |
| Gene | Probe Name | Probes
sequences | Targeted SNP |
| APOE ε
probes | rs429358T |
GACGTGTGCGGC | T |
| | rs429358C |
GACGTGCGCGGC | C |
| | rs7412C |
GCAGAAGCGCCTGG | C |
| | rs7412T |
GCAGAAGTGCCTGG | T |
| APOE
promoter probes | rs449647A |
TCTCAAACTCCTG | A |
| | rs449647T |
TCTCAATCTCCTG | T |
| | rs769446T |
GTGAGCTACCGC | T |
| | rs769446C |
GTGAGCCACCGC | C |
| | rs405509A |
GTCTGTATTACTG | T |
| | rs405509C |
GTCTGGATTACTG | G |
DNA methylation analysis
Based on the availability and quantity of DNA
samples, a sub group of 119 samples (58 with dyslipidemia and 61
without dyslipidemia) were randomly selected to examine whether
APOE promoter methylation was associated with a risk of
dyslipidemia. The median age of the subjects studied was 62 years
(age range 40-88 years), 70 were males and 49 were females. A
minimum concentration of genomic DNA >50 ng was used in the
methylation analysis and bisulfite converted using (CiTi Converter
DNA Methylation kit, Gdansk) according to the manufacturer's
protocol, by which the unmethylated cytosine residues were
converted into thymine residues, while the methylated cytosines
remained unchanged. A total of six CpG dinucleotides in the
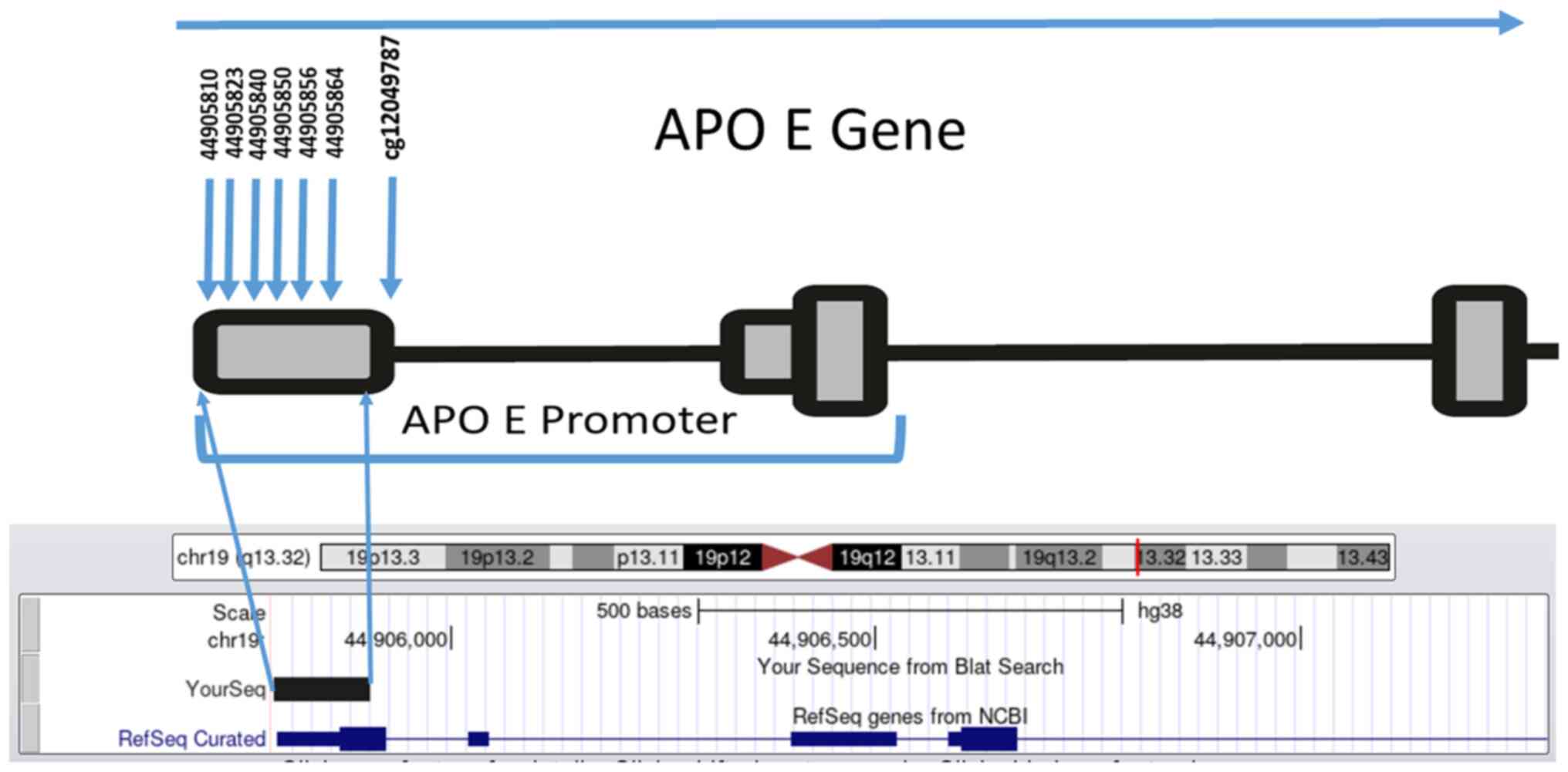
APOE promoter, (CpG1-CpG6; Fig.
1) located on chromosome 19 (44905755-44906078) were studied.
This region was selected based on a previous study which showed
that CpG (cg12049787) within the APOE promoter was
associated with AD and correlated with APOE expression
(cg26190885, cg08955609) (19).
Thus, a fragment of 150 bp that contained six CpGs upstream of the
CpG (cg12049787), was amplified using two newly designed primers
(Fig. 1).
The sequences of the primers, modified with over
hanged Illumina adaptors at the 5' ends, that were used for
amplification of our target fragment are shown in Table I. PCR was performed in a final
volume of 25 µl consisting of 12.5 µl (X2) hot start master mix
(PCRBIO HS Taq MixRed, PCR Biosystems, Ltd.), 10 µM of each primer
and 5 µl bisulfite converted DNA. DNA library preparation and
barcoding were performed as described above. The methylation ratios
were determined using a Python script (methratio.py) and
MethylDackel free program on the UseGalaxy.eu
website.
Statistical analysis
All data were analyzed using SPSS version 20.0 (IBM
Corp). Demographic characteristics are reported as the mean ± SD or
n (%). Comparisons between the two groups were performed using an
unpaired Student's t-test or a χ2 test, as appropriate
and confirmed by a nonparametric analysis using Mann-Whitney test.
Allele and genotype differences between groups and deviations from
Hardy-Weinberg (HW) equilibrium were assessed using a χ2
test. Stepwise logistic regression analysis was applied to assess
whether DNA methylation levels at the six selected CpG
dinucleotides were associated with dyslipidemia.
Results
Study participants
The study included 204 T2DM individuals; the
anthropometric, clinical, and biochemical measurements for T2D
patients with and without dyslipidemia are presented in Table II. Mean BMI, TC, TG, and LDL-C were
significantly higher in participants with dyslipidemia than in
those without dyslipidemia (P<0.05) and confirmed by a
nonparametric analysis using Mann-Whitney test (Table SI). No significant differences were
found in age, sex, systolic and diastolic blood pressure, fasting
blood glucose level, and HbA1C (P>0.05).
| Table IIDemographics and biochemical
characteristics of study subjects. |
Table II
Demographics and biochemical
characteristics of study subjects.
| Variable | With dyslipidemia,
n=96e | Without
dyslipidemia, n=108e | P-value |
|---|
| Age | 62.3 (9.86) | 63.1 (10.53) | 0.62c |
| Sex female/male,
n | 34/62 | 45/63 | 0.36d |
| Body mass index,
kg/m2 | 29.9 | 25 |
2.43x10-18b,c |
| Systolic blood
pressure, mmHg | 135.02 (18.75) | 139.85 (20.49) | 0.08a,d |
| Diastolic blood
pressure, mmHg | 77.97 (13.11) | 81.06 (11.15) | 0.07a,c |
| HbA1C, % | 8.02 (1.3) | 8.13 (1.5) | 0.59d |
| Fasting blood
glucose, g/dl | 241.62
(108.66) | 227.22 (92.82) | 0.31d |
| Total triglyceride,
mg/dl | 252.6 (148.79) | 149.68 (60.32) |
4.18x10-9b,d |
| Total cholesterol,
mg/dl | 237.38 (86.47) | 161.78 (50.7) |
5.34x10-12b,d |
| High-density
lipoprotein cholesterol, mg/dl | 36.07 (13.17) | 49.58 (17.99) |
3.94x10-9b,d |
| Low-density
lipoprotein cholesterol, mg/dl | 161.51 (54.32) | 96.18 (41.64) |
1.06x10-17b,d |
APOE genotypes and allele
frequencies
The genotype and allelic distributions of the
APOE gene are described in Table III. The distribution of
APOE genotypes was in HW equilibrium in both groups
(P>0.05). No significant differences were observed in the
APOE genotypes and alleles frequencies between the two
groups (P>0.05). In all subjects, the frequency of each genotype
in descending order was (ε3/ε3, 79.4%; ε2/ε3, 9.3%; ε3/ε4, 7.4%;
ε2/ε2, 2.5%; ε2/ε4, 1.5%). No ε4/ε4 genotype was detected. The ε3
allele was the most common (81.3%) followed by the ε2 allele
(11.2%) and then the ε4 allele (7.4%).
| Table IIIAPOE genotypes and allele
frequencies and their distribution in type 2 diabetes mellitus
patients with and without dyslipidemia. |
Table III
APOE genotypes and allele
frequencies and their distribution in type 2 diabetes mellitus
patients with and without dyslipidemia.
| Genotype | All subjects, n
(%) | With dyslipidemia,
n (%) | Without
dyslipidemia, n (%) |
P-valuea |
|---|
| ε2/ε2 | 5 (2.5) | 4 (4.2) | 1 (0.9) | 0.19 |
| ε2/ε3 | 19 (9.3) | 8 (8.3) | 11 (10.2) | 0.81 |
| ε2/ε4 | 3 (1.5) | 2 (2.1) | 1 (0.9) | 0.60 |
| ε3/ε3 | 162 (79.4) | 75 (78.1) | 87 (80.6) | 0.73 |
| ε3/ε4 | 15 (7.4) | 7 (7.3) | 8 (7.4) | 1 |
| ε2 | 27 (11.2) | 14 (12.4) | 13 (10.2) | 0.68 |
| ε3 | 196 (81.3) | 90 (79.6) | 106 (82.8) | 0.62 |
| ε4 | 18 (7.4) | 9(8) | 9(7) | 0.81 |
Association between APOE genotypes
with the risk of dyslipidemia
Logistic regression adjusted for age, sex, and BMI
showed no association between APOE genotypes and the risk of
dyslipidemia (Table IV). A
comparison of lipid parameters between ε3/ε3 subjects and ε4 allele
carriers in both groups revealed no significant differences in mean
values for the LDL-C, HDL-C, TG, and TC levels (Table V).
| Table IVAssociation of APOE genotypes
with the risk of dyslipidemia in type 2 diabetes mellitus
patients. |
Table IV
Association of APOE genotypes
with the risk of dyslipidemia in type 2 diabetes mellitus
patients.
| Genotype | All subjects, n
(%) | With dyslipidemia,
n (%) | Without
dyslipidemia, n (%) | Adjusted OR 95%
CIa | P-value | OR 95% CI | P-value |
|---|
| ε2/ε2 | 5 (2.5) | 4 (4.2) | 1 (0.9) | 0.28
(0.01-3.66) | 0.36 | 4.6 (0.51-42) | 0.17 |
| ε2/ε3 | 19 (9.3) | 8 (8.3) | 11 (10.2) | 1.25
(0.39-4.17) | 0.71 | 0.8 (0.32-2.2) | 0.72 |
| ε2/ε4 | 3 (1.5) | 2 (2.1) | 1 (0.9) | 1.34
(0.05-20.20) | 0.83 | 2.3 (0.21-26) | 0.5 |
| ε3/ε3 | 162 (79.4) | 75 (78.1) | 87 (80.6) | 1.00 | NA | 1.00 | NA |
| ε3/ε4 | 15 (7.4) | 7 (7.3) | 8 (7.4) | 0.67
(0.17-2.67) | 0.57 | 1.02
(0.35-2.9) | 0.98 |
| Table VComparison of the lipid profiles
between ε3/ε3 genotype subjects and ε4 allele carriersd. |
Table V
Comparison of the lipid profiles
between ε3/ε3 genotype subjects and ε4 allele carriersd.
| | With
dyslipidemia | Without
dyslipidemia |
|---|
| Variable | ε3/ε3 | ε4 | P-value | ε3/ε3 | ε4 | P-value |
|---|
| Age,
yearse | 62.4 (9.8) | 61.1(11) | 0.61a | 64.3 (10.3) | 57.6 (10.4) | 0.009a,b |
| Sex, female/ male,
n | 28/47 | 12/5 | 0.544b | 37/50 | 8/12 | 0.521c |
| Body mass index,
kg/m2e | 29.7(4) | 30.8 (2.7) | 0.289a | 24.9 (3.6) | 23.7 (3.3) | 0.184b |
| Total cholesterol,
mg/dle | 238.37(91) | 232.44 (77.95) | 0.84a | 160.78 (49.19) | 162.78 (66.16) | 0.93b |
| Total triglyceride,
mg/dle | 251.53
(161.56) | 251.56
(116.82) | 1a | 150.15 (62.02) | 131.67 (37.19) | 0.21b |
| High-density
lipoprotein, mg/dle | 36.57 (13.93) | 36.44 (8.38 ) | 0.97a | 49.01 (18.42) | 48.33 (14.44) | 0.90b |
| Low-density
lipoprotein, mg/dle | 164.23 (56.55) | 136 (46.81) | 0.12a | 90.89 (35.17) | 114.44 (68.68) | 0.34b |
Allelic and genotypic frequency of
APOE promoter variants and the risk of dyslipidemia
The genotypes and alleles distributions of the three
promoter variants (rs769446, rs449647, and rs405509) are shown in
Table VI. No deviations from the
expected HW frequencies were found in both groups (P>0.05). The
genotype distribution and allele frequency of the three promoter
variants were not statistically different between the two studied
groups (P>0.05; Table VI).
Moreover, no statistical differences were observed in the genotype
frequencies of the three APOE promoter variants stratified
according to the ε2, ε3/ε3, and ε4 status (data not shown).
| Table VIGenotypes and alleles frequencies of
APOE promoter variants among type 2 diabetes mellitus
patients with and without dyslipidemia. |
Table VI
Genotypes and alleles frequencies of
APOE promoter variants among type 2 diabetes mellitus
patients with and without dyslipidemia.
| SNP rs | Genotype/
allele | Total, n (%) | With dyslipidemia,
n (%) | Without
dyslipidemia, n (%) | Odds
ratioa | 95% Confidence
intervala | P-value |
|---|
| rs769446 | TT | 181 (88.7) | 87(91) | 94(87) | 1.00 | | 0.14 |
| | TC | 23 (11.3) | 9(9) | 14(13) | 0.44 | (0.14-1.34) | |
| | T | 385(94) | 183(95) | 202(94) | 1.00 | | 0.14 |
| | C | 23(6) | 9(5) | 14(6) | 0.44 | (0.14-1.34) | |
| rs449647 | AA | 84 (41.2) | 42(44) | 42(39) | 1.00 | | 0.07 |
| | AT | 92 (45.1) | 41(43) | 51(47) | 0.42 | (0.19-0.92) | |
| | TT | 28 (13.7) | 13(14) | 15(14) | 0.81 | (0.29-2.30) | |
| | A | 260 (63.7) | 125(65) | 135(62) | | | 0.28 |
| | T | 148 (36.3) | 67(35) | 81(38) | 0.76 | (0.46-1.25) | |
| rs405509 | GG | 57 (27.9) | 26(27) | 31(29) | 1.00 | | 0.58 |
| | GT | 99 (48.5) | 48(50) | 51(47) | 0.96 | (0.42-2.19) | |
| | TT | 48 (23.5) | 22(23) | 26(24) | 1.51 | (0.56-4.09) | |
| | G | 213 (52.2) | 100(52) | 113(52) | 1.00 | | 0.44 |
| | T | 195 (47.8) | 92(48) | 103(48) | 1.22 | (0.74-2.00) | |
DNA methylation level of the APOE
promoter region
A sub group of 119 patients was included to study
the methylation pattern among T2D patients with and without
dyslipidemia. The clinical characteristics of this subgroup (n=119;
58 with dyslipidemia and 61 without dyslipidemia) are described in
Table SII. The mean BMI was higher
in patients with dyslipidemia (30.2±3.8 kg/m2) compared
with those without dyslipidemia (24.8±3.6 kg/m2)
(P<0.05). The lipid profiles including TC, TG, and LDL were also
significantly higher in patients with dyslipidemia compared with
those without dyslipidemia (P<0.05; Table SII). Percent methylation for each
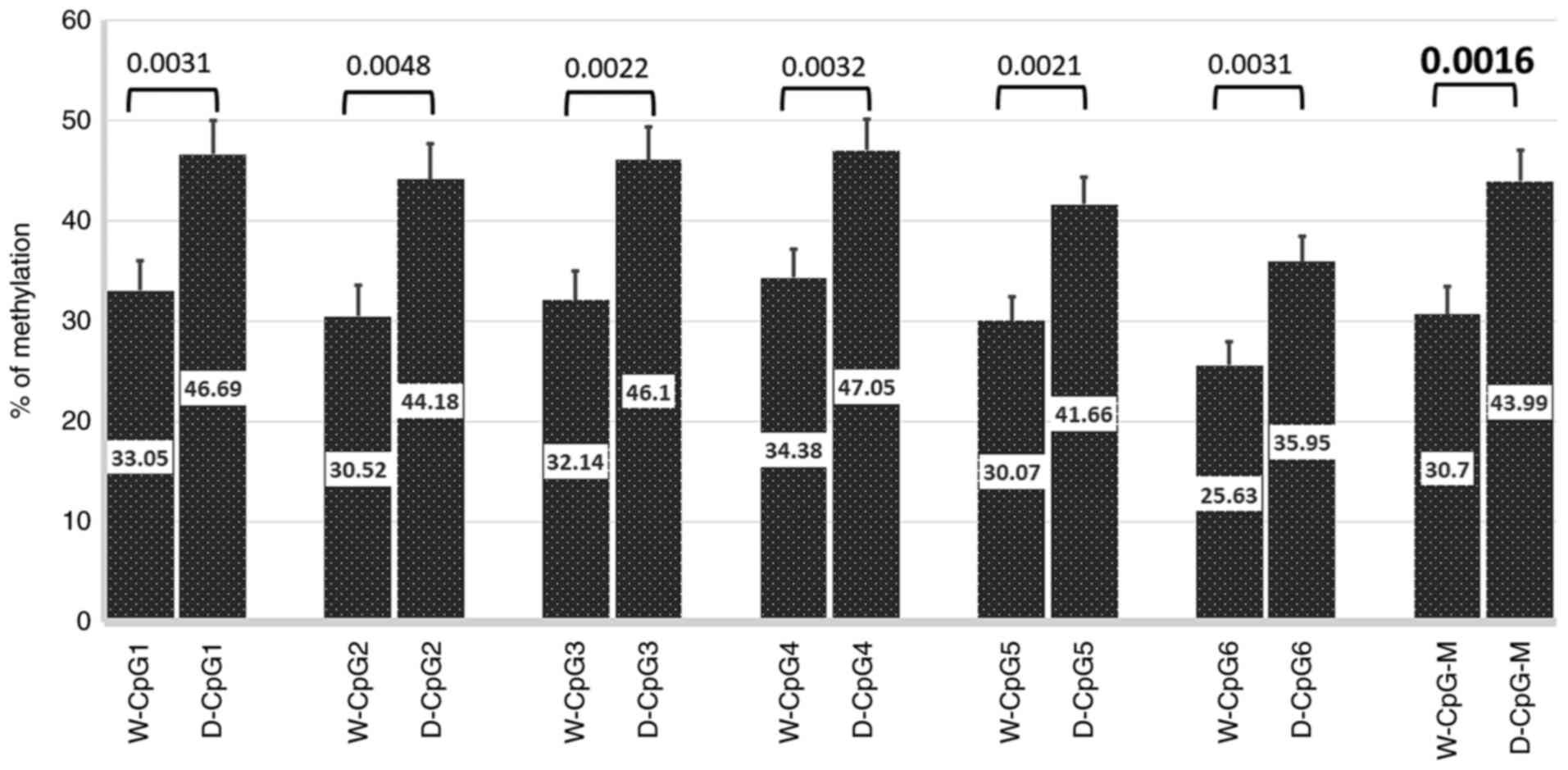
CpG (1-6)
across the APOE promoter was compared between diabetic
patients with and without dyslipidemia. The percent methylation
level at any CpG site was significantly higher in patients with
dyslipidemia compared with those without dyslipidemia: CpG1,
P=0.0031; CpG2, P=0.0048; CpG3, P=0.0022; CpG4, P=0.0032; CpG 5,
P=0.0021, CpG6, P=0.0031, mean CpG (1-6)
methylation, P=0.0016 (Fig. 2).
The relationship between mean methylation levels and
the risk of dyslipidemia was analyzed using a logistic regression
model adjusted for potential confounding factors. the analysis
showed a significant association of methylation level at the CpGs
(1-6)
with increased risk of dyslipidemia odds ratio (OR)=1.038; 95%
confidence interval (CI)=1.012-1.064); P=0.004 (adjusted for age,
sex, BMI, HDL, LDL levels, and ε4 allele, Table VII). Moreover, LDL levels were
revealed to be an independent risk factor for dyslipidemia
development (OR=1.025; 95% CI=1.013-1.038); P<0.001 (adjusted
for age, sex, BMI, HDL, and ε4 allele). BMI was positively
associated with the risk of dyslipidemia (OR=1.458; 95%
CI=1.212-1.754); P<0.001 (adjusted for age, sex, LDL, HDL, and
ε4 allele).
| Table VIILogistic regression analysis of the
association between each variable and risk of dyslipidemia. |
Table VII
Logistic regression analysis of the
association between each variable and risk of dyslipidemia.
| Variable | Odds ration | 95% confidence
interval | Adjusted
P-value |
|---|
| Mean CpG(1-6) | 1.038 | 1.012-1.064 | 0.004b |
| Age | 1.049 | 0.987-1.115 | 0.126 |
| Sex | 1.466 | 0.463-4.641 | 0.515 |
| Body mass
index | 1.458 | 1.212-1.754 | 0.00a |
| High-density
lipoprotein | 1 | 0.954-1.047 | 0.988 |
| Low-density
lipoprotein | 1.025 | 1.013-1.038 | 0.00a |
| ε4 carrier | 1.63 | 0.376-7.067 | 0.514 |
Stratification of LDL and BMI mean values by sex and
age showed no significant differences in each group with and
without dyslipidemia (Table
SIII).
Discussion
It is now well established that the APOE gene
plays an important role in lipid metabolism and its genetic
variations are associated with cognitive function, diabetes and
cardiovascular disease (20,21).
To the best of our knowledge, the effect of APOE
polymorphisms and methylation profile on diabetic dyslipidemia has
never been studied in the Palestinian population. First, we
investigated the relationship between the APOE polymorphisms
and the risk of dyslipidemia in T2D patients. The current study
showed that there was no significant difference in the distribution
of ε2, ε3, and ε4 alleles among T2D patients with and without
dyslipidemia. In all T2D subjects, the ε3 allele was the most
common (81.3%), followed by the ε2 allele (11.2%) and then the ε4
allele (7.4%), which is different from that reported in Turkish
(22), Egyptian (23) and Saudi diabetic patients (24). The differences in APOE allele
frequency in our Palestinian population could indicate differences
in disease risk. A Palestinian study conducted on healthy men
(n=140) reported that the overall prevalence of dyslipidemia was
66.4% and hypo HDL was the most prevalent type of dyslipidemia
(59.3%), which was followed by hypertriglyceridemia (20%) (25). In contrast, dyslipidemia prevalence
was 78.7% in men and 80.4% in women in Turkish adults (26), 33% in Saudi adults (27), and 36.8% in Egyptian adults
(28), which may reflect different
genetic predispositions, socioeconomic status, and lifestyles of
the studied subjects. Another Palestinian study revealed that hypo
HDL was highly prevalent among obese and normal-weight adults
indicating that factors other than BMI and central obesity could
influence hypo HDL in Palestinians; the study revealed a
significant association of hypo HDL with marital status, tobacco
smoking, and occupational exposure to pesticides (29).
Although the three ApoE isoforms differ in their
binding affinity to LDL receptors resulting in differences in
clearance and uptake of lipoproteins and lipid levels (30), our results showed no differences in
serum lipid levels (TC, TG, LDL-C, and HDL-C) between the ε4 and
ε3/ε3 carriers in both groups with and without dyslipidemia. In
agreement with these results, Al-Shammari et al (31) reported no association between
APOE allelic patterns and the blood lipid levels in healthy
Kuwaiti Arab subjects and patients with combined hyperlipidemia. A
recent study conducted in 2021 on Indonesian T2D patients reported
no association between TC, TG, and LDL-C levels with APOE
polymorphisms, but showed that ε2 and ε4 carriers had lower levels
of HDL-C (32). Other studies
revealed that APOE polymorphisms were associated with serum
lipid levels in healthy controls but not in T2D patients (33,34). A
meta-analysis of 16 studies revealed an association of APOE
polymorphisms with the levels of TC, TG, HDL-C, and LDL-C (35). However, the inconsistent results may
be attributed to the differences in the included populations, the
sample sizes, genotyping methods, and other risk factors. In the
present study, we used amplicon based NGS assay, which provides DNA
sequences with minimum genotyping error compared with other
traditional genotyping methods. DNA was sequenced with a quality
score of >20, representing an error rate of 1 in 100 with a
corresponding call accuracy of 99%. Moreover, no significant
associations were found between the promoter variants rs769446,
rs449647, and rs405509 with the risk of dyslipidemia or changes in
the lipid profile. Notably, among the ε3/ε3 subjects who had
rs405509 GG genotype, there was a trend towards higher LDL and
triglyceride levels than in TT carries, but this did not reach
statistical significance (data not shown). These findings were
previously described in the general population (36). Hence, future studies with larger
sample sizes including non-diabetic subjects are warranted.
No previous studies have investigated APOE
methylation in relation to diabetic dyslipidemia in Palestinians,
to the best of our knowledge. Therefore, to clarify the
contribution of APOE promoter methylation to the risk of
diabetic dyslipidemia, we used 119 available DNA samples and
evaluated the levels of APOE promoter methylation in the T2D
patients with and without dyslipidemia. A total of six CpG sites
were identified in the APOE promoter within the area
44905755-44906078 on chromosome 19 which were differentially
methylated between the studied groups. We observed that the
APOE promoter was significantly hypermethylated in T2D
patients with dyslipidemia compared with those without dyslipidemia
for the six CpG sites. These results indicated that DNA methylation
changes in the APOE promoter may be involved in the
progression of diabetic dyslipidemia. These results could be
attributed to the fact that the APOE promoter region has
important epigenetic regulatory functions in which hypermethylation
of this region could modify gene expression and consequently
influence blood lipid levels and participate in the pathogenesis of
dyslipidemia. However, the function of this candidate element
should be tested in vivo to validate its transcriptional
regulatory activity and to investigate its effect on the ApoE
level. A recent study revealed a negative correlation between the
levels of total APOE RNA and DNA methylation at the
APOE CGI in the frontal lobe tissues (37). Another APOE genotyping and
sequencing study conducted on 10,369 individuals revealed that
dementia risk increased with decreasing ApoE levels caused by rare
genetic variations other than the common ε2/ε3/ε4 polymorphisms
(38).
In addition, a review study reported that the common
and rare APOE variants alongside environmental factors and
epigenetics are associated with variations in lipids and
lipoprotein levels affecting the clinical presentation of
dyslipidemia (39). Together, these
results indicated that molecular screening of the APOE gene
and study of epigenetic variations are crucial to understanding the
implications of the APOE variants in each of these diseases,
which would be possible using high throughput sequencing, i.e NGS.
In the present study, logistic regression analysis revealed that
higher DNA methylation levels at the six CpGs in the promoter
region of APOE increases the odds of dyslipidemia
independently of the APOE ε4 carrier. Karlsson et al
(40) also reported that
APOE allelic variation and increased methylation levels in
the promoter region may act independently to increase the risk of
dementia. Another case-control study showed that a higher DNA
methylation level at two CpGs in the APOE promoter was
independently associated with atherosclerotic cerebral infarction
(41). In contrast to our results,
Mur et al (42) showed that
DNA methylation levels at five CpGs in the APOE promoter
region were higher in ε4 carriers compared to ε3 carriers.
It is well known that the endogenous pattern of DNA
methylation is influenced by environmental factors such as
environmental pollutants, and the social environment including
early life stressors (maternal stress) that have a marked effect on
DNA methylation during fetal development and throughout life. These
epigenetic changes may affect neuronal structure and function and
thus an organism's health behaviors (43,44)
which are however beyond the scope of this study. In the current
study we confirmed the known effect of age on methylation levels, a
group of non-diabetic healthy individuals (n=58) of younger age
(mean age ± SD, 45.6±8.5) tended to have lower methylation levels
at the six analyzed CpG sites (mean ± SD, 11.595±7.944) (data not
shown).
However, additional detailed study is required to
investigate the association between DNA methylation and age between
the two sex subgroups. Furthermore, LDL levels and BMI were
revealed as independent risk factors for dyslipidemia, which
confirmed that the progression of diabetic dyslipidemia is a
consequence of genetic and other risk factors that affect lipid
levels such as age, smoking, and a high fat intake (45).
In addition to the small sample size, some
limitations should be addressed in our study. First, the selected
six CpGs in the APOE promoter region may not represent the
methylation pattern of the whole APOE gene, other CpGs may
also be related to the risk of dyslipidemia. Second, confounding
factors such as smoking, energy intake, folic acid supplementation,
dyslipidemia/antidiabetic medications, and family history might
affect DNA methylation patterns. A recent study reported that
statin use was an independent factor of higher ABCG1
methylation (at the cg06500161 site) and this was correlated with
the differential expression of genes involved in both lipid
metabolism and glycemic pathways (46). Unfortunately, no information on the
treatment or family history of diabetes and dyslipidemia was
available for all study participants; therefore we could not
include these confounding factors in our statistical analysis.
A study conducted on stunted growth revealed a
significant association between low dietary intakes (lower intakes
of protein and carbohydrate) and increased global DNA methylation
(47). Generally, the DNA
methylation process is catalyzed by DNA methyltransferases enzymes
(DNMTs) in which the methyl group is transferred from
S-adenosylmethionine and incorporated into carbon 5 of the cytosine
residue followed by a release of s-adenosylhomocysteine (48). Therefore, any genetic deletion or
silencing of these enzymes will induce passive demethylation of the
CpG sites in gene promoters and subsequently aberrant gene
expression (48). Furthermore, some
dietary intakes that act as methyl donors may influence DNA
methylation either by inhibiting DNMTs enzymes or by changing the
availability of substrates required for these enzymatic reactions
(49).
In conclusion, this study demonstrates that
APOE ε genotypes and the promoter variants (rs769446,
rs449647, and rs405509) are not associated with an increased risk
of diabetic dyslipidemia. Our study revealed that APOE DNA
methylation levels at the six CpGs in the promoter region are
associated with diabetic dyslipidemia. Differential DNA methylation
at these loci might serve as a biomarker for diabetic dyslipidemia
and represent an interesting possibility for future research to
identify a new therapeutic target to reverse the methylation of
APOE promoter and may improve the clinical management of
diabetic dyslipidemia.
Supplementary Material
Demographic and biochemical
characteristics of study subjects (mean ranks).
Clinical and biochemical data for
participants who were included in the methylation analysis compared
to those who were not included.
Distribution of age and sex on BMI and
LDL values for participants who included in methylation
analysis
Acknowledgements
Not applicable.
Funding
Funding: This work was funded by the Deanship of Scientific
Research (Al-Quds University-Palestine, Abu Dis, Palestine).
Availability of data and materials
The original data generated using next generation
sequencing dataset has been deposited in the National Center for
Biotechnology Information Sequence Read Archive (BioProject ID:
PRJNA830334, http://www.ncbi.nlm.nih.gov/bioproject/830334; and
BioProject ID: PRJNA830330, http://www.ncbi.nlm.nih.gov/bioproject/830330). The
datasets used and/or analyzed during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
SE and AN conceived the study. SC and AN performed
the analysis. KE, ME, MG and AS performed the experiments and
curated the data. SE wrote the manuscript. SC and AN reviewed and
edited the manuscript. All authors read and approved the final
manuscript. SE and AN confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The study was conducted according to the guidelines
expressed in the Declaration of Helsinki, and written informed
consent was obtained from all enrolled participants. The study
procedure was approved by the local ethical committee at Al-Quds
University (East Jerusalem, Palestine; approval no. 71/REC).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yu CE and Foraker J: Epigenetic
considerations of the APOE gene. [corrected]. Biomol Concepts.
6:77–84. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Belloy ME, Napolioni V and Greicius MD: A
Quarter century of APOE and Alzheimer's disease: Progress to date
and the path forward. Neuron. 101:820–838. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ashiq S and Ashiq K: The association of
apolipoprotein-E (APOE) gene polymorphisms with coronary artery
disease: A systematic review and meta-analysis. Egypt J Med Hum
Genet. 22(16)2021.
|
|
4
|
Smelt AH and de Beer F: Apolipoprotein E
and familial dysbetalipoproteinemia: Clinical, biochemical, and
genetic aspects. Semin Vasc Med. 4:249–257. 2004.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Sun Y, Wei R, Yan D, Xu F, Zhang X, Zhang
B, Yimiti D, Li H, Sun H, Hu C, et al: Association between APOE
polymorphism and metabolic syndrome in Uyghur ethnic men. BMJ Open.
6(e010049)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Villeneuve S, Brisson D, Marchant NL and
Gaudet D: The potential applications of apolipoprotein E in
personalized medicine. Front Aging Neurosci. 6(154)2014.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Maitland-van der Zee AH, Stricker BH,
Klungel OH, Mantel-Teeuwisse AK, Kastelein JJ, Hofman A, Leufkens
HG, van Duijn CM and de Boer A: Adherence to and dosing of
beta-hydroxy-beta-methylglutaryl coenzyme A reductase inhibitors in
the general population differs according to apolipoprotein
E-genotypes. Pharmacogenetics. 13:219–223. 2003.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Cai C, Wen Z and Li L: The relationship
between ApoE gene polymorphism and the efficacy of statins
controlling hyperlipidemia. Am J Transl Res. 13:6772–6777.
2021.PubMed/NCBI
|
|
9
|
Zhang L, He S, Li Z, Gan X, Li S, Cheng X,
Yang N and Zheng F: Apolipoprotein E polymorphisms contribute to
statin response in Chinese ASCVD patients with dyslipidemia. Lipids
Health Dis. 18(129)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Bizzarro A, Seripa D, Acciarri A, Matera
MG, Pilotto A, Tiziano FD, Brahe C and Masullo C: The complex
interaction between APOE promoter and AD: An Italian case-control
study. Eur J Hum Genet. 17:938–945. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bekris LM, Lutz F and Yu CE: Functional
analysis of APOE locus genetic variation implicates regional
enhancers in the regulation of both TOMM40 and APOE. J Hum Genet.
57:18–25. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ciceri F, Rotllant D and Maes T:
Understanding epigenetic alterations in Alzheimer's and Parkinson's
disease: Towards targeted biomarkers and therapies. Curr Pharm Des.
23:839–857. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bae MG, Kim JY and Choi JK: Frequent
hypermethylation of orphan CpG islands with enhancer activity in
cancer. BMC Med Genomics. 9 (Suppl 1)(S38)2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Kumar R, Nandhini LP, Kamalanathan S,
Sahoo J and Vivekanadan M: Evidence for current diagnostic criteria
of diabetes mellitus. World J Diabetes. 7:396–405. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jellinger PS, Dickey RA, Ganda OP, Mehta
AE, Nguyen TT, Rodbard HW, Seibel JA, Shepherd MD and Smith DA:
AACE Lipid Guidelines Committee. The American Association of
Clinical Endocrinologists. AACE medical guidelines for clinical
practice for the diagnosis and treatment of dyslipidemia and
prevention of atherogenesis. Endocr Pract. 6:162–213.
2000.PubMed/NCBI
|
|
16
|
Goodyear MD, Krleza-Jeric K and Lemmens T:
The declaration of Helsinki. BMJ. 335:624–625. 2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Krishnaveni P and Gowda VM: Assessing the
validity of friedewald's formula and anandraja's formula for serum
LDL-cholesterol calculation. J Clin Diagn Res. 9:BC01–BC04.
2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ereqat S, Cauchi S, Eweidat K, Elqadi M
and Nasereddin A: Estrogen receptor 1 gene polymorphisms (PvuII and
XbaI) are associated with type 2 diabetes in Palestinian women.
PeerJ. 7(e7164)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Shao Y, Shaw M, Todd K, Khrestian M,
D'Aleo G, Barnard PJ, Zahratka J, Pillai J, Yu CE, Keene CD, et al:
DNA methylation of TOMM40-APOE-APOC2 in Alzheimer's disease. J Hum
Genet. 63:459–471. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Shatwan IM, Winther KH, Ellahi B, Elwood
P, Ben-Shlomo Y, Givens I, Rayman MP, Lovegrove JA and Vimaleswaran
KS: Association of apolipoprotein E gene polymorphisms with blood
lipids and their interaction with dietary factors. Lipids Health
Dis. 17(98)2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Satizabal CL, Samieri C, Davis-Plourde KL,
Voetsch B, Aparicio HJ, Pase MP, Romero JR, Helmer C, Vasan RS,
Kase CS, et al: APOE and the association of fatty acids with the
risk of stroke, coronary heart disease, and mortality. Stroke.
49:2822–2829. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Duman BS, Oztürk M, Yilmazer S and Hatemi
H: Apolipoprotein E polymorphism in Turkish subjects with Type 2
diabetes mellitus: Allele frequency and relation to serum lipid
concentrations. Diabetes Nutr Metab. 17:267–274. 2004.PubMed/NCBI
|
|
23
|
Galal AA, Abd Elmajeed AA, Elbaz RA, Wafa
AM and Elshazli RM: Association of apolipoprotein E gene
polymorphism with the risk of T2DM and obesity among Egyptian
subjects. Gene. 769(145223)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Alharbi KK, Khan IA and Syed R:
Association of apolipoprotein E polymorphism with type 2 diabetes
mellitus in a Saudi population. DNA Cell Biol. 33:637–641.
2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ali I, Kharma A, Samara M, Odeh S, Jaradat
N, Zaid AN and Ahmad MA: Prevalence of dyslipidemia in undiagnosed
palestinian men: A cross-sectional study. J Lipids.
2019(3473042)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Bayram F, Kocer D, Gundogan K, Kaya A,
Demir O, Coskun R, Sabuncu T, Karaman A, Cesur M, Rizzo M, et al:
Prevalence of dyslipidemia and associated risk factors in Turkish
adults. J Clin Lipidol. 8:206–216. 2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Alzahrani GS, Aljehani SM and Al-Johani
JJ: Risk factors of dyslipidemia among Saudi population, 2017.
Egypt J Hosp Med. 71:2262–2265. 2018.
|
|
28
|
Reda A, Ragy H, Saeed K and Alhussaini MA:
A semi-systematic review on hypertension and dyslipidemia care in
Egypt-highlighting evidence gaps and recommendations for better
patient outcomes. J Egypt Public Health Assoc.
96(32)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Damiri B, Aghbar A, Alkhdour S and Arafat
Y: Characterization and prevalence of metabolic syndrome among
overweight and obese young Palestinian students at An-Najah
National University. Diabetes Metab Syndr. 12:343–348.
2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Mahley RW: Central nervous system
lipoproteins: ApoE and regulation of cholesterol metabolism.
Arterioscler Thromb Vasc Biol. 36:1305–1315. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Al-Shammari S, Fatania H, Al-Radwan R and
Akanji AO: Apolipoprotein E polymorphism and lipoprotein levels in
a Gulf Arab population in Kuwait: A pilot study. Ann Saudi Med.
24:361–364. 2004.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Maratni NP, Saraswati MR, Dewi NN, Yasa I,
Eka Widyadharma IP, Putra IB and Suastika K: Association of
apolipoprotein E gene polymorphism with lipid profile and ischemic
stroke risk in type 2 diabetes mellitus patients. J Nutr Metab.
2021(5527736)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Srirojnopkun C, Kietrungwilaikul K,
Boonsong K, Thongpoonkaew J and Jeenduang N: Association of APOE
and CETP TaqIB polymorphisms with type 2 diabetes mellitus. Arch
Med Res. 49:479–485. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Seo JY, Youn BJ, Cheong HS and Shin HD:
Association of APOE genotype with lipid profiles and type 2
diabetes mellitus in a Korean population. Genes Genomics.
43:725–735. 2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhou T, Li H, Zhong H, Zhong Z and Lin S:
Association of apoE gene polymorphisms with lipid metabolism in
renal diseases. Afr Health Sci. 20:1368–1381. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Radwan ZH, Wang X, Waqar F, Pirim D,
Niemsiri V, Hokanson JE, Hamman RF, Bunker CH, Barmada MM, Demirci
FY and Kamboh MI: Comprehensive evaluation of the association of
APOE genetic variation with plasma lipoprotein traits in U.S.
whites and African blacks. PLoS One. 9(e114618)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Lee EG, Tulloch J, Chen S, Leong L, Saxton
AD, Kraemer B, Darvas M, Keene CD, Shutes-David A, Todd K, et al:
Redefining transcriptional regulation of the APOE gene and its
association with Alzheimer's disease. PLoS One.
15(e0227667)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Rasmussen KL, Tybjaerg-Hansen A,
Nordestgaard BG and Frikke-Schmidt R: APOE and
dementia-resequencing and genotyping in 105,597 individuals.
Alzheimers Dement. 16:1624–1637. 2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Khalil YA, Rabès JP, Boileau C and Varret
M: APOE gene variants in primary dyslipidemia. Atherosclerosis.
328:11–22. 2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Karlsson IK, Ploner A, Wang Y, Gatz M,
Pedersen NL and Hägg S: Apolipoprotein E DNA methylation and
late-life disease. Int J Epidemiol. 47:899–907. 2018.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhang H, Zhao X, Wang C, Du R, Wang X, Fu
J and Sun Q: A preliminary study of the association between
apolipoprotein E promoter methylation and atherosclerotic cerebral
infarction. J Stroke Cerebrovasc Dis. 28:1056–1061. 2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Mur J, McCartney DL, Walker RM, Campbell
A, Bermingham ML, Morris SW, Porteous DJ, McIntosh AM, Deary IJ,
Evans KL and Marioni RE: DNA methylation in APOE: The relationship
with Alzheimer's and with cardiovascular health. Alzheimers Dement
(NY). 6(e12026)2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Mitchell C, Schneper LM and Notterman DA:
DNA methylation, early life environment, and health outcomes.
Pediatr Res. 79:212–219. 2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Cacabelos R and Torrellas C: Epigenetics
of aging and Alzheimer's disease: Implications for pharmacogenomics
and drug response. Int J Mol Sci. 16:30483–30543. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chaudhary R, Likidlilid A, Peerapatdit T,
Tresukosol D, Srisuma S, Ratanamaneechat S and Sriratanasathavorn
C: Apolipoprotein E gene polymorphism: Effects on plasma lipids and
risk of type 2 diabetes and coronary artery disease. Cardiovasc
Diabetol. 11(36)2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Liu Y, Shen Y, Guo T, Parnell LD,
Westerman KE, Smith CE, Ordovas JM and Lai CQ: Statin use
associates with risk of type 2 diabetes via epigenetic patterns at
ABCG1. Front Genet. 11(622)2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Iqbal MS, Rahman S, Haque MA, Bhuyan MJ,
Faruque ASG and Ahmed T: Lower intakes of protein, carbohydrate,
and energy are associated with increased global DNA methylation in
2- to 3-year-old urban slum children in Bangladesh. Matern Child
Nutr. 15(e12815)2019.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Hervouet E, Peixoto P, Delage-Mourroux R,
Boyer-Guittaut M and Cartron PF: Specific or not specific
recruitment of DNMTs for DNA methylation, an epigenetic dilemma.
Clin Epigenetics. 10(17)2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Mahmoud AM and Ali MM: Methyl donor
micronutrients that modify DNA methylation and cancer outcome.
Nutrients. 11(608)2019.PubMed/NCBI View Article : Google Scholar
|