Introduction
Generally, sepsis, a major public health burden
worldwide, is caused by an unbalanced response from the host to an
infection (1,2). Organ failure is a hallmark of sepsis,
and it is related to high morbidity and mortality during sepsis
(3,4). Among patients with sepsis, ~40% are at
risk of developing acute lung injury (ALI) (5). Sepsis can trigger lung cell apoptosis,
leading to the progression of ALI (6,7).
Although numerous studies investigated ALI induced by sepsis, their
outcomes remain unclear. Thus, there is an urgent need to explore
novel strategies for the treatment of sepsis-induced ALI.
Polydatin (also known as
resveratrol-3-O-β-mono-D-glucoside), a natural polyphenol compound,
was extracted from the roots of Polygonum cuspidatum Siebold
& Zucc. (8). Polydatin was
shown to exhibit anti-inflammatory effects in multiple diseases.
For example, polydatin downregulated IL-17 expression in activated
human peripheral blood monocytes (9). Lv et al (10) found that polydatin could attenuate
spinal cord injury by inhibiting microglial apoptosis (10). Moreover, polydatin relieved
sepsis-induced lung injury through upregulation of HO-1(11). However, the mechanism by which
polydatin regulates the development of sepsis-induced ALI remains
unclear.
Spi-B, a transcription factor that belongs to the
E26 transformation-specific (ETS) family, is highly expressed in
plasmacytoid dendritic cells (pDCs) (12). Silencing of Spi-B suppressed pDC
generation from CD34+ progenitor cells (13). In addition, Spi-B-knockout mice were
reported to have defects in T cell-dependent humoral immune
responses and activation of B-cells (14). The results from the aforementioned
studies revealed that Spi-B acts as a crucial modulator in
inflammation and immune responses. Nevertheless, the relation
between polydatin and Spi-B in sepsis-induced ALI remains unclear.
Conversely, NF-κB and PI3K/Akt signaling are crucial mediators in
sepsis-induced ALI (15,16), and Spi-B can activate NF-κB and
PI3K/Akt signaling (17,18). Polydatin was reported to inactivate
NF-κB and PI3K/Akt signaling (8,19).
Therefore, the current study aimed to explore the relationship
between polydatin and Spi-B in sepsis-induced ALI. The results of
the present study may provide novel insights into exploring
strategies for the treatment of sepsis-induced ALI.
Materials and methods
Cell culture, treatment, and
transfection
Pulmonary microvascular endothelial cells (PMVECs;
Pricella) and 293T cells (American Type Culture Collection) were
maintained in DMEM supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37˚C with 5% CO2. PMVECs were
transfected with pcDNA3.1 [negative control (NC)] or pcDNA3.1-Spi-B
[Spi-B overexpression (OE)] using Lipofectamine™ 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) for 6 h and cultured for a further
18 h at 37˚C. pcDNA3.1-NC and Spi-B OE were obtained from Shanghai
GenePharma Co., Ltd.
To mimic sepsis-induced ALI in vitro, PMVECs
were treated with lipopolysaccharide (LPS; cat. no. L2630, 100
ng/ml) for 24 h. PMVECs were pretreated with polydatin (0.5 mM) for
4 h, and then exposed to LPS (100 ng/ml) for 24 h at 37˚C. LPS,
polydatin (cat. no. 15721) and dexamethasone (Dex; cat. no. D1756)
were obtained from MilliporeSigma.
ELISA
The levels of IL-6 (cat. no. H007-1-2), TNF-α (cat.
no. H052-1-2), IL-1β (cat. no. H002-1-2) and IFN-α (cat. no.
H023-1-2) in the supernatants of PMVECs or bronchoalveolar lavage
fluid (BALF) of mice were evaluated using ELISA commercial kits
(Nanjing Jiancheng Bioengineering Institute) using a microplate
reader (DR-200Bs, Diatek GmbH).
Reverse transcription-quantitative
(RT-q)PCR
The RNAiso Plus reagent (cat. no. 9018, Takara Bio,
Inc.) was used to extract total RNA from cells or tissues. An
EntiLink™ 1st Strand cDNA Synthesis Kit [cat. no. EQ003, ELK
(Wuhan) Biotechnology Co., Ltd.] was used to reverse transcribe
total RNA (1 µg) into cDNA, according to the manufacturer's
protocol. qPCR was subsequently performed using EnTurbo™ SYBR Green
PCR SuperMix [cat. no. EQ001, ELK (Wuhan) Biotechnology Co., Ltd.]
with the following thermocycling conditions: Initial denaturation
at 94˚C for 2 min; followed by 35 cycles of 94˚C for 30 sec and
55˚C for 45 sec. The levels of mRNA were quantified using the
2-ΔΔCq method (20) and
normalized to the internal reference gene β-actin. The following
primer pairs were used for qPCR: Spi-B forward,
5'-TGGGTACTTCAGGGATCCAG-3' and reverse, 5'-TGAGGCTCTTCCCTCACTGT-3';
and β-actin forward, 5'-CTGGAACGGTGAAGGTGACA' and reverse,
5'-CGGCCACATTGTGAACTTTG-3'.
Western blotting
Cells were lysed using the RIPA lysis buffer (cat.
no. P0013B, Beyotime Institute of Biotechnology) on ice, samples
were then centrifuged at 10,000 x g, for 5 min at 4˚C. Next, total
protein concentrations were quantified using a BCA kit (cat. no.
P0010, Beyotime Institute of Biotechnology). Total protein (40
µg/lane) was separated by SDS-PAGE on a 10% SDS-gel and then
transferred to PVDF membranes (cat. no. IPVH00010, MilliporeSigma).
Subsequently, the membranes were incubated overnight at 4˚C with
the following primary antibodies: Anti-PI3K (1:1,000; cat. no.
4249; CST Biological Reagents Co., Ltd.); anti-p-PI3K (1:1,000;
cat. no. 17366; CST; Biological Reagents Co., Ltd.); anti-Akt
(1:1,000; cat. no. ab8805; Abcam); anti-p-Akt (1:1,000; cat. no.
ab81283; Abcam); anti-NF-κB1 (1:1,000; cat. no. ab288751; Abcam),
anti-Spi-B (1:1,000; cat. no. ab283286; Abcam), and anti-β-actin
(1:1,000; cat. no. ab8226; Abcam). The membranes were then
incubated with HRP-conjugated secondary antibodies (1:5,000; cat.
no. ab7090; Abcam) for 1 h at room temperature. An ECL kit (cat.
no. AS1059, ASPEN) was used for visualizing the signals using
β-actin as the loading control. Image-Pro Plus version 6.0 (Media
Cybernetics, Inc.) was used for densitometry analysis.
Adeno-associated virus (AAV)
infection
For overexpression of Spi-B in animal experiments,
293T cells were transfected with pAV-TBG-Spi-B (AAV8-Spi-B; Vigene
Biosciences) for 24 h at 37˚C. The cultures containing the virus
were centrifuged at 2,000 x g for 15 min at 4˚C and the
supernatants were collected and filtered using a 0.45 µm filter.
The viral titer of AAV8-Spi-B was 7.94x1013 vg/ml.
Animal study
C57BL/6 mice (n=36; 6-8 weeks old, 18-22 g) were
obtained from Beijing Vital River Laboratory Animal Technology Co.,
Ltd., and randomly assigned into one of six groups: Sham, cecum
ligation and puncture (CLP); CLP + polydatin; CLP + Dex, CLP +
AAV8-Spi-B; and CLP + polydatin + AAV8-Spi-B (n=6/group). To
establish the sepsis-induced ALI model, 30 mice underwent CLP
surgery as previously described (21). Subsequently, mice in the CLP +
AAV8-Spi-B group were injected with 100 µl AAV-Spi-B (10 µM)
through the tail vein. Mice in the CLP + polydatin and CLP + Dex
groups were orally administrated with 50 mg/kg polydatin and 40
mg/kg Dex respectively 48 h. In addition, mice in polydatin +
AAV8-Spi-B were administrated with 100 µl AAV-Spi-B (10 µM) via the
tail vein and treated with polydatin (50 mg/kg) via gavage.
Polydatin, Dex, and AAV8-Spi-B treatments were performed 48 h
before CLP. Finally, BALFs and lung tissues of mice were collected.
Lung tissues were lysed using RIPA lysis buffer, and then ground in
a tissue grinder (KZ-II; Wuhan Servicebio Technology Co., Ltd.).
Next, samples were centrifuged at 4˚C at 12,000 x g for 10 min, and
the supernatant was then collected and used for western blot
analysis. Dex was used as a positive control. All procedures were
performed in accordance with the Guide for the Care and Use of
Laboratory Animals (22). The
Ethics Committee of Zhongshan Hospital approved the present study
(approval no. ZH20210809). The mice were euthanized using
CO2 at a displacement rate of 40% of the chamber
volume/min (CO2 flow rate, 2.5 l/min), and animal death
was confirmed by cessation of heartbeat.
Hematoxylin and eosin (H&E)
staining
Lung tissues of mice were fixed in 4%
paraformaldehyde (cat. no. AS1018, ASPEN) at room temperature
overnight and then the tissues were paraffin-embedded and sectioned
(4-µm thick). Subsequently, the sections were stained with H&E
solution (cat. no. C0105S, Beyotime Institute of Biotechnology) at
room temperature and the injury to lung tissues was observed using
a light microscope (magnification, x200).
Statistical analysis
Data are presented as the mean ± standard deviation
of three independent experimental repeats. For comparison among
multiple groups (>2 groups), a one-way ANOVA followed by a
Tukey's post hoc test was used. P<0.05 was considered to
indicate a statistically significant difference.
Results
Polydatin attenuates the symptoms of
sepsis-induced ALI in vivo
To investigate the function of polydatin in
sepsis-induced ALI in vivo, a sepsis-induced mouse ALI model
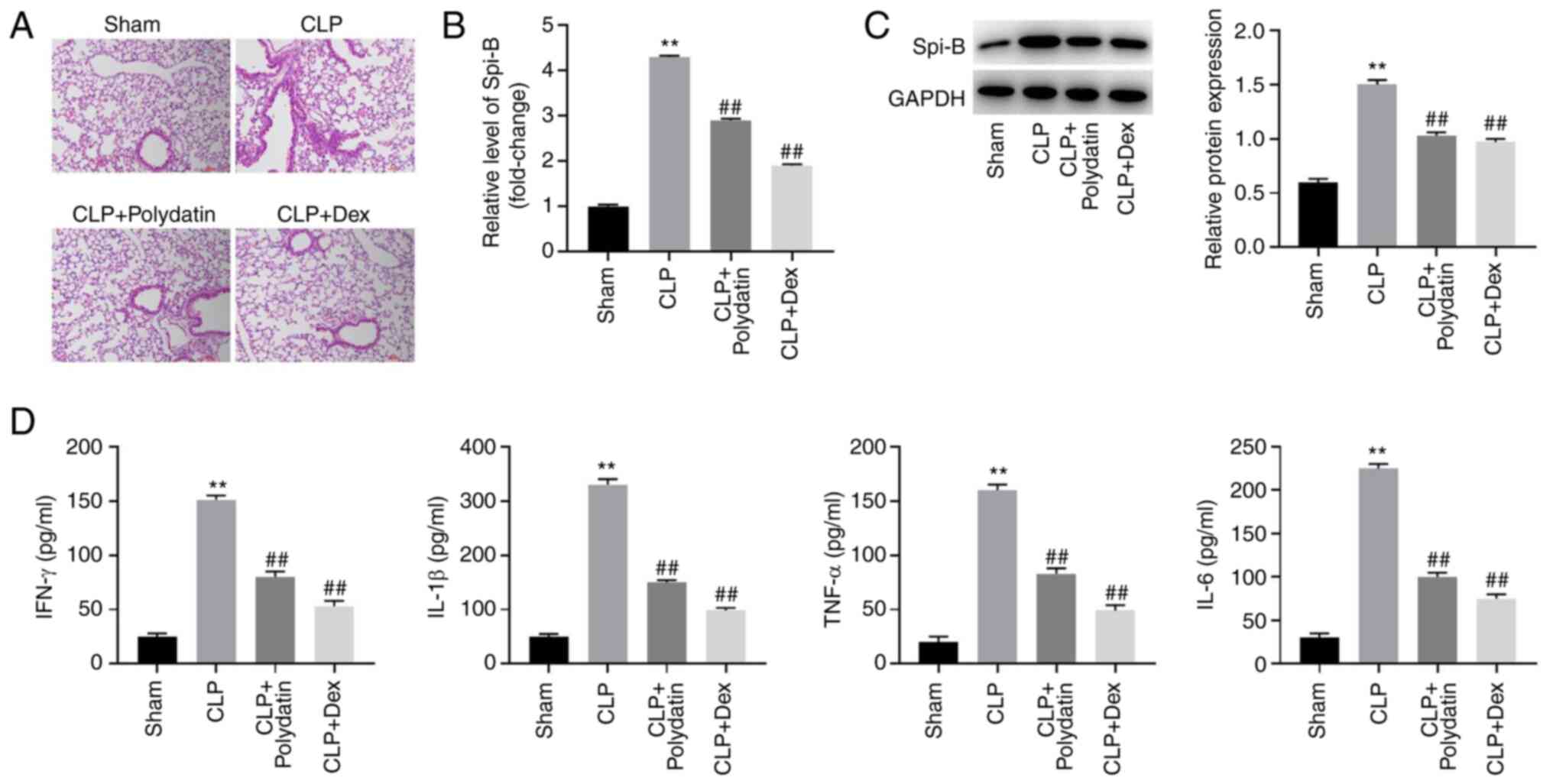
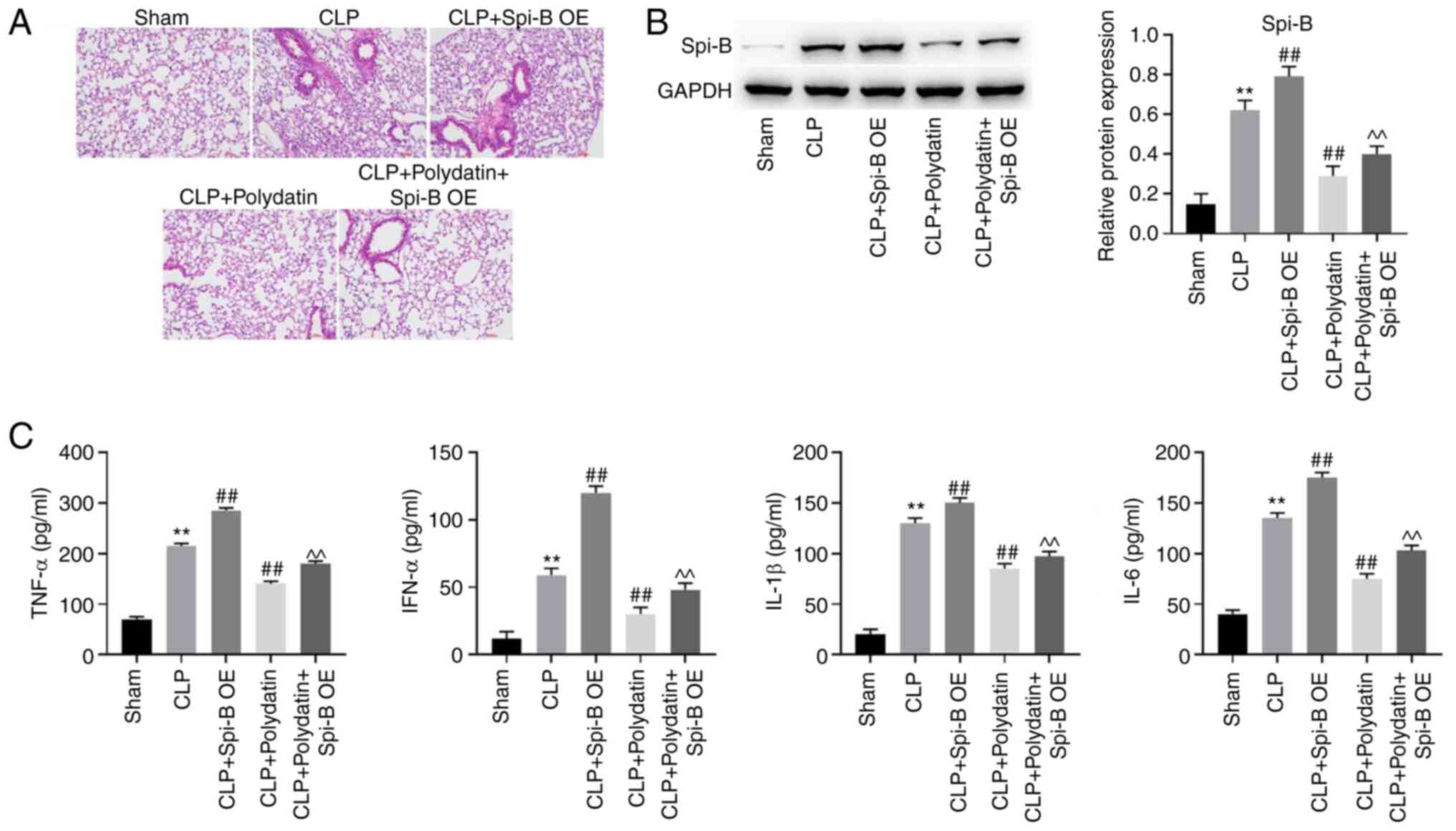
was constructed. As shown in Fig.
1A, inflammatory infiltration was observed in mice after CLP
surgery, whereas this effect was abolished via treatment with
polydatin or Dex (positive control, a drug that has been used to
treat ALI) (23). Compared to the
sham group, the mRNA levels of Spi-B were increased 4.3-fold and
the protein levels of Spi-B were increased 2.5-fold in mice
subjected to CLP (Fig. 1B and
C). However, the mRNA and protein
levels of Spi-B in the CLP + polydatin group were reduced by 33 and
31% compared to the CLP group, respectively; and the mRNA and
protein levels of Spi-B in the CLP + Dex group were reduced by 56
and 35% compared to the CLP group (Fig.
1B and C). These results showed
that the expression of Spi-B in mice was upregulated by CLP, an
effect that was partially counterbalanced following treatment with
polydatin or Dex (Fig. 1B and
C). In addition, CLP increased the
levels of IFN-α, TNF-α, IL-6, and IL-1β in the BALF of mice, while
the pro-inflammatory effect of CLP was inhibited by polydatin
(Fig. 1D). Taken together, these
results show that polydatin could attenuate the symptoms of
sepsis-induced ALI in vivo.
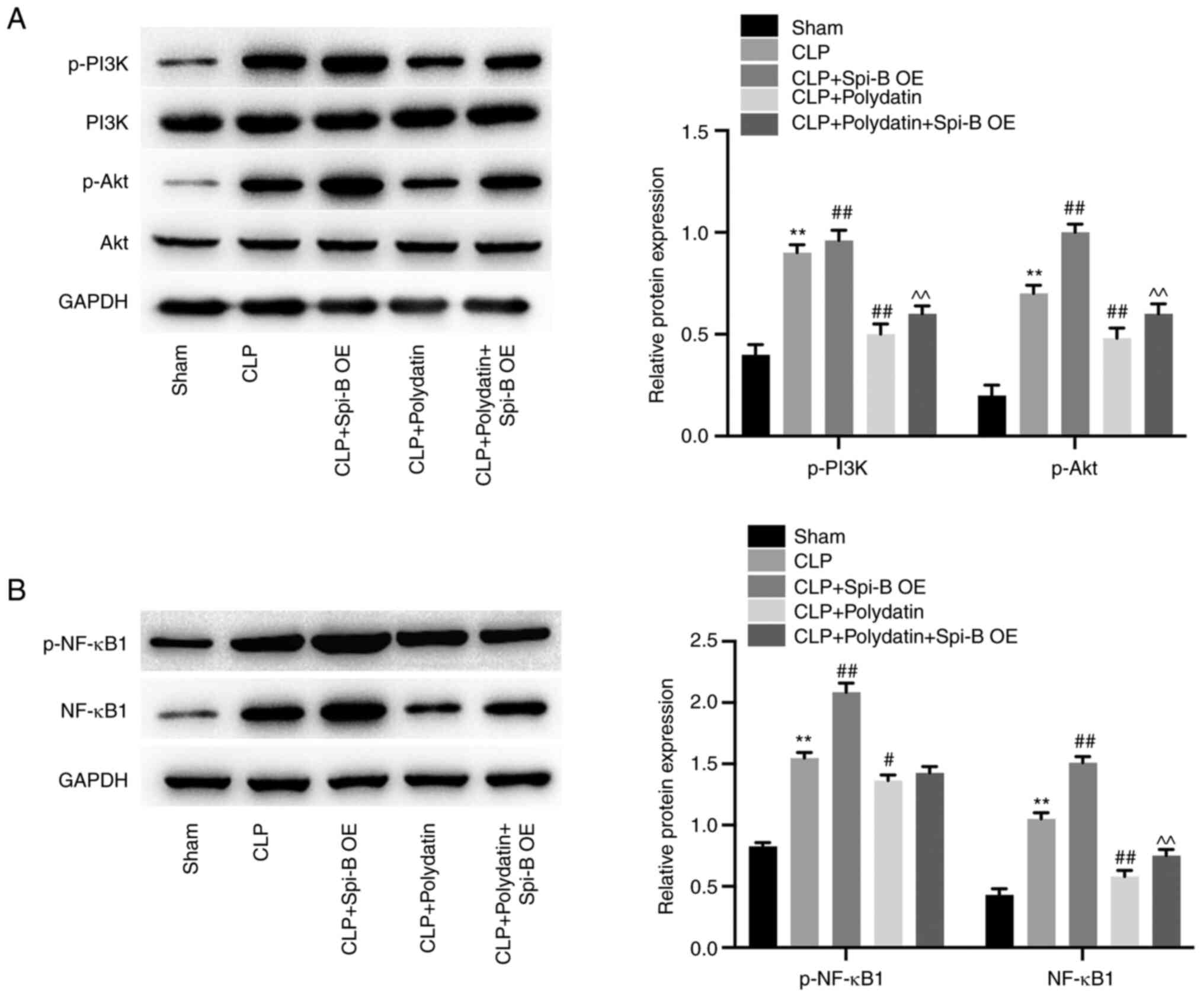
Polydatin antagonizes CLP-activated
PI3K/Akt and NF-κB signaling in the lungs of the sepsis-induced ALI
mouse model
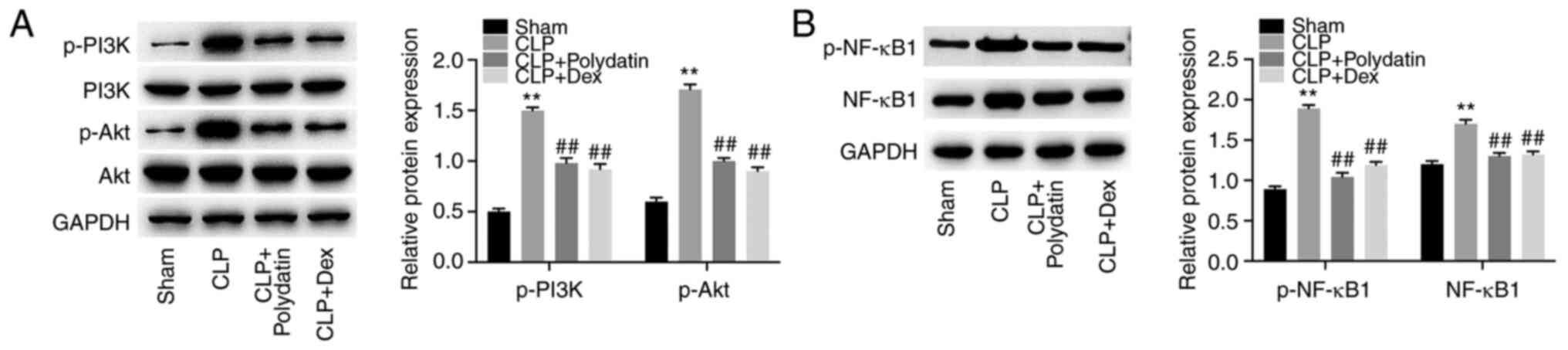
Western blotting was performed to assess the effects
of polydatin on the NF-κB and PI3K/Akt signaling pathway. CLP
increased p-PI3K, p-Akt, p-NF-κB, and NF-κB levels in mice;
however, these phenomena were partially reversed by treatment with
polydatin or Dex (Fig. 2A and
B). Polydatin was able to reverse
CLP-activated PI3K/Akt and NF-κB signaling in mice.
Polydatin antagonizes LPS-induced
inflammatory responses in PMVECs via the downregulation of
Spi-B
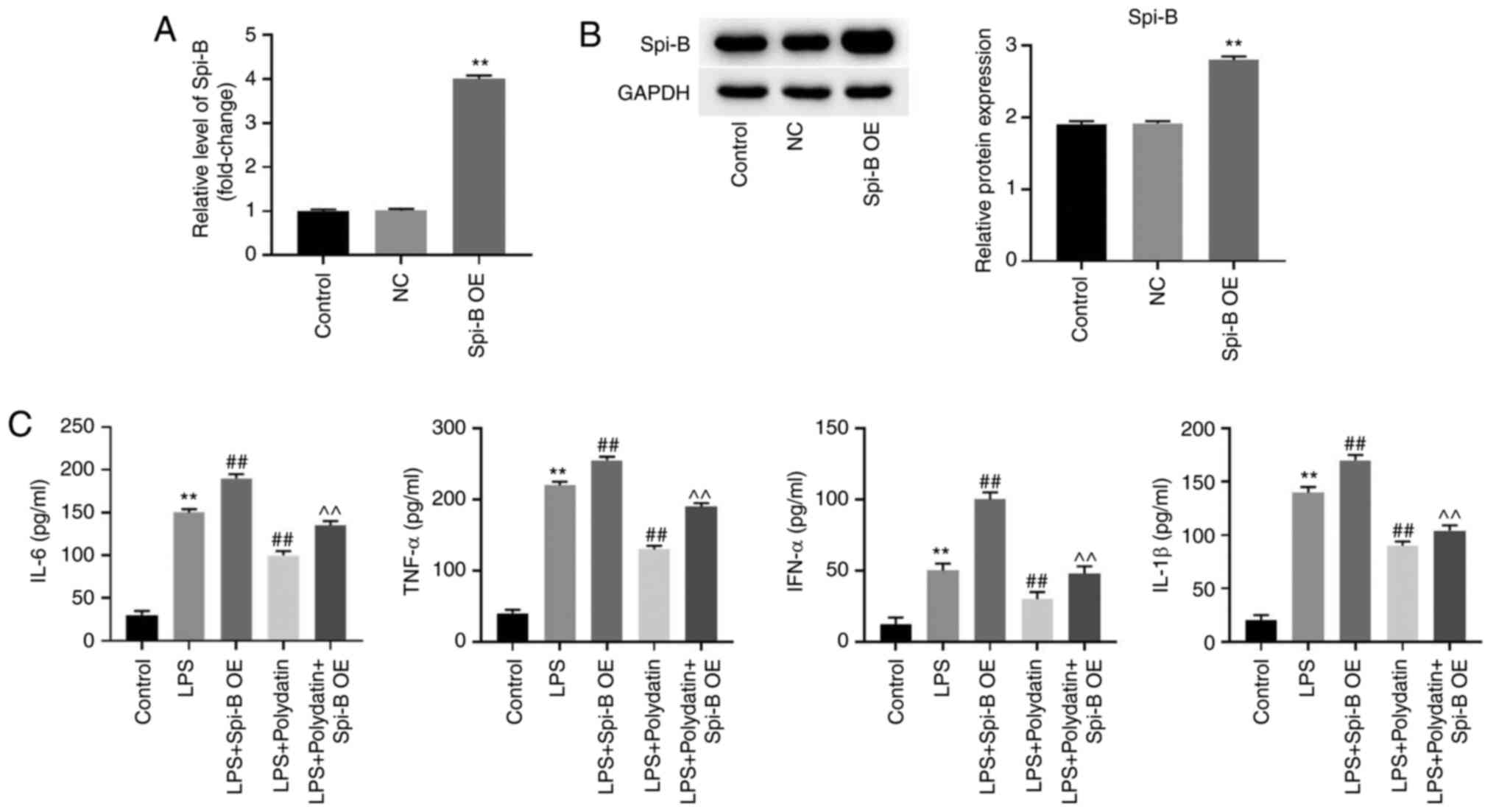
To evaluate the role of Spi-B in sepsis-induced ALI,
PMVECs were transfected with the Spi-B OE vector. The RT-qPCR data
indicated that the expression of Spi-B in PMVECs was increased
following Spi-B OE vector transfection (Fig. 3A and B). In addition, LPS-induced upregulation
of IL-6, IL-1β, TNF-α, and IFN-α in the supernatant of PMVECs was
further increased following Spi-B overexpression, while the
upregulation of cytokines was reversed by polydatin (Fig. 3C). Meanwhile, the anti-inflammatory
effect of polydatin was significantly restored by pcDNA3.1-Spi-B
transfection (Fig. 3C).
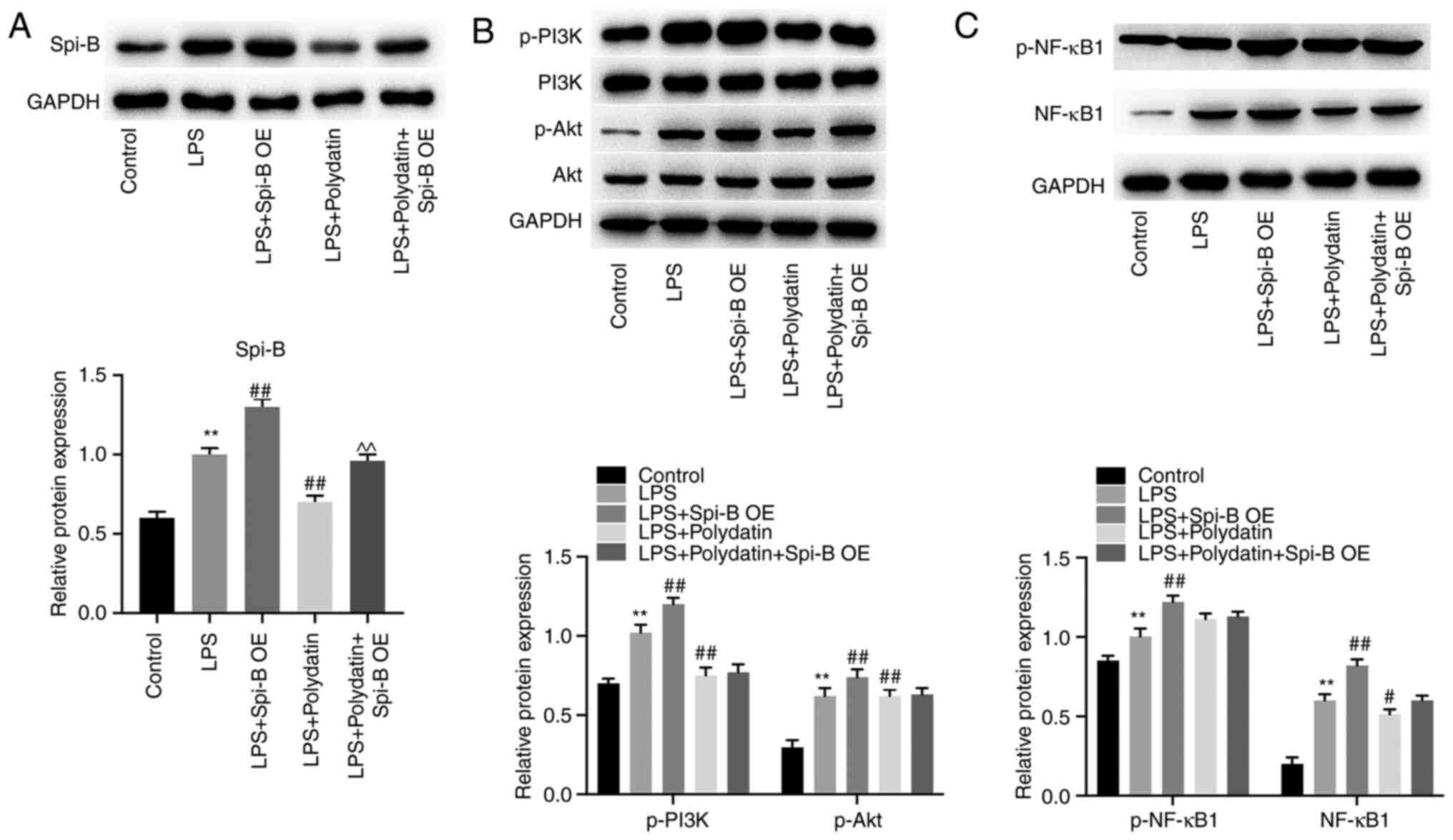
Next, to explore the relationships between
polydatin, Spi-B, PI3K/Akt, and NF-κB signaling, western blotting
was used. As shown in Fig. 4A,
LPS-induced upregulation of Spi-B in PMVECs was significantly
reversed by polydatin, while this phenomenon was partially
abolished by Spi-B OE. In addition, LPS upregulated p-Akt, p-PI3K,
and NF-κB1 levels in PMVECs; however, polydatin partially reversed
the effect of LPS on these proteins (Fig. 4B and C). Meanwhile, pcDNA3.1-Spi-B slightly
affected the levels of p-Akt, p-PI3K, and NF-κB in PMVECs
co-treated with LPS and polydatin (P>0.05, Fig. 4B and C). Together, it was shown that polydatin
reversed LPS-induced inflammatory responses in PMVECs via
downregulation of Spi-B.
Spi-B overexpression abolishes the
anti-inflammatory effect of polydatin in CLP mice
To further confirm the relation between polydatin
and Spi-B in sepsis-induced ALI in vivo, western blotting
was performed. As shown in Fig. 5A,
the protective effect of polydatin against ALI in CLP mice was
significantly abolished by Spi-B overexpression. Consistently,
Spi-B overexpression increased the levels of Spi-B in CLP mice even
when treated with polydatin (Fig.
5B). In addition, IL-1β, IL-6, TNF-α, and IFN-α levels in the
CLP mice were decreased by polydatin, while Spi-B OE transfection
reversed this phenomenon (Fig. 5C).
In summary, the anti-inflammatory effect of polydatin in CLP mice
was abolished by Spi-B overexpression.
Spi-B overexpression abolishes the
inhibitory effect of polydatin on PI3K/Akt and NF-κB signaling
The function of polydatin in PI3K/Akt and NF-κB
signaling in vivo was further explored. The data indicated
that CLP increased p-Akt, p-PI3K, p-NF-κB1, and NF-κB1 levels in
mouse lung tissues; however, polydatin treatment completely
abolished these effects (Fig. 6A
and B). Consistently, the
polydatin-induced downregulation of p-PI3K, p-Akt, and NF-κB1 in
CLP mice was partially restored by Spi-B OE. In summary, polydatin
inactivated PI3K/Akt and NF-κB signaling in CLP mice through the
downregulation of Spi-B.
Discussion
It has been previously reported that Polydatin can
mediate inflammatory responses. For example, Chen et al
(24) found that polydatin exerted
anti-inflammatory effects in LPS-induced macrophages, while
Oliviero et al (25) found
that polydatin could prevent calcium pyrophosphate crystal-induced
arthritis. IL-6, TNF-α, IL-1β, and IFN-α are pro-inflammatory
factors that play crucial roles in mediating inflammation response
in various diseases including ALI (26,27).
Tian et al (27) found that
methionine enkephalin could attenuate ALI in influenza A
virus-infected mice by decreasing IL-6, TNF-α, IL-1β, and IFN-α
levels. The present study found that polydatin notably reduced
IL-6, TNF-α, IL-1β, and IFN-α levels in LPS-treated PMVECs and in
CLP mice, suggesting that polydatin could suppress inflammatory
responses in sepsis-induced ALI. Meanwhile, it was demonstrated
that polydatin could inhibit the progression of sepsis-induced ALI
(11). The present study found that
polydatin significantly attenuated the progression of
sepsis-induced ALI in vitro and in vivo. Thus, the
present study was consistent with previous work. In addition, it
was also found that Spi-B was downregulated following polydatin
treatment in sepsis-induced ALI. Thus, it is hypothesized that
polydatin may act as an inhibitor of Spi-B in inflammatory
responses.
Spi-B is known to be a crucial mediator in immune
reactions and inflammatory responses (28,29),
and its upregulation could lead to the dysfunction of T and B cells
(30). Consistently, the current
study found that overexpression of Spi-B could reverse the
anti-inflammatory effect of polydatin. Conversely, it was shown
that the levels of cytokines secreted by macrophages were
significantly increased during the progression of ALI (31). Therefore, the results of the present
study were consistent with this previous study. In addition, Jiang
et al (32) demonstrated
that polydatin alleviated LPS-induced ALI progression by
downregulating TLR4. TLR4 upregulation could promote inflammation
by positively regulating NF-κB signaling (33,34).
Therefore, the similarity in function between Spi-B and TLR4 may
result in the similarity between the present results and the study
by Jiang et al (32).
It was reported that polydatin could prevent
1-methyl-4-phenylpyridinium-induced neurotoxicity by enhancing the
activity of myocyte enhancer factor 2D (MEF2D) and polydatin could
interact with MEF2D (35). In
addition, the transcriptional inhibitor was reported to reduce
promoter activity (36,37). Based on the aforementioned studies,
polydatin may inhibit the expression of Spi-B by interacting with a
transcriptional factor.
PI3K/Akt signaling is known to be a modulator of
cell growth (38) and it could be
activated during the occurrence of ALI (16,39).
In addition, upregulation of NF-κB signaling could lead to the
dysregulation of inflammatory reactions and this phenomenon could
result in the development of lung tissue injury (16,40).
It was demonstrated that Spi-B could promote the activation of
PI3K/Akt and NF-κB signaling during the inflammatory responses
(17,18). Consistently, the present study found
PI3K/Akt and NF-κB signaling could be activated by Spi-B
overexpression in sepsis-induced ALI. Moreover, previous studies
indicated polydatin was able to downregulate the PI3K/Akt and NF-κB
signaling pathways (11,32). Therefore, it could be suggested that
polydatin could reverse the progression of sepsis-induced ALI via
the downregulation of PI3K/Akt and NF-κB signaling.
According to Huang et al (41), polydatin prevented LPS-induced
Parkinson's disease through the mediation of the AKT/GSK3β-nuclear
factor erythroid 2-related factor 2 (Nrf2)/NF-κB axis. In addition,
the current study indicated that polydatin could alleviate
sepsis-induced ALI through inhibition of Spi-B and Spi-B OE could
activate Akt and NF-κB signaling. Thus, polydatin may inhibit NF-κB
signaling through the mediation of AKT/GSK3β-Nrf2 signaling. The
detailed function of polydatin in GSK3β-Nrf2 will be investigated
in future.
The present study has certain shortcomings such as:
i) The interaction between polydatin and transcriptional factors in
ALI induced by sepsis needs to be further investigated; and ii) the
detailed function of polydatin in GSK3β-Nrf2 requires further
investigation.
In conclusion, polydatin prevented sepsis-induced
ALI via the downregulation of Spi-B. Taken together, the present
findings revealed that polydatin alleviated sepsis-induced ALI via
downregulation of Spi-B. This research may provide novel insights
in uncovering novel strategies for the treatment of sepsis-induced
ALI.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by funding from Zhongshan
Hospital (grant no. ZH20208904).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article and are available from
FAIRsharing.org: 4TU.ResearchData;
4TU.ResearchData, DOI: 10.25504/FAIRsharing.zcveaz. The raw data is
available from https://figshare.com/s/43e3da3395e65cf0ea5c.
Authors' contributions
QL, FL, YS and YY were responsible for the
conception and design of the study. QL, FL, MX and WC performed the
experiments. ZT, MX and WC performed the analysis and
interpretation of the data. ZT wrote the manuscript. YS and YY
revised the manuscript. All authors confirm the authenticity of all
the raw data and have read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of Zhongshan Hospital (approval no. ZH20210809).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xin K, Sun J, Liu P, Ge J, Leng C and Pang
F: Expression and significance of HMGB1 in patients with sepsis and
effects on prognosis. All Life. 13:164–170. 2020.
|
|
2
|
Gao L, Shi Q, Li H, Guo Q and Yan J:
Prognostic value of baseline APACHE II score combined with uric
acid concentration for short-term clinical outcomes in patients
with sepsis. All Life. 13:416–425. 2020.
|
|
3
|
Fu D, Shen J and Shi H: Sevoflurane
suppresses oxidation-induced stress and inflammatory responses, via
promotion of Nrf2-induced antioxidant signaling. All Life.
13:131–143. 2020.
|
|
4
|
Steinhagen F, Hilbert T, Cramer N, Senzig
S, Parcina M, Bode C, Boehm O, Frede S and Klaschik S: Development
of a minimal invasive and controllable murine model to study
polymicrobial abdominal sepsis. All Life. 14:265–276. 2021.
|
|
5
|
Andrews P, Azoulay E, Antonelli M,
Brochard L, Brun-Buisson C, Dobb G, Fagon JY, Gerlach H, Groeneveld
J, Mancebo J, et al: Year in review in intensive care medicine,
2004. I. Respiratory failure, infection, and sepsis. Intensive Care
Med. 31:28–40. 2005.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hudson LD, Milberg JA, Anardi D and
Maunder RJ: Clinical risks for development of the acute respiratory
distress syndrome. Am J Respir Crit Care Med. 151 (2 Pt 1):293–301.
1995.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Herridge MS, Tansey CM, Matte A, Tomlinson
G, Diaz-Granados N, Cooper A, Guest CB, Mazer CD, Mehta S, Stewart
TE, et al: Functional disability 5 years after acute respiratory
distress syndrome. N Engl J Med. 364:1293–1304. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Xiong Q, Yan Z, Liang J, Yuan J, Chen X,
Zhou L, Hu Y, Wu J, Jing Y, Zhang Q, et al: Polydatin alleviates
high-fat diet induced atherosclerosis in apolipoprotein E-deficient
mice by autophagic restoration. Phytomedicine.
81(153301)2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lanzilli G, Cottarelli A, Nicotera G,
Guida S, Ravagnan G and Fuggetta MP: Anti-inflammatory effect of
resveratrol and polydatin by in vitro IL-17 modulation.
Inflammation. 35:240–248. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lv R, Du L, Zhang L and Zhang Z: Polydatin
attenuates spinal cord injury in rats by inhibiting oxidative
stress and microglia apoptosis via Nrf2/HO-1 pathway. Life Sci.
217:119–127. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Li XH, Gong X, Zhang L, Jiang R, Li HZ, Wu
MJ and Wan JY: Protective effects of polydatin on septic lung
injury in mice via upregulation of HO-1. Mediators Inflamm.
2013(354087)2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Sasaki I, Hoshino K, Sugiyama T, Yamazaki
C, Yano T, Iizuka A, Hemmi H, Tanaka T, Saito M, Sugiyama M, et al:
Spi-B is critical for plasmacytoid dendritic cell function and
development. Blood. 120:4733–4743. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Laramee AS, Raczkowski H, Shao P, Batista
C, Shukla D, Xu L, Haeryfar SMM, Tesfagiorgis Y, Kerfoot S and
DeKoter R: Opposing roles for the related ETS-Family transcription
factors Spi-B and Spi-C in Regulating B cell differentiation and
function. Front Immunol. 11(841)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Garrett-Sinha LA, Hou P, Wang D, Grabiner
B, Araujo E, Rao S, Yun TJ, Clark EA, Simon MC and Clark MR: Spi-1
and Spi-B control the expression of the Grap2 gene in B cells.
Gene. 353:134–146. 2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Zhou J, Lin J, Zhang H, Zhu F and Xie R:
LncRNA HAND2-AS1 sponging miR-1275 suppresses colorectal cancer
progression by upregulating KLF14. Biochem Biophys Res Commun.
503:1848–1853. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Liu Y, Wu H, Nie YC, Chen JL, Su WW and Li
PB: Naringin attenuates acute lung injury in LPS-treated mice by
inhibiting NF-κB pathway. Int Immunopharmacol. 11:1606–1612.
2011.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Takagi Y, Shimada K, Shimada S, Sakamoto
A, Naoe T, Nakamura S, Hayakawa F, Tomita A and Kiyoi H: SPIB is a
novel prognostic factor in diffuse large B-cell lymphoma that
mediates apoptosis via the PI3K-AKT pathway. Cancer Sci.
107:1270–1280. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li SK, Abbas AK, Solomon LA, Groux GM and
DeKoter RP: Nfkb1 activation by the E26 transformation-specific
transcription factors PU.1 and Spi-B promotes Toll-like
receptor-mediated splenic B cell proliferation. Mol Cell Biol.
35:1619–1632. 2015.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jin YL, Xin LM, Zhou CC and Ren Y:
Polydatin exerts anti-tumor effects against renal cell carcinoma
cells via induction of caspase-dependent apoptosis and inhibition
of the PI3K/Akt pathway. Onco Targets Ther. 11:8185–8195.
2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Lv X, Zhang XY, Zhang Q, Nie YJ, Luo GH,
Fan X, Yang S, Zhao QH and Li JQ: lncRNA NEAT1 aggravates
sepsis-induced lung injury by regulating the miR-27a/PTEN axis. Lab
Invest. 101:1371–1381. 2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
National Institute of Health: Guide for
the Care and Use of Laboratory Animals. The National Academies
Press, Washington, DC, 1996.
|
|
23
|
Geng P, Ma T, Xing J, Jiang L, Sun H, Zhu
B, Zhang H, Xiao H, Wang J and Zhang J: Dexamethasone ameliorates
H2S-induced acute lung injury by increasing claudin-5 expression
via the PI3K pathway. Hum Exp Toxicol. 37:626–635. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chen G, Yang Z, Wen D, Guo J, Xiong Q, Li
P, Zhao L, Wang J, Wu C and Dong L: Polydatin has anti-inflammatory
and antioxidant effects in LPS-induced macrophages and improves
DSS-induced mice colitis. Immun Inflamm Dis. 9:959–970.
2021.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Oliviero F, Galozzi P, Scanu A, Galuppini
F, Lazzarin V, Brocco S, Ravagnan G, Sfriso P, Ramonda R, Spinella
P, et al: Polydatin prevents calcium pyrophosphate crystal-induced
arthritis in mice. Nutrients. 13(929)2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shen P, Han L, Chen G, Cheng Z and Liu Q:
Emodin attenuates acetaminophen-induced hepatotoxicity via the
cGAS-STING Pathway. Inflammation. 45:74–87. 2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tian J, Jiao X, Wang X, Geng J, Wang R,
Liu N, Gao X, Griffin N and Shan F: Novel effect of methionine
enkephalin against influenza A virus infection through inhibiting
TLR7-MyD88-TRAF6-NF-κB p65 signaling pathway. Int Immunopharmacol.
55:38–48. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Huang Q, Liu J, Wu S, Zhang X, Xiao Z, Liu
Z and Du W: Spi-B Promotes the Recruitment of Tumor-Associated
Macrophages via Enhancing CCL4 expression in lung cancer. Front
Oncol. 11(659131)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Miyazaki R, Saiga H, Kato T, Bakoshi T,
Senba R, Shintani A, Suzuki M, Takao K, Sasaki I, Iizuka A, et al:
The mechanism of action of Spi-B in the transcriptional activation
of the interferon-α4 gene. Biochem Biophys Res Commun. 525:477–482.
2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Su GH, Chen HM, Muthusamy N, Garrett-Sinha
LA, Baunoch D, Tenen DG and Simon MC: Defective B cell
receptor-mediated responses in mice lacking the Ets protein, Spi-B.
EMBO J. 16:7118–7129. 1997.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gotts JE, Chun L, Abbott J, Fang X,
Takasaka N, Nishimura SL, Springer ML, Schick SF, Calfee CS and
Matthay MA: Cigarette smoke exposure worsens acute lung injury in
antibiotic-treated bacterial pneumonia in mice. Am J Physiol Lung
Cell Mol Physiol. 315:L25–L40. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jiang Q, Yi M, Guo Q, Wang C, Wang H, Meng
S, Liu C, Fu Y, Ji H and Chen T: Protective effects of polydatin on
lipopolysaccharide-induced acute lung injury through
TLR4-MyD88-NF-κB pathway. Int Immunopharmacol. 29:370–376.
2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Gao L, Tang X, He Q, Sun G, Wang C and Qu
H: Exosome-transmitted circCOG2 promotes colorectal cancer
progression via miR-1305/TGF-β2/SMAD3 pathway. Cell Death Discov.
7(281)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ogawa N, Nakajima S, Tamada K, Yokoue N,
Tachibana H, Okazawa M, Oyama T, Abe H, Yamazaki H, Yoshimori A, et
al: Trimebutine suppresses Toll-like receptor 2/4/7/8/9 signaling
pathways in macrophages. Arch Biochem Biophys.
711(109029)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Cao J, Guo B, Li S, Zhang X, Zhang X,
Zhang G, Sun Y, Wang Y, Song X and Zhang Z: Neuroprotection against
1-Methyl-4-phenylpyridinium-induced cytotoxicity by naturally
occurring polydatin through activation of transcription factor
MEF2D. Neuroreport. 32:1065–1072. 2021.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hou XJ, Ye LX, Ai XY, Hu CG, Cheng ZP and
Zhang JZ: Functional analysis of a PISTILLATA-like gene CcMADS20
involved in floral organs specification in citrus. Plant Sci.
319(111263)2022.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhu M, Bin J, Ding H, Pan D, Tian Q, Yang
X, Wang L and Yue Y: Insights into the trihelix transcription
factor responses to salt and other stresses in Osmanthus fragrans.
BMC Genomics. 23(334)2022.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wei H, Xue Q, Sun L and Lv J: BRD4
inhibition protects against myocardial ischemia/reperfusion injury
by suppressing inflammation and oxidative stress through PI3K/AKT
signaling pathway. J Cardiovasc Pharmacol. 78:839–846.
2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhou B, Weng G, Huang Z, Liu T and Dai F:
Arctiin prevents LPS-Induced acute lung injury via inhibition of
PI3K/AKT signaling pathway in mice. Inflammation. 41:2129–2135.
2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ali FEM, Ahmed SF, Eltrawy AH, Yousef RS,
Ali HS, Mahmoud AR and Abd-Elhamid TH: Pretreatment with Coenzyme
Q10 combined with aescin protects against sepsis-induced acute lung
injury. Cells Tissues Organs. 210:195–217. 2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Huang B, Liu J, Meng T, Li Y, He D, Ran X,
Chen G, Guo W, Kan X, Fu S, et al: Polydatin prevents
lipopolysaccharide (LPS)-induced Parkinson's disease via regulation
of the AKT/GSK3β-Nrf2/NF-κB signaling axis. Front Immunol.
9(2527)2018.PubMed/NCBI View Article : Google Scholar
|