Introduction
It is well known that microorganisms release primary
and secondary metabolites as well as peptides and proteins into
their environment (in either the host organism or the nutrition
medium). The function of releasing these products is mainly
unknown. Recently, we demonstrated that Klebsiella
pneumoniae (K. pneumoniae) and Streptococcus
pneumoniae release proteins with a molecular weight of 30–50
kDa into the nutritional medium during the exponential growth phase
(1). The use of this protein
fraction for vaccination conferred complete protection of mice from
K. pneumoniae infection. It was proven that this protection
was not due to protease, one of the key invasion factors of K.
pneumoniae, nor by antigens of the bacterial cell wall.
Notably, optimal protection was observed with proteins obtained in
the exponential growth phase of the bacteria, i.e., proteins
involved in the transition of bacteria from a resting to a
proliferative state. This result may be of significant practical
value for the onset of the infection process. However, the absence
of the amino acid sequences of these proteins did not allow
identification of these proteins.
Recently, the genomes of 9 strains of
Staphylococcus aureus (S. aureus) have been sequenced
and annotated [http://www.ncbi.nlm.nih.gov/genomes/lproks.cgi;
http://www.tigr.org] (14). A more detailed database of S.
aureus is now available with both genomic and proteomic
information (2). During the past
decade, several investigations have addressed the issue of secreted
proteins from S. aureus (3,4). It
can be hypothesized that the cellular and molecular mechanisms
responsible for transition from a resting to a proliferative state
are common among microorganisms. Nutritional substances taken up by
bacteria are proposed primer signals to start growth. Due to the
energy required to boost metabolism, subsets of proteins rather
than the total spectrum of enzymes are induced. Therefore, the aim
of the present investigation was to determine the protein spectrum
synthesized and released by S. aureus at the early stage of
growth.
Materials and methods
Animal experimentation
The experiments were carried out using 2- to
3-month-old Balb/c male mice. The animals were obtained from
Stolbovaya Company (Moscow, Russia). The mice received standard
laboratory feed and tap water ad libitum. All experiments
were carried out in accordance with the legal regulations for
animal experimentation in Russia and with official permission of
the Institute of Experimental Diagnosis and Therapy of Tumors of
the N.N. Blokhin Russian Cancer Research Center and the Mechnikov
Research Institute for Vaccine and Sera, Russian Academy of Medical
Science, Moscow, Russia (registration code 076). Mice were
sacrificed prior to death during the experiments.
Bacterial strains and growth
conditions
S. aureus strain 6 was obtained from the
collection of the L.A. Tarasevich State Scientific Research
Institute for Standardization and Quality Control of Biologicals,
Moscow, Russia. It was previously shown that this strain is the
most virulent for conventional mice among all S. aureus
strains (5). Liquid nutritional
medium based on casein hydrolyzed from pancreas supplemented with
yeast extract and glucose was used for cultivation (6). Five to seven colonies isolated from
agar culture were transferred into a flask with 200 ml nutritional
medium. After 12–14 h of incubation, the growing culture was
transferred into a flask with 2 l of medium at a 1:10 proportion of
culture to medium, and incubated at 37°C at 90 rmp/min on a
Multi-Shaker Multi PSU-20 (BioSan Ltd., Riga, Latvia). The
bacterial growth was monitored using a spectrophotometer (Genesys
10 UV; Thermo Spectronic, Rochester, NY, USA) by measuring the
optical density at 565 nm. The number of colonies was determined,
calculated 24 h after incubation on agar plates and inoculated with
serial dilutions of the bacterial culture. The efficacy of the
cultivation process was evaluated every hour by the density of the
biomass calculated as the number of colony forming units
(CFUs).
Extraction of S. aureus extracellular
proteins
Bacterial cells were separated from the culture
media by sterilization filtration using 0.22-μM pore filters
(Millipore, MA, USA). For subsequent protein extraction, 80%
ammonium sulphate precipitation was used. The precipitate was
centrifuged (1,7). The pellet was dissolved in 0.01 M
Tris-buffer (pH 7.4) with 0.01% sodium aside and transferred into
50 mM ammonium carbonate using a PD-10 column (GE Health Care,
USA).
One-dimensional SDS-PAGE
Proteins were separated by electrophoresis in 12.5%
polyacrylamide gel (8). The
samples were dissolved in buffer consisting of 63 mM Tris/HCL, pH
6.8, 10% (v/v) glycerol, 2% (m/v sodium dodecyl sulphate (SDS) and
30 μM bromophenol blue. Samples were applied in quantities of 20 μg
protein per lane.
Liquid chromatography coupled with mass
spectrometry (LC-MS) analysis
After SDS-PAGE electrophoresis (12.5% gel), gels
were stained with Coomasse Brilliant Blue R-250, and protein strips
were dissected. Pieces, 1x1 mm, were cut out from the protein
strip. These gel pieces were washed for 5 min in 70 μl 50% 200 mM
NH4HCO3 – 50% acetonitryl (v/v) and
transferred to 70 μl acetonitryl for 20 min. Acetonitryl was
removed, and the gels were dried using a SpeedVac for 20 min. Dried
gel pieces were transferred to a solution containing 3 μl trypsin
(15 ng/μl) in 50 mM NH4HCO3, incubated on ice
for 30 min, and then kept overnight at 37°C.
LC-MS analysis was performed on a 1200 Series
HPLC-Chip (Agilent Technology, Inc., Santa Clara, CA, USA) in
combination with an Agilent 6520 Accurate-Mass Q-TOF LC/MS system
(Agilent Technology, Inc.).
In the present investigation, an ID chip with a
150×0.075 mm analytic column and 40 nl concentrating column were
used. The conditions of analysis were as follows. A 5-μl peptide
solution sample was obtained from the gel after trypsin digestion.
The flow speed was 300 nl/min for the pumps of the analytic and 3
μl/min for the pump of the concentrating columns. The composition
of liquid phase A was 5% acetonitryl and 0.1% formic acid. Liquid
phase B consisted of 90% acetonitryl and 0.1% formic acid.
Gradients: 0% B to 50% during 50 min, then 80% B during 60 min up
to 65 min, then 0% B at 65.1 min; stopping the separation. The
samples were applied on the chip in 100% of buffer A.
Bioinformatic methods
Data analysis was performed by comparing MS/MS
spectra against the European Bioinformatics Institute non-redundant
proteome set of Swiss-Prot entries through the Spectrum Mill MS
Proteomics Workbench (Agilent), with no static or dynamic
modifications. The other settings of the Spectrum Mill MS
Proteomics Workbench were: species subset database, human;
instrument, Agilent Q-TOF; precursor mass tolerance, 5 ppm; digest
specificity, no enzyme; and search mode, identity. Results were
autovalidated after each search using the default parameters for
autovalidation.
Protein database searches were performed with
Spectrum Mill MS Proteomic Workbench Rev A.03.03.084. SR4 (licensed
to Kurchatov Institute, Moscow, Russia) using the MSDB database of
protein sequences (http://www.proteomics.leeds.ac.uk/bioninf/msdb.html;
restoration August 31, 2006; Proteomics Department at the
Hammersmith Campus of Imperial College, London, UK), with no static
or dynamic modifications. The other settings of the Spectrum Mill
MS Proteomics Workbench were: species subset database, S.
aureus; instrument, Agilent Q-TOF; precursor mass tolerance, 5
ppm; digest specificity, trypsin; maximum number of missed
cleavages, 1; and search mode, identity. Results were autovalidated
after each search using the default parameters for
autovalidation.
Protein measurement
The amount of secreted proteins in the immune doses
was measured by the Bradford method (9).
Statistics
Analysis of variance with the Bonferroni post
hoc test was used for statistical analyses. Analyses were
performed with Prism (version 5.0b) software. Values were
considered significant at a P-value <0.05.
Results
Bacterial growth and SDS-PAGE
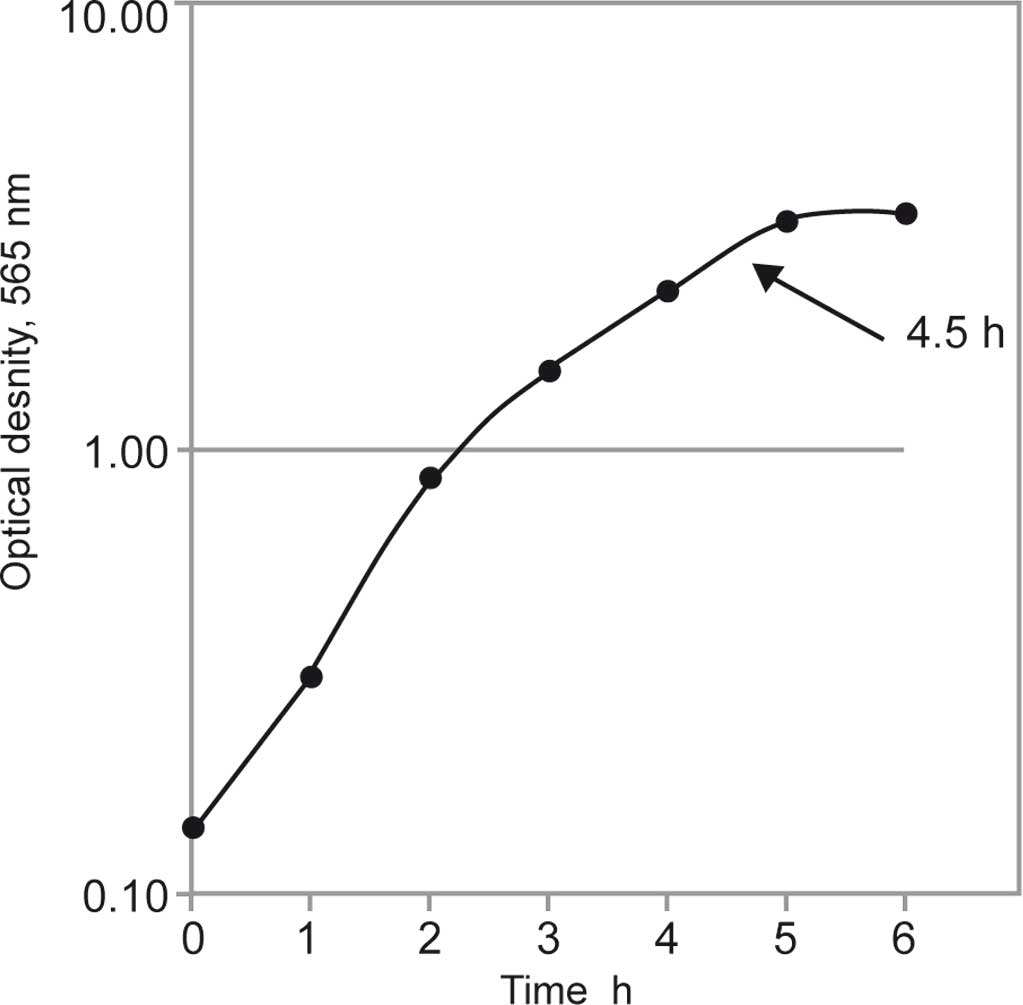
We first analyzed the growth kinetics under
cultivation conditions described in Materials and methods. As shown
in Fig. 1, the exponential growth
phase of S. aureus strain 6 ended after 5 h. At the end of
the exponential growth phase (after 4.5 h of cultivation), the
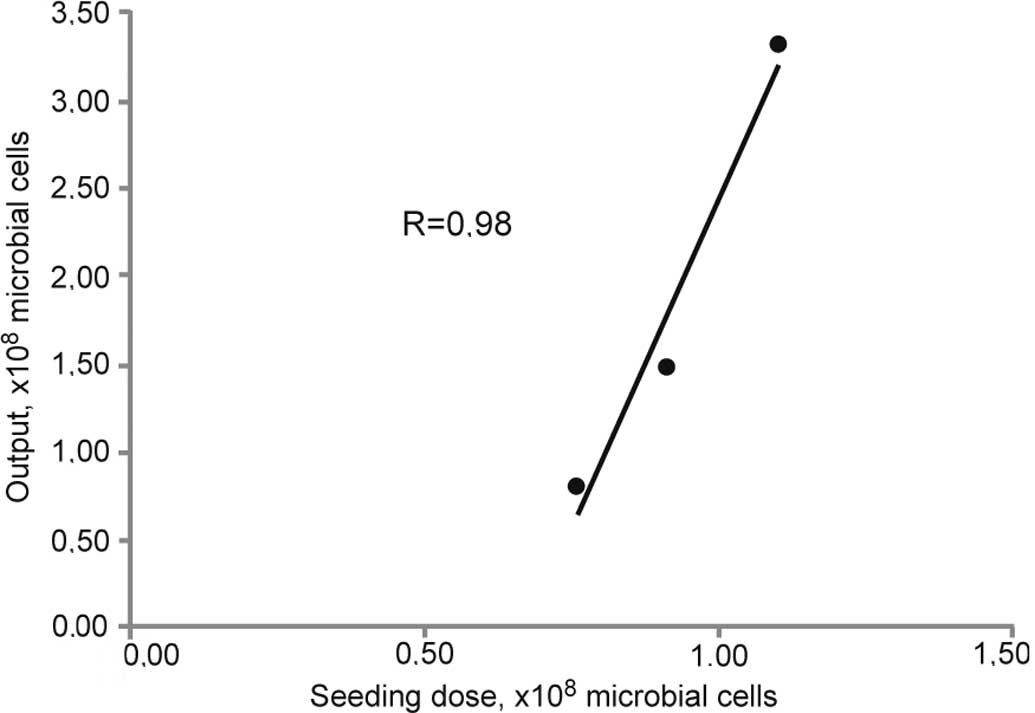
number of CFUs was counted. A linear correlation between the number
of seeded cells and the efficacy of the cultivation process was
observed as measured by biomass density (Fig. 2). All further experiments were
performed with a seeding number of 1×108 CFUs and a
4.5-h cultivation time.
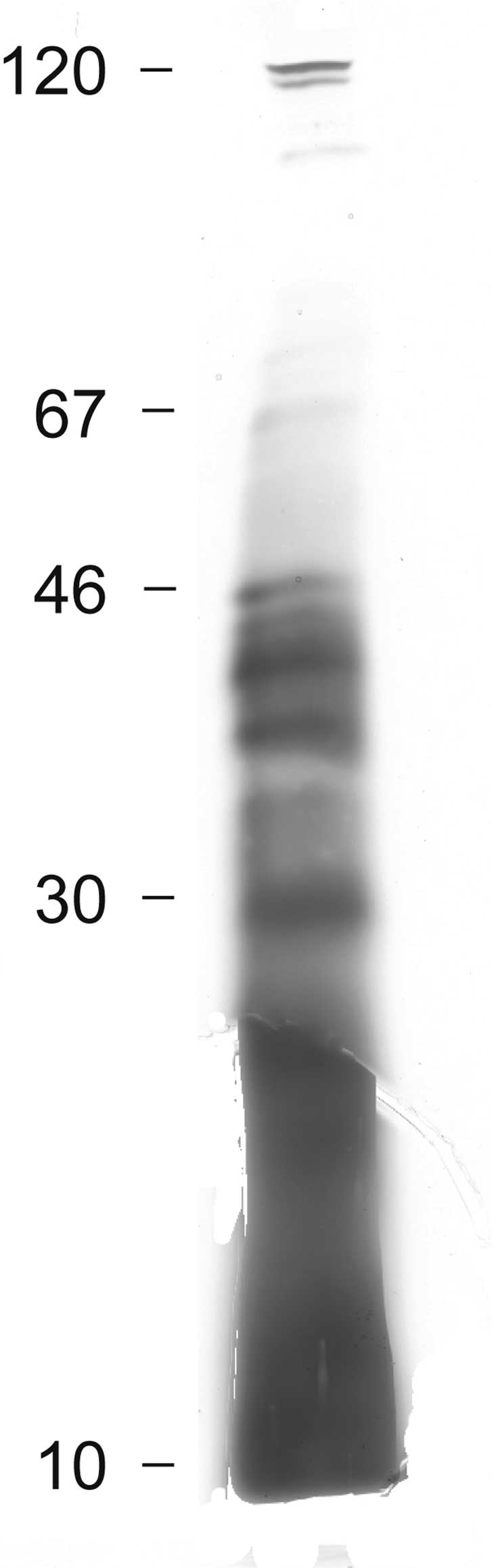
The bacterial cells were separated from the culture
medium which was further processed as previously described
(1,7). A filtrate of the culture medium was
subjected to SDS-PAGE (Fig. 3).
Electrophoretic separation showed two faint bands in the region of
100 kDa and four intense and several weak bands between 30 and 50
kDa. Below 30 kDa, no distinct bands were observed, although silver
staining revealed the presence of undefined biological material.
This may be indicative of polysaccharides and proteoglycans as
previously identified in K. pneumoniae (1).
LC-MS analysis
The protein bands between 30 and 50 kDa were cut out
and digested by trypsin, and subjected to LC-MS and MSDB analyses.
A total of 208 peptides were identified. One hundred and fourteen
of them were unique, and the average number of the unique peptides
for one protein was 10.4 (median seven peptides/protein). As a
result, 11 proteins were determined. Using PSOTb software (10), the isoelectric points (pI) of the
proteins were calculated. A cytosolic localization was determined
for 3 proteins (CP-N3, 5 and 11). One protein appeared to be
localized in the cell membrane (CM-N4-lipoprotein) and one protein
was extracellular (EC-N9-superoxide dismutase). The localization of
the other 6 proteins could not be determined using PSORTb (Table I). These 6 proteins were further
analyzed using the Uniprot Knowledgebase and the database of the
European Institute for Bioinformatics (11). This analysis revealed that 4 of
these 6 proteins were cytoplasmatically located and played a role
in cellular metabolism: carbohydrate metabolism (N8 – protein
SAS0528, hexulose-6-phosphate synthase), glycolysis (N1 – fructose
bisphosphate aldolase class 1 and N10 – transaldolase) and glycine
cleavage (N2). The supposed glycerophosphoryl-diester
phosphodiesterase (N7) and protein SA0295 (N6) contained N-terminal
signal peptides, indicating that they were extracellular localized
or non-covalently linked to the bacterial cell wall. SA0295 (N6)
was highly similar to the amino acid sequence of S. aureus
acid phosphatase. Hence, by using two approaches, 7 out of 11
proteins were identified as having a cytoplasmic origin and 3 out
of 11 were secreted (3,4).
 | Table IExtracellular proteins identified by
MALDI-TOF mass-spectrometry in the exponential growth phase of
S. aureus strain 6. |
Table I
Extracellular proteins identified by
MALDI-TOF mass-spectrometry in the exponential growth phase of
S. aureus strain 6.
| N | Accession no. | Gene symbol | Ordered locus
name | Protein
identification | MW (Da) | pI | Protein score | Coincident
peptides | The most similar
strains | Localization
according to PSORTb
|
|---|
| CP | CM | CW | EC | Total |
|---|
| 1 | Q2YWF3 | fba | SAB2479 | Fructose bisphosphate
aldolase class 1 | 32907 | 4.69 | 788 | 25 | RF122, MRSA252 | 2.50 | 2.50 | 2.50 | 2.50 | U |
| 2 | B89855 | gcvH | SA0760 | Glycine cleavage
system H protein | 14072 | 3.70 | 140 | 2 | N315, RF122, NCTC
8325, MW2 | 2.50 | 2.50 | 2.50 | 2.50 | U |
| 3 | A89896 | fabD | SA1073 | Malonyl CoA-acyl
carrier protein transacylase | 33628 | 4.59 | 97 | 3 | N315, RF122,
MRSA252, NCTC 8325, MRSA476 | 9.67 | 0.01 | 0.15 | 0.17 | CP |
| 4 | H89832 | SA0587 | SA0587 | Lipoprotein,
Streptococcal adhesin PsaA homologue | 35049 | 9.16 | 120 | 2 | N315 | 0.00 | 9.68 | 0.17 | 0.16 | CMa |
| 5 | F89850 | pgk | SA0728 | Phosphoglycerate
kinase | 42575 | 4.96 | 373 | 14 | N315, RF122 | 10.00 | 0.00 | 0.00 | 0.00 | CP |
| 6 | D89795 | SA0295 | SA0295 | Similar to outer
membrane protein precursor | 33331 | 10.08 | 353 | 20 | N315, MRSA252 | 0.00 | 3.33 | 3.33 | 3.33 | Ua |
| 7 | H89862 | glpQ | SA0820 | Putative
glycerophosphoryl diester phosphodiesterase | 35289 | 9.14 | 87 | 2 | N315, MRSA252 | 0.00 | 0.15 | 4.55 | 5.15 | Ua |
| 8 | D89825 | SAS0528 | SAS0528 | Putative
hexulose-6-phosphate synthase | 22422 | 4.32 | 244 | 3 | N315, RF122, MW2,
USA300 | 2.50 | 2.50 | 2.50 | 2.50 | U |
| 9 | H89935 | sodA | SA1382 | Superoxide
dismutase | 22697 | 4.86 | 387 | 17 | N315 | 0.01 | 0.09 | 0.18 | 9.72 | EC |
| 10 | F89963 | SA1599 | SA1599 | Similar to
transaldolase | 25742 | 4.51 | 260 | 7 | N315, RF122,
JH1 | 2.50 | 2.50 | 2.50 | 2.50 | U |
| 11 | G89850 | tpiA | SA0729 | Triosephosphate
isomerase | 27245 | 4.51 | 539 | 19 | N315, RF122, NCTC
8325 | 9.67 | 0.01 | 0.15 | 0.17 | CP |
Protective activity of the secreted
proteins
The protein bands between 30 and 50 kDa were used
for immunization of mice. S. aureus strain 6
(4×108 cells) was then injected into the animals. The
protective activity of this vaccination has been estimated by the
survival of animals after infection. Two different doses of
secreted proteins (2 and 30 μg) were injected into mice, both of
which revealed protective activity. As shown in Table II, all of the animals of the
control group died. Vaccination of mice with the secreted proteins
led to survival of half of the animals (P<0.05). In a second
experiment, the LD50 values were determined in
vaccinated and non-vaccinated mice by injection of varying numbers
of injected S. aureus strain 6 cells. Inoculates with six
different cell densities of S. aureus strain 6 ranging from
1×107 to 6×108 cells were injected into the
peritoneal cavity of mice. In the immunized mice, the
LD50 was 4.3×108 microbial cells and in
control mice 1.8×108 cells.
| Table IIProtective activity of the secreted
proteins of S. aureus strain 6. |
Table II
Protective activity of the secreted
proteins of S. aureus strain 6.
| Immunization dose
(μg/mice) | No. of mice per
group | No. of surviving
mice | Statistical
significance |
|---|
| Control | - | 10 | 0 | |
| 1st vaccinated
group | 30 | 10 | 5 | P<0.05 |
| 2nd vaccinated
group | 2 | 10 | 6 | P<0.05 |
Discussion
In the present investigation, we found 11 proteins
in the culture medium of exponentially growing S. aureus
strain 6. Additional compounds identified by gel electrophoresis
presumably were polysaccharides and proteoglycans as recently
observed in the culture medium of K. pneumonie (1). A significant finding was that we did
not identify proteins which are known to be secreted by bacteria,
but also membrane-bound and cytoplasmic proteins. To minimize the
contamination of the nutritional medium with dying cells, which
non-specifically release their intracellular constituents, we only
used cells in the exponential growth phase. Thereby, a massive
destruction of cells releasing intracellular proteins was
prevented. For this reason, we conclude that these proteins were
secreted from the bacteria into the culture medium by specific
transport processes. Another reason for the specificity of this
observation is that only this small profile of 11 proteins was
present in the medium and not the broad spectrum of hundreds of
intracellular proteins as could be expected from cell lysis.
Notably, similar profiles of secreted proteins as determined in the
present investigation for S. aureus, were also described for
K. pneumoniae and Streptococcus pneumoniae (1).
If we assume that these proteins were specifically
released by the exponentially growing cells via yet unknown
secretory pathways, the question arises as to the function of these
proteins. The determination of the isoelectric points revealed that
two proteins were in the alkaline diapason (pI for protein N4 is
9.16 and for protein N6 is 10.08), whereas all others had their
isoelectric point at acidic pH values. N6 was assigned as a protein
transporter by database mining, while the function of N4 is
unknown. According to the general physicochemical properties of
proteins, it is highly probable that they form complexes with
acidic proteins. These complexes would then have a neutral charge.
A neutral charge of protein complexes is one of the conditions
sufficient for molecular transport through cellular membranes.
Hence, it can be hypothesized that complex formation of alkaline
and acidic proteins may explain the secretion of the 11 proteins
identified by us.
It has been shown that decreased isoelectric points
are associated with weaker immunogenicity of proteins (12). Hence, it can be expected that the 9
acidic proteins in our study may exert weak immunogenicity, whereas
the two alkaline proteins provoke strong immune reactions. The
explanation for a causative association between molecular charge
and immunogenicity is that cell membranes normally have a negative
charge. Therefore, acidic molecules (with negative charge) are
rejected by membranes of immune competent cells and alkaline
molecules (with a positive charge) are attracted.
Determination of the proposal functions
of the identified proteins
Superoxide dismutase (N9) [sodA (SA1382)] is
well known as a component of the antioxidant stress response. This
enzyme is predominantly expressed at the end of the exponential
growth phase of S. aureus (13) and may take part in carbohydrate
metabolism-related redox reactions, i.e., in energy transport. Two
other extracellular proteins – glycerophosphoryl diester
phosphodiesterase (N7) [glpQ (SA0820)] (14) and protein N6 (SA0295), with
similarity to acid phosphatase S. aureus (15), probably hydrolyze biological
macromolecules assimilated by bacterial cells, e.g., glycerine, fat
acids and phosphatases.
As predicted by the PSORTb software, protein N4
[lipoprotein (SA0587)] is localized in the cell membrane, as this
protein contains a transmembrane internal spiral as a signal
peptide. There is a high similarity of this protein with
ATP-binding cassette substrate-linking lipoprotein SA0587, which
transports metal ions into cells. Furthermore, it was reported that
this protein, which has a positive charge, may transport negatively
charged enzymes (16). It is
conceivable, therefore, that basic residues in this protein may be
involved in electrostatic interactions with negatively charged
substrates
Of the 7 cytoplasmic proteins, 5 are involved in
carbohydrate metabolism: N1 – fructose bisphosphate aldolase class
1 [fba (SAB2479)], N11 – triosephosphate isomerase
[tpiA (SA0729)] and N5 – phosphoglycerate kinase [pgk
(SA0728)] – are glycolys enzymes. N10 – transaldolase (SA1599)
links glycolys with the pentose phosphate shunt. N8 – proposed
hexulose-6-phosphate synthase [sgaH (SAS0528)] catalyses the
reversible reaction of assimilated formaldehyde by
ribuloso-5-phosphate to arabinoso-6-phosphate. The reversed
reaction is also a part of the de novo pyrimidine
biosynthesis. The other two enzymes, N3 – transacylase malonyl
CoA-acyl carrier protein [fabD (SA1073)] and N2 – protein of
the H system cleavage glycine [gcvH (SA0760)] are involved
in the biosynthesis of fatty acids and decarboxylation of glycine,
respectively.
The exponential growth phase is very important for
the development of bacteria, since resting bacterial cells have
only limited energy reserves. Therefore, the start of growth is
causatively linked with the activation of energy metabolism.
The result of the present investigation that S.
aureus releases only a limited number rather than thousands of
proteins was confirmed by comparable investigations of other
authors. Ziebandt et al identified 43 extracellular S.
aureus proteins, including those controlled by the additional
regulator-gene (agr) and/or alternative factor σB
(sigB) (17). Nakano et
al identified 29 extracellular proteins produced by
methicillin-resistant strains of S. aureus (14) using two-dimensional electrophoresis
linked with the N-terminal sequence of amino acids. At the time
point of the transition from the resting to the proliferative
state, bacterial cells do not have enough energy and nutritional
resources to synthesize the entire spectrum of proteins. Therefore,
only a limited number of proteins with crucial functional
importance is synthesized by the microorganisms. We speculate that
the host immune response may inhibit bacterial growth by attracting
extracellular bacterial proteins leading to energy depletion rather
than acting on the bacterial cell itself.
Our hypothesis that enzymes of the carbohydrate
metabolism act as signals for the development of infections is
supported by the fact that patients with diabetes mellitus more
frequently suffer from severe bacterial complications than
non-diabetic subjects. There are several hints from the literature
supporting this point of view. Diabetic patients are prone to
develop all types of infections. Pneumococcal infections are a
common cause of morbidity and mortality among individuals affected
with diabetes (18). Furthermore,
pyomyositis is a relatively infrequent, subacute primary bacterial
muscle infection. Due to its non-specific clinical symptoms, it is
unlikely to be diagnosed early particularly in diabetic patients.
This delay in diagnosis may be fatal (19).
In a large nationwide survey of diabetic patients
undergoing a variety of non-cardiac surgical procedures, glucose
control in the first 24 h after surgery was poor, and mean serum
glucose concentrations of ≥150 mg/dl during this time period were
associated with increased rates of postoperative infectious
complications (20).
These data are in accord with results showing that
various growth factors, which regulate cell growth also influence
glucose metabolism (21). In this
case, increased concentrations of carbohydrates may be a signal for
microorganisms ‘to wake up’ and grow. Such a growth signal may also
be too strong to kill microorganisms by chemotherapy, as they have
enough energy to survive a challenge by drugs.
In conclusion, the identification of carbohydrate
metabolism-related enzymes and transporter proteins in the
extracellular medium of S. aureus may have implications for
the development of new strategies for prevention and therapy of
S. aureus infections. Preliminary data concerning the
protective activity of the immunization of animals by these
secreted proteins confirms our assumption of the importance of
these proteins for the viability of microorganisms.
References
|
1
|
Trishin AV, Donenko FV, Kurbatova EA,
Voyushin KE, Kiselevsky MV, Egorova NB, Gruber IM, Semenova IB and
Semenov BF: Protective activity of secreted proteins of
Streptococcus pneumoniae and Klebsiella pneumoniae.
Zh Microbiol. 4:46–50. 2008.PubMed/NCBI
|
|
2
|
Jones RC, Deck J, Edmondson RD and Hart
ME: Relative quantitative comparisons of the extracellular protein
profiles of Staphylococcus aureus UAMS-1 and its sarA, agr,
and sarA agr regulatory mutants using 1D-PAGE and nanocapillary LC
coupled with tandem MS. J Bacteriol. 190:5265–5278. 2008.PubMed/NCBI
|
|
3
|
Burlak C, Hammer CH, Robinson MA, Whitney
AR, McGavin MJ, Kreiswirth BN and Deleo FR: Global analysis of
community-associated methicillin-resistant Staphylococcus
aureus exoproteins reveals molecules produced in vitro and
during infection. Cell Microbiol. 9:1172–1190. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sibbald MJ, Ziebandt AK, Engelmann S,
Hecker M, de Jong A, Harmsen HJ, Raangs GC, Stokroos I, Arends JP,
Dubois JY and van Dijl JM: Mapping the pathways to staphylococcal
pathogenesis by comparative secretomics. Microbiol Mol Biol Rev.
70:755–788. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Egorova NB and Korzaya LI: The effect of
repeated administration of staphylococcal immune preparations on
the development of local staphylococcal infection in mice. J Hyg
Epidemiol Microbiol Immunol. 27:203–210. 1983.PubMed/NCBI
|
|
6
|
Kuzmenko OM, Zlygostev SA, Mikhaylova NA,
Gruber IM, Akhmatova NK, Kurbatova EA and Cherkasova LS:
Characteristics of antigenic complexes of Staphylococcus
aureus vaccine strains obtained in different cultivation
conditions. Zh Microbiol. 2:51–54. 2010.PubMed/NCBI
|
|
7
|
Donenko FV, Sitdikova SM, Syrtsev AV,
Gradyushko AT, Kiselevsky MV, Serebryakova MV and Efferth T:
Hemoglobin-associated proteins isolated from blood serum of Ehrlich
carcinoma-bearing mice. Int J Oncol. 32:885–893. 2008.PubMed/NCBI
|
|
8
|
Laemmli UK: Cleavage of structural
proteins during the assembly of head of bacteriophage T4. Nature.
227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gardy JL, Laird MR, Chen F, Rey S, Walsh
CJ, Ester M and Brinkman FSL: PSORTb v. 2.0: expanded prediction of
bacterial protein subcellular localization and insights gained from
comparative proteome analysis. Bioinformatics. 21:617–623. 2005.
View Article : Google Scholar
|
|
11
|
The UniProt Consortium: The universal
protein resource (UniProt). Nucleic Acids Res. 35:D193–D197.
2007.
|
|
12
|
Hattori M, Miyakawa S, Ohama Y, Kawamura
H, Yoshida T, To-o K, Kuriki T and Takahashi K: Reduced
immunogenicity of beta-lactoglobulin by conjugation with acidic
oligosaccharides. J Agric Food Chem. 52:4546–4553. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Clements MO, Watson SP and Foster SJ:
Characterisation of the major superoxide dismutase of
Staphylococcus aureus and its role in starvation survival,
stress resistance, and pathogenicity. J Bacteriol. 181:3898–3903.
1999.PubMed/NCBI
|
|
14
|
Nakano M, Kawano Y, Kawagish M, Hasegawa
T, Iinuma Y and Oht M: Two-dimensional analysis of exoproteins of
methicillin-resistant Staphylococcus aureus (MRSA) for
possible epidemiological applications. Microbiol Immunol. 46:11–22.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du Plessis EM, Theron J, Joubert L, Lotter
T and Watson TG: Characterization of a phosphatase secreted by
Staphylococcus aureus strain 154, a new member of the
bacterial class C family of nonspecific acid phosphatases. Syst
Appl Microbiol. 25:21–30. 2002.PubMed/NCBI
|
|
16
|
Alexeyev MF and Winkler HH: Complete
replacement of basic amino acid residues with cysteines in
Rickettsia prowazekii ATP/ADP translocase. Biochim Biophys
Acta. 1565:1362002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ziebandt AK, Becher D, Ohlsen K, Hacker J,
Hecker M and Engelmann S: The influence of agr and sigmaB in growth
phase dependent regulation of virulence factors in
Staphylococcus aureus. Proteomics. 4:3034–3047. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mohan V, Unnikrishnan R, Thomas N,
Bhansali A, Wangnoo SK and Thomas K: Pneumococcal infections and
immunization in diabetic patients. J Postgrad Med. 57:78–81. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abdullah ZS, Khan MU, Kodali SK and Javaid
A: Pyomyositis mimicking osteomyelitis detected by SPET/CT. Hell J
Nucl Med. 13:277–279. 2010.PubMed/NCBI
|
|
20
|
King JT, Goulet JL, Perkal MF and
Rosenthal RA: Glycemic control and infections in patients with
diabetes undergoing noncardiac surgery. Ann Surg. 253:158–165.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Heiden MGV, Plas DR, Rathmell JC, Harris
MH and Thompson CB: Growth factors can influence cell growth and
survival though effects on glucose metabolism. Mol Cell Biol.
21:5899–5912. 2001. View Article : Google Scholar : PubMed/NCBI
|