Introduction
Influenza has been responsible for millions of
deaths worldwide. Influenza virus type A is antigenically highly
variable and is responsible for most cases of epidemic influenza.
In 1997, the first instance of human respiratory disease and death
associated with the H5N1 avian influenza virus was documented. In
2009, the outbreak of human infection with a novel influenza A
(H1N1) virus, which spread through sustained human-to-human
transmission, caused a world pandemic.
When a person is infected with uncomplicated
influenza, the symptoms usually last for a short period and the
patient recovers rapidly, but serious complications may occur in
elderly individuals and those with underlying chronic diseases.
Pneumonia is a common complication of influenza virus infection.
Primary viral pneumonia is a major manifestation of human H5N1
disease, and is a common cause of death of patients with influenza
virus infection (1). Yet, the
biological basis for the severity of human H5N1 disease and fatal
cases of the novel influenza A (H1N1) virus infection remains
unclear.
High levels of interferon-γ-inducible protein-10
(IP-10) have been observed in several viral diseases (2,3).
Infection of H5N1 virus was found to lead to the hyper-induction of
proinflammatory cytokines in human primary macrophage cultures
in vitro, and patients with H5N1 disease were found to have
unusually high serum concentrations of chemokine IP-10 (4). The hyper-induction of chemokines may
contribute to the severity of human H5N1 disease and this may be
relevant to the pathogenesis of pneumonia caused by influenza
virus.
To test the hypothesis whether IP-10 is an important
chemokine in the development of airway inflammation caused by
virus, mice were infected with influenza virus after administration
of murine IP-10, and the severity of pneumonia was compared to that
of the mouse group infected with influenza virus alone. Another
mouse group was infected with respiratory syncytial virus (RSV)
after being injected with murine IP-10, and also the inflammatory
response of the lung was compared to that of the mouse group
infected with RSV alone.
Materials and methods
Mice
Pathogen-free 8-week-old female BALB/c mice were
purchased from the Animal Center of China Medical University. Mice
were bred and/or housed under specific, pathogen-free conditions
until the day of sacrifice. The institutional ethics committee for
animal use and care approved all animal-related experiments and
procedures.
IP-10 administration to mice
Murine recombinant IP-10 was purchased from Shanghai
BioSun Sci & Tech Co., Ltd. Mice received intraperitoneal
(i.p.) injections of 30 μg/kg IP-10 in 0.2 ml of
phosphate-buffered saline (PBS; Gibco/BRL) on day 0–2 of virus
infection.
Viral infection
Mouse-adapted influenza A/FM/1/47 H1N1 viruses were
cultivated in the allantoic cavity of 10-day-old chicken embryos by
incubating infected embryos at 34°C for 2–3 days. RSV was
propagated in HeLa cells.
Under light anesthesia, mice were inoculated
intranasally (i.n.) with 50 μl of 1:10 diluted influenza
virus stock containing 104 50% egg lethal dose (ELD50)
per milliliter of allantoic fluid or 107 plaque forming
units (PFUs) of RSV in a volume of 50 μl. For influenza
virus-infected mice, Group 1 was infected with influenza virus
without administration of IP-10; Group 2 was infected with
influenza virus after administration of IP-10; Group 3 was
mock-infected with PBS; and Group 4 was injected with IP-10 without
virus infection. For RSV-infected mice, Group 1 was infected with
RSV alone and Group 2 was infected with RSV after administration of
IP-10; Group 3 was mock-infected. At 5 days post-infection, mice
were euthanized. The number of mice per group was 10.
Measurement of lung weight
The mice were sacrificed by cervical dislocation.
Lungs were removed, and the lung index of lungs, i.e., lung
weight/body weight × 100, was determined as an indication of lung
inflammation, as previously described (5).
Pulmonary histopathology
The left lobe of the lung was perfused, fixed in 10%
buffered formalin and embedded in paraffin. Mutiple sections
(4-μm) were stained with H&E. Slides were analyzed and
scored for cellular inflammation under light microscopy by two
independent pathologists, as previously described (6). Inflammatory infiltrates were scored
by enumerating the layers of inflammatory cells surrounding the
vessels, bronchioles and alveolar infiltration. An overall severity
score was calculated for each animal by adding the individual
scores. Histopathology scores for animal from two independent
experiments were pooled and averaged, and the standard error of the
mean (SEM) was calculated. Lung histopathological results of the
virus infection group and IP-10 administration/virus infection
group were compared using the Mann-Whitney U test. Differences were
considered significant at a P-value <0.05.
Results
Lung index
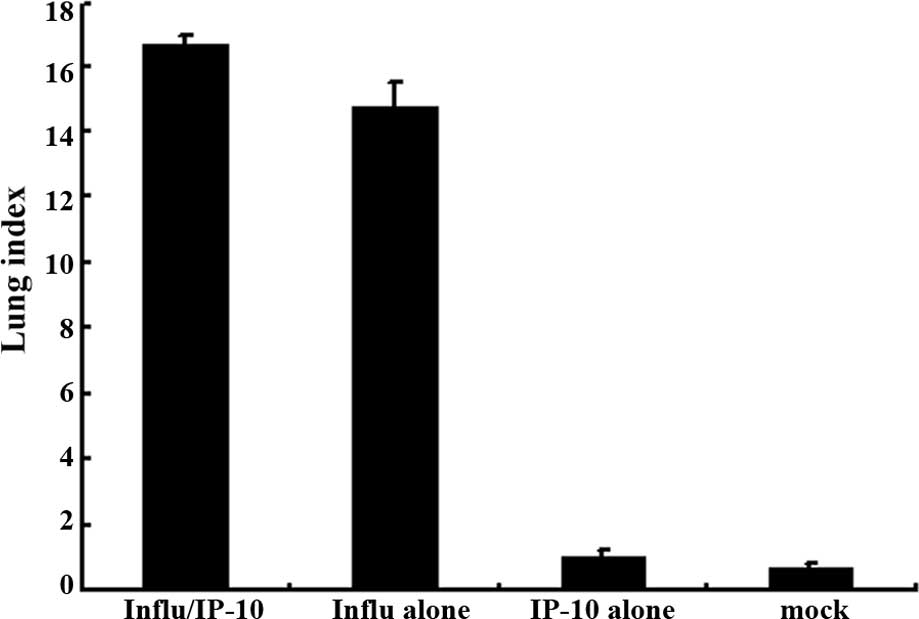
Fig. 1 shows the
lung index in the different groups of mice. The extent of
inflammation was evaluated by the increase in lung weight using the
lung index. There was a significant increase in the lung index in
Group 2 mice which were infected with the influenza virus after
administration of IP-10 (Fig. 1A)
and in Group 2 mice infected with RSV after administration of IP-10
(Fig. 1B).
Lung inflammation in the influenza
virus-infected mice
A well-known function of chemokines is to drive
leukocyte migration through tissues to target microenvironments.
Therefore, the histological appearance of the lungs and the degree
of inflammation were investigated. Group 2 (8-week-old female
BALB/c mice) was infected with influenza virus after administration
of murine IP-10 and the result was compared with Group 1 mice which
were infected with influenza virus without IP-10 administration and
the mock-infected Group 3. On day 5 post-infection, multiple
H&E-stained lung sections were analyzed and inflammation was
scored using a previously described scoring scale (6).
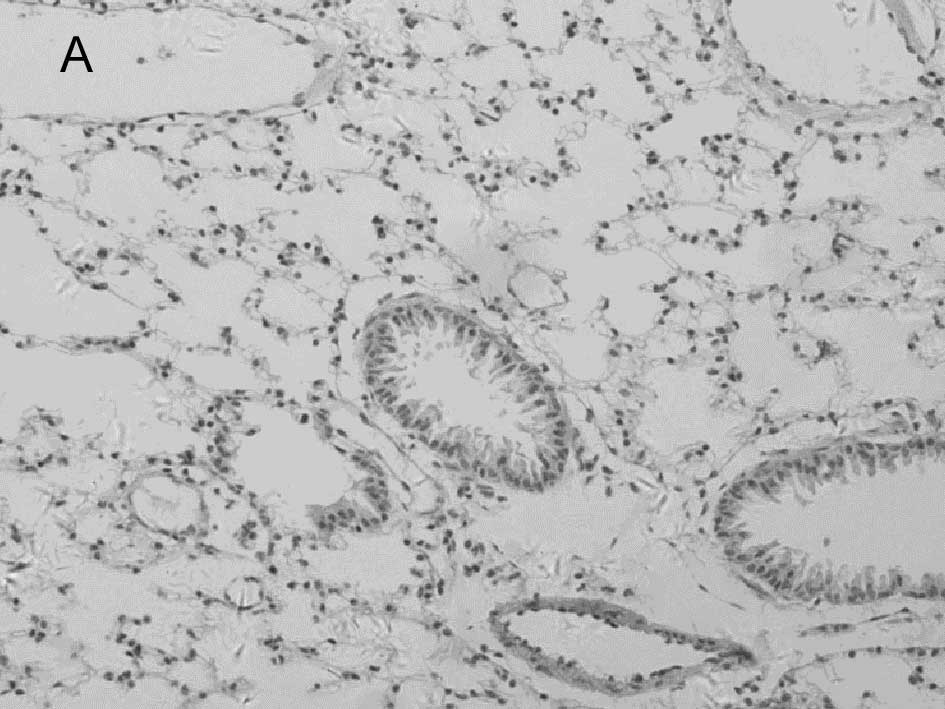
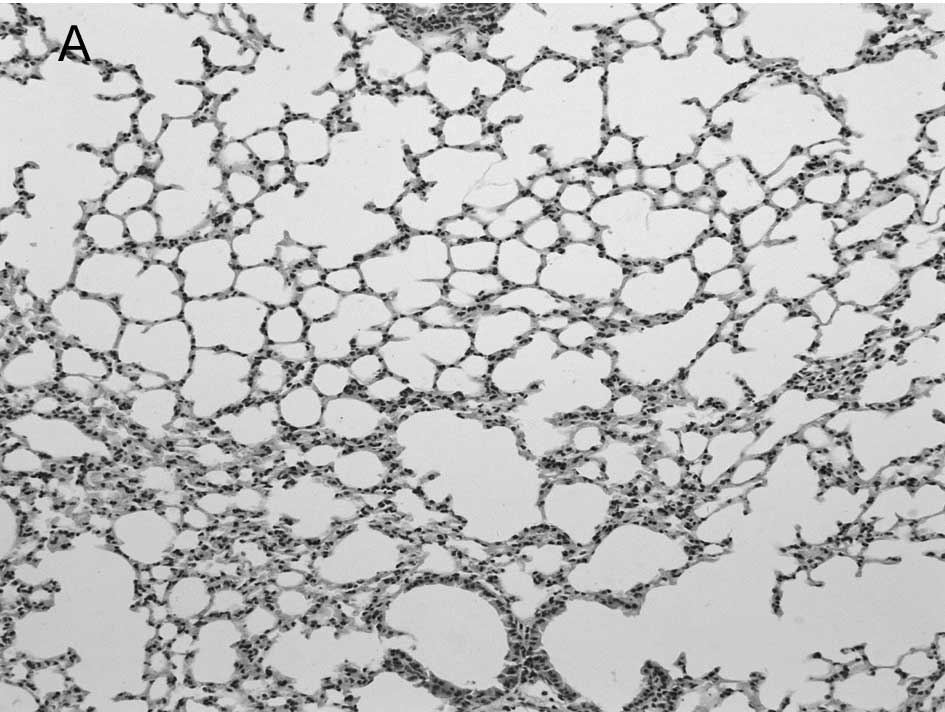
In the mock-infected mice (Fig. 2A) and Group 4 which was injected
with IP-10 alone (data not shown), few infiltrating cells were
found in interstitial/alveolar ducts and around bronchioles or
vessels. The mice in Group 2 which were infected with influenza
virus after administration of IP-10, had a greater degree of
interstitial and alveolar inflammation (Fig. 2C) compared to the mice in Group 1
(Fig. 2B) which were infected with
influenza virus alone. The pulmonary pathology of Group 2 was acute
exudative diffuse alveolar damage with hyaline membrane formation
and the major cell types of infiltration were lymphocytes and, to a
lesser extent, macrophages. The peribronchiolar inflammation in
Group 2 was also much more severe compared to Group 1, with diffuse
damage of structures.
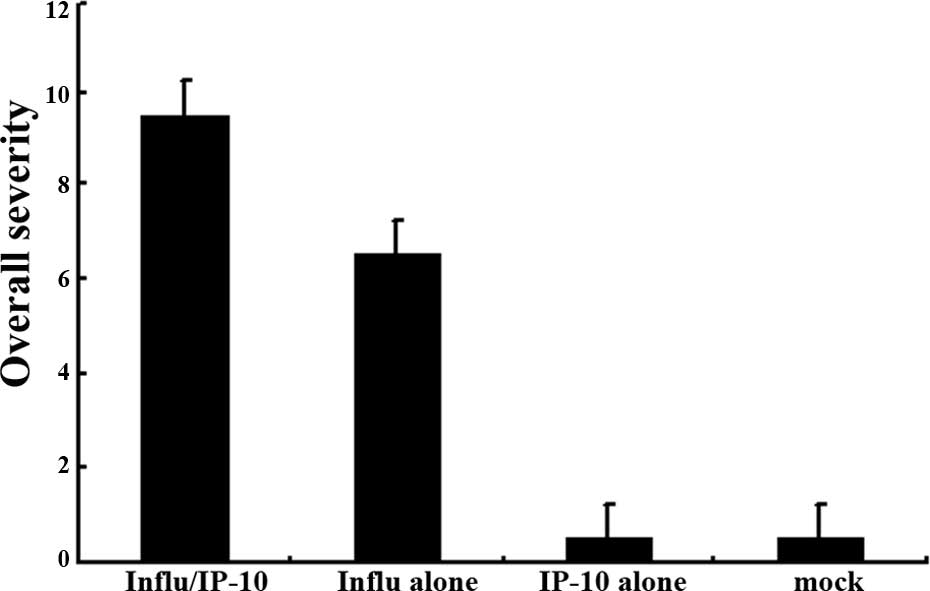
The overall severity scores differed significantly
(P<0.05) between the two groups (Groups 1 and 2), and the
overall severity score indicated a trend toward a greater magnitude
in Group 2 (Fig. 4A).
Lung inflammation in RSV-infected
mice
To survey the pulmonary inflammation of mice
infected with RSV after administration of murine IP-10, the
histological appearance of the lung and the degree of inflammation
of RSV-infected mice were investigated. The mice were also divided
into three groups similar to the influenze-infected mice. The
RSV-infected and IP-10-injected mice (Group 2) were compared to
Group 1 mice which were infected with RSV without IP-10
administration and mock-infected Group 3. On day 5 post-infection,
multiple H&E-stained lung sections were analyzed, and
inflammation was scored using a scoring scale as described
above.
Group 2 mice which were infected with RSV virus
after administration of IP-10 had a greater degree of interstitial
and alveolar inflammation (Fig.
3C) compared to Group 1 mice which were infected with RSV alone
(Fig. 3B) and mock-infected mice
(Fig. 3A). The pulmonary pathology
of Group 2 was acute exudative diffuse alveolar damage, but no
hyaline membrane formation was noted, and the major cell types
involved in infiltration were lymphocytes which was the same as the
influenza virus-infected group; the peribronchiolar inflammation in
this group was also much more severe compared to Group 1, with
damage of structures.
The overall severity scores differed significantly
(P<0.05) between the two groups (Groups 1 and 2) (Fig. 4B).
Discussion
Patients with H5N1 disease may present with primary
viral pneumonia complicated by syndromes of acute respiratory
distress and multiple organ dysfunction. Findings in the
respiratory tree consist of diffuse alveolar damage with
interstitial lymphoplasmacytic infiltrate and the formation of
hyaline membranes in alveoli/alveolar ducts in addition to
necrotizing bronchitis and bronchiolitis. In cases of coincident
bacterial pneumonia, a massive infiltration of neutrophils into
alveolar air spaces is observed. The interstitial pneumonia caused
by the H5N1 virus is much more severe compared to the pneumonia
caused by other types of influenza viruses (7). In the present study, intranasal
administration of influenza virus to mice led to the development of
pneumonia, characterized by alveolar damage with lymphoplasmacytic
infiltration and interstitial edema. A pseudostratified change in
columnar epithelial cells of bronchitis and bronchiolitis occurred.
The mock-infected group had no interstitial and alveolar
inflammation.
Chemokines are a large family of proteins that
mediate the movement and activation of diverse groups of
inflammatory cells. A subfamily of chemokines, including IP-10,
plays a key role in promoting type I immune responses. IP-10
contributes to lung inflammatory cell recruitment and activation
which directs CD8(+) T-cell recruitment to the lungs (8). IP-10 is an important factor in the
clearance of certain pathogens, but it also causes severe
inflammatory reaction in lungs and may be associated with pulmonary
pathological changes that cause acute respiratory distress in
patient. Dawson et al (9)
found IP-10 mRNA overexpression in the lungs after influenza virus
infection. Patients with H5N1 disease have unusually high serum
concentrations of chemokine IP-10, and the hyper-induction of this
chemokine may contribute to the unusual severity of human H5N1
disease (10). The results of the
present study showed that the mice infected with influenza virus
after administration of IP-10 suffered a more fulminant and
necrotizing diffuse alveolar and bronchiole damage with acute
exudative inflammation as well as hyaline membrane formation and
strongly reactive lymphocytes; the result also showed that the
pneumonia of this group was more severe compared to the group of
mice infected with influenza virus alone. The overall severity
scores were significantly higher for the group of IP-10-injected
mice. This result indicates that IP-10 plays an important role in
the development of severe pneumonia.
RSV is the most important cause of lower respiratory
tract illness in infants and children. The group of mice infected
with RSV after administration of IP-10 demonstrated a greater
degree of interstitial and alveolar inflammation than the group of
mice infected with RSV without IP-10 administration. These results
suggest that IP-10 is also associated with the severity of
pneumonia caused by RSV. However, RSV-induced pneumonia had less
interstitial and alveolar inflammation than that of the influenza
virus-infected group (the overall severity scores differed
significantly between the two groups; P<0.05). This indicates
that IP-10 may have a much more important correlation with the
severity of influenza virus-induced pneumonia. Zeng et al
(11) showed that IP-10 was an
important component of innate immunity against extracellular
bacterial pathogens of the lung. Nevertheless, in certain viral
pneumonias the level of IP-10 was significantly elevated and
produced immunopathological processes involved in lung injury
(12). In conclusion, IP-10 is an
important chemokine and is associated with the severity of
pneumonia caused by several types of viruses and may be used as a
candidate for immunoadjuvant therapy in the future.
References
|
1.
|
CY ChengLLM PoonAS LauW LukYL LauKF
ShortridgeY GuanJSM PeirisInduction of proinflammatory cytokines in
human macrophages by influenza A (H5N1) viruses: a mechanism for
the unusual severity of human
disease?Lancet36018311837200210.1016/S0140-6736(02)11772-712480361
|
|
2.
|
A AntonelliC FerriP FallahiSM FerrariM
SebastianiD FerrariM GiuntiS FrascerraS TolariF FranzoniF GalettaS
MarchiE FerranniniHigh values of CXCL10 serum levels in mixed
cryoglobulinemia associated with hepatitis C infectionAm J
Gastroenterol10324882494200810.1111/j.1572-0241.2008.02040.x18775023
|
|
3.
|
S BhowmickR DusejaS DasMB AppaiahgiriS
VratiA BasuInduction of IP-10 (CXCL10) in astrocytes following
Japanese encephalitisNeurosci
Lett4144550200710.1016/j.neulet.2006.11.07017287085
|
|
4.
|
MCW ChanCY CheungWH ChuiSW TsaoJM
NichollsYO ChanRWY ChanHT LongLLM PoonY GuanJSM
PeirisProinflammatory cytokine responses induced by influenza A
(H5N1) viruses in primary human alveolar and bronchial epithelial
cellsRespir Res6135147200510.1186/1465-9921-6-13516283933
|
|
5.
|
EM AllenVL MooreJO StevensStrain variation
in BCG-induced chronic pulmonary inflammation in mice. I. Basic
model and possible genetic control by non-H-2 genesJ
Immunol1193433471977
|
|
6.
|
AE KajonAP GigliottiKS HarrodAcute
inflammatory response and remodeling of airway epithelium after
subspecies B1 human adenovirus infection of the mouse lower
respiratory tractJ Med Virol71233244200310.1002/jmv.10475
|
|
7.
|
KT JefferyMM DavidThe pathology of
influenza virus infectionAnn Rev
Pathol3499522200810.1146/annurev.pathmechdis.3.121806.154316
|
|
8.
|
X ZengTA MooreMW NewsteadJC DengNW
LukacsTJ StandifordIP-10 mediates selective mononuclear cell
accumulation and activation in response to intrapulmonary
transgenic expression and during adenovirus-induced pulmonary
inflammationJ Interferon Cytokine
Res25103112200510.1089/jir.2005.25.103
|
|
9.
|
TC DawsonMA BeckWA KuzielF HendersonN
MaedaContrasting effects of CCR5 and CCR2 deficiency in the
pulmonary inflammatory response to influenza A virusAm J
Pathol15619511959200010.1016/S0002-9440(10)65068-710854218
|
|
10.
|
JSM PeirisWC YuCW LeungCY CheungWF NgJM
NichollsTK NgKH ChanST LaiWL LimKY YuenY GuanRe-emergence of fatal
human influenza A subtype H5N1
diseaseLancet363617619200410.1016/S0140-6736(04)15595-514987888
|
|
11.
|
X ZengTA MooreMW NewsteadJC DengSL
KunkelAD LusterTJ StandifordInterferon-inducible protein 10, but
not monokine induced by gamma interferon, promotes protective type
1 immunity in murine klebsiella pneumoniae pneumoniaInfect
Immun7382268236200510.1128/IAI.73.12.8226-8236.2005
|
|
12.
|
JY ChienPR HsuehWC ChengCJ YuPC
YangTemporal changes in cytokine/chemokine profiles and pulmonary
involvement in severe acute respiratory
syndromeRespirology11715722200610.1111/j.1440-1843.2006.00942.x17052299
|