Introduction
Lung cancer is the most common cause of
cancer-related mortality worldwide. The two major subtypes are
small-cell lung cancer (SCLC) and non-small-cell lung cancer
(NSCLC) (1,2). Early stages of NSCLC are treated with
curative surgical resection. However, over 60% of all NSCLC
patients already have advanced or metastatic tumors at the time of
diagnosis and are not suitable for surgery. The overall 5-year
survival rate of patients with NSCLC remains extremely poor
(3–5). Therefore, it is crucial to employ new
therapeutic strategies in the treatment of NSCLC. With the
constantly evolving knowledge of the molecular pathogenesis of lung
cancer, targeted therapies have recently been introduced (6). Moreover, antisense gene therapies,
such as small RNA interference (RNAi), have been evaluated in
recent years (7–10).
Cyclin D1 is a pivotal cell cycle-regulatory protein
that controls the cell cycle transition from G1 to S phase. It is
also intricately involved in the regulation of apoptosis depending
on the proliferative and differentiated state of the cell (11–14).
Overexpression of the Cyclin D1 protein was reported in various
types of cancer, such as breast, esophageal and lung cancer
(15–17). Betticher et al reported that
Cyclin D1 was associated with poor tumor differentiation and was
known to be a negative indicator in NSCLC (17). When Cyclin D1 is overexpressed, it
may increase the risk of tumor progression and early onset of
cancer (19–22). Cyclin D1 overexpression enhances
cell proliferation and cell cycle progression (19–22).
Certain studies have reported targeted therapy aimed at Cyclin D1
by small RNAi (18).
Down-regulation of the expression of Cyclin D1 inhibits tumor
growth (18). In this regard, we
hypothesized that the dysregulation of Cyclin D1 occurs relatively
early in the process of tumorigenesis and may be promising for
cancer therapy.
Bcl-xL is a critical member of the Bcl-2 family and
is correlated to several malignancies, including NSCLC (23–26).
As an anti-apoptotic protein, the overexpression of Bcl-xL may
inhibit the mitochondrial cytochrome release, which is a mechanism
by which cancer cells escape apoptosis and regulate the apoptosis
of two signaling pathways, the extrinsic or death receptor pathway
and the intrinsic or mitochondrial pathway (23). Previous studies have also reported
targeted therapy aimed at Bcl-xL by small RNAi (25,26).
Substantial research has shown that the down-regulation of
anti-apoptotic gene expression is capable of sensitizing cancer
cells to anticancer drugs and promoting cell apoptosis (25,26).
Thus, Bcl-xL is a potential new therapeutic target in NSCLC.
As noted, a number of studies have reported RNAi
aimed at Cyclin D1 or Bcl-xL. However, the effect of combining the
two genes in vitro for an intervention study is unclear in
NSCLC. Biliran et al have reported that the expression of
Bcl-xL remained relatively high in the cells with overexpressed
Cyclin D1 (27). Huang et
al have reported that combined therapy with the two genes
prolonged survival in mice with ovarian cancer (28). Thus, we formulated a hypothesis
that combined interference of the two genes is a promising new
strategy for improving lung cancer outcomes.
In the present study, we aimed to determine whether
combined interference was superior to single interference.
Materials and methods
Construction of shRNA vectors
The pcDNA6.2-GW/EmGFP-miR vector was purchased from
Invitrogen (Carlsbad, CA, USA) with a genetically engineered
improved murine miR-155 skeleton structure containing a terminal
loop and an internal loop. The recombinant plasmid
pcDNA6.2-GW/EmGFP-miR that expressed a cytomegalovirus (CMV)
promoter-driven micro30 short hairpin RNA (shRNA) targeting Cyclin
D1 (Cyclin D1 shRNA), Bcl-xL (Bcl-xL shRNA) and a combination of
the two genes (Cyclin D1-Bcl-xL shRNA) were constructed,
respectively. The micro30 shRNA reverse sequencing primer site (C)
occurred at bases 1607–1626. Green fluorescent protein (GFP) assays
were implemented by co-transfection of cancer cells with plasmids
encoding GFP and corresponding shRNA in order to observe the
efficacy of transfection. The shRNAs were designed to target human
Bcl-xL (accession no. NM_138578.1) and Cyclin D1 (accession no.
NM_053056.2) mRNA. The sequences are shown in Table I. The micro30 shRNAs were
synthesized by Invitrogen.
 | Table I.The sequences of micro30 shRNA sense
strands. |
Table I.
The sequences of micro30 shRNA sense
strands.
| Gene | Sequence |
|---|
| MR075-3-F |
TGCTGTGTAGATGCACAGCTTCTCGGGTTTTGGCCACTGACTGACCCGAGAAGGTGCATCTACA |
| MR075-3-R |
CCTGTGTAGATGCACCTTCTCGGGTCAGTCAGTGGCCAAAACCCGAGAAGCTGTGCATCTACAC |
| MR076-1-F |
TGCTGAGAGAAAGTCAACCACCAGCTGTTTTGGCCACTGACTGACAGCTGGTGTGACTTTCTCT |
| MR076-1-R |
CCTGAGAGAAAGTCACACCAGCTGTCAGTCAGTGGCCAAAACAGCTGGTGGTTGACTTTCTCTC |
Cell culture
The human lung adenocarcinoma cell lines A549 and
NCI-H441 were cultured in RPMI-1640 (Invitrogen) containing 10%
fetal bovine serum (Invitrogen). The stock was maintained in a 5%
CO2 incubator at 37°C in a humidified atmosphere. The
cancer cells were divided into four groups, i.e., blank control
(untreated cells), Cyclin D1 shRNA, Bcl-xL shRNA and Cyclin
D1-Bcl-xL shRNA (transfected cells).
Plasmid transfection
The plasmids were transfected into A549 and NCI-H441
cells according to the manufacturer’s instructions for
Lipofectamine™2000 (Invitrogen). Briefly, prior to transfection,
2–3×105 cells in 2 ml of growth medium without
antibiotics were placed in 6-well plates. The 6-well plates were
washed with 2 ml OptiMEM (Gibco, Invitrogen, USA) twice, and
another 2 ml OptiMEM containing 4 μg pcDNA6.2-GW/EmGFP-miR mixed
with 10 μl of lipofectamine was added to form liposomes. Following
incubation at 37°C for 4–6 h, the 6-well plates were placed in 2 ml
medium containing 10% FBS in a 5% CO2 incubator at 37°C
in a humidified atmosphere overnight.
Real-time reverse
transcription-polymerase chain reaction (RT-PCR)
The transfected cells were collected for real-time
RT-PCR. Transfected and untreated cells were collected and washed
with phosphate-buffered saline (PBS). Total RNA was extracted from
the cells of the four groups using TRIzol reagent (Invitrogen) in a
single-step method and cDNA was generated with a
PrimeScript® RT reagent kit (Takara, Shiga, Japan) at a
total volume of 20 μl according to the manufacturer’s instructions.
Expression levels of target gene were normalized to the
housekeeping gene β-actin (ΔCt). Gene expression values
were then calculated based on the ΔΔCt method using the
equation: RQ=2−ΔΔCt. PCR amplification was
performed with SYBR® Premix Ex Taq™ (Takara), under the
following PCR conditions: 95°C for 30 sec, followed by 40 cycles of
95°C for 5 sec, 60°C for 30 sec, and a dissociation stage of 95°C
for 15 sec, 60°C for 1 min, and 95°C for 15 sec. The real-time
primer sequences were designed and synthesized by Takara Customer
Services (Dalian, China). The primer sequences used were: human
β-actin, sense: 5′-GCAAGCAGGAGTATGACGAG-3′ and antisense:
5′-CAAATAAAGCCATGCCAATC-3′ (144 bp); Cyclin D1, sense:
5′-ATGTTCGTGGCCTCTAAGATGA-3′ and antisense:
5′-CAGGTTCCACTTGAGCTTGTTC-3′ (138 bp); Bcl-xL, sense:
5′-AGCTTGGATGGCCACTTACCTG-3′ and antisense:
5′-TGCTGCATTGTTCCCATAGAGTTC-3′ (100 bp). Each assay was performed
in triplicate and repeated three times.
Western blot analysis
Transfected cells were collected for Western blot
analysis. Following transfection with shRNAs 72 h later, the cells
were lysed in EBC buffer with protease inhibitor on ice and
centrifuged at 10,000 x g for 10 min at 4°C. Lysates were separated
by 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel
electrophoresis (PAGE) and then transferred electrophoretically
onto polyvinylidene fluoride (PVDF) membranes. The membranes were
blocked in PBS-T containing 5% non-fat dry milk and 0.1% Tween-20
for 2 h at room temperature. Subsequently, the membranes were
washed with PBS-T containing 0.1% Tween-20 and incubated with
rabbit anti-human Bcl-xL antibody (Cell Signaling, Beverly, MA,
USA) at a dilution of 1:1000, and mouse anti-human Cyclin D1
antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at a
dilution of 1:500 overnight at 4°C. The membranes were then
incubated with horseradish peroxidase (HRP)-conjugated secondary
antibodies. The peroxidase activity was detected using the enhanced
chemiluminescence system (Pierce, Rockford, IL, USA), according to
the manufacturer’s instructions.
Cell proliferation analysis
To determine the effect of Cyclin D1 and Bcl-xL
shRNAs on cell proliferation, cells were seeded in 12-well plates
at a density of 20×104 cells/well and cultivated with
RPMI-1640 in the CO2 incubator at 37°C. Following
transfection with Cyclin D1 and Bcl-xL shRNAs 48 h later, untreated
cells and the transfected cells were collected. The total cell
number was determined with a hematocytometer under an inverted
microscope. Each assay was performed in triplicate and repeated
three times.
Dimethylthiazol-diphenyltetrazolium
bromide (MTT) assay
Cell proliferation was assessed by the MTT assay.
A549 and NCI-H441 cells were seeded in 96-well plates and
transfected with shRNAs for 48 h. Then, 10 μl of 5 mg/ml MTT (in
PBS) was added to each well and continually incubated for 4 h at
37°C. The formazan granules obtained from cells were dissolved in
150 μl dimethyl sulfoxide (DMSO) for 10 min. Cell viability was
then measured in terms of optical density (OD) at a wavelength of
490 nm. Each cell viability assay was performed in quadruplicate
and repeated three times.
Annexin V/propidium iodide (PI)
staining
To determine the effect of Cyclin D1 and Bcl-xL
shRNAs on cell apoptosis, cells were seeded in 6-well plates at a
final concentration of 3×105/ml and transfected with
those plasmids, respectively. Cells were collected, washed with PBS
and successively resuspended in 500 μl binding buffer. Cells were
incubated with 5 μl FITC-conjugated human Annexin V (KeyGen
Biotech, Nanjing, China) in the dark for 15 min at 4°C and then
stained with 5 μl PI (KeyGen Biotech). After 10 min, samples were
immediately analyzed with a FACSCalibur (BD Biosciences, San Jose,
USA) flow cytometer. Each assay was performed in triplicate and
repeated three times.
Statistical analysis
Data were expressed as the mean ± standard deviation
(SD). Differences in the data were analyzed by ANOVA. P<0.05 was
considered to be statistically significant. Statistical analysis
was carried out using SPSS 13.0 software.
Results
Effects of three shRNAs on Cyclin D1 and
Bcl-xL expression in cancer cells
The plasmids were transfected into A549 and NCI-H441
cells. The efficacy of transfection was detected by measuring the
percentage of the fluorescent cells, which were transfected by the
plasmids containing the GFP gene. Results showed the efficacy of
transfection of these plasmids in the A549 and NCI-H441 cells to be
>70%.
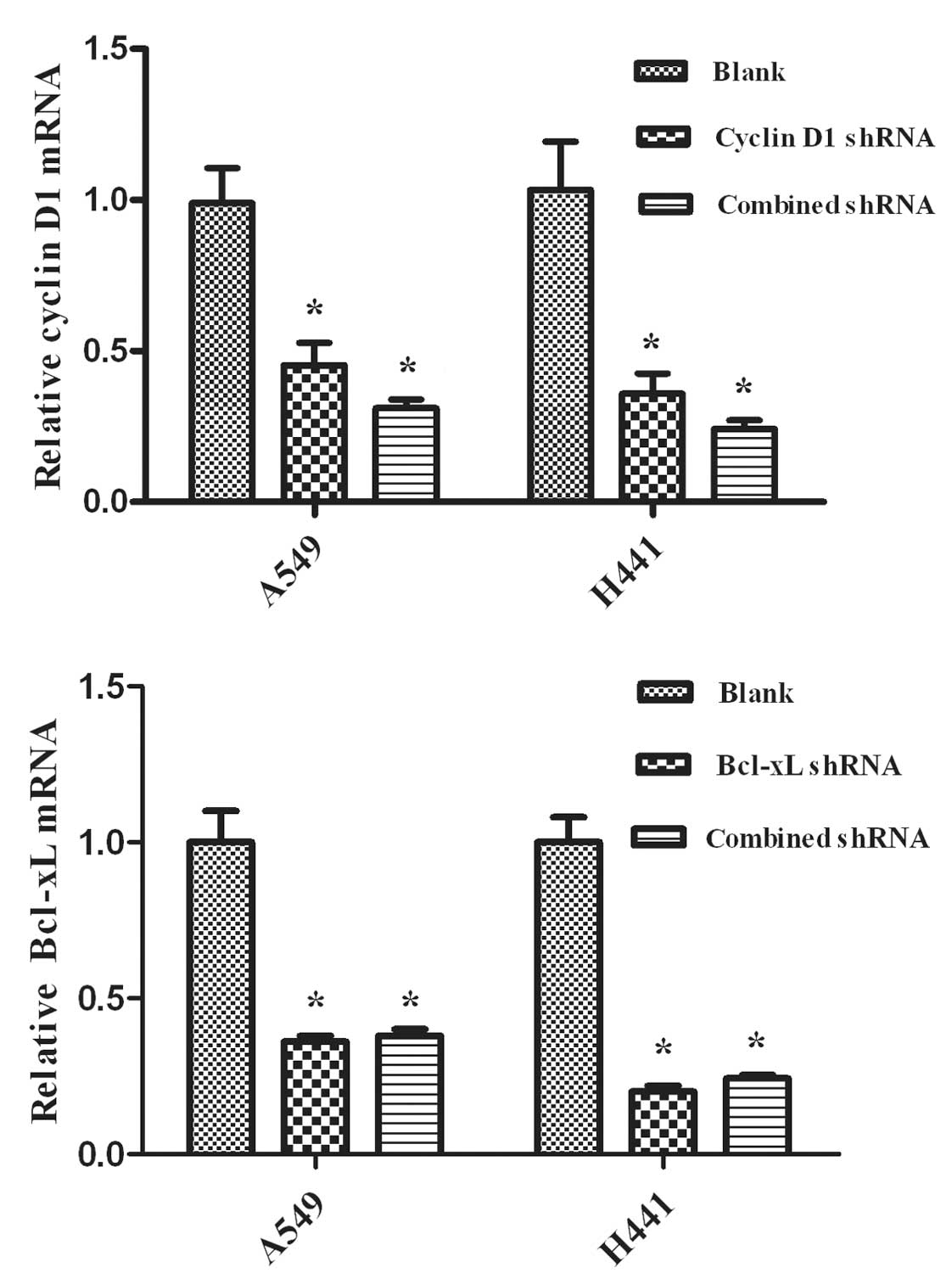
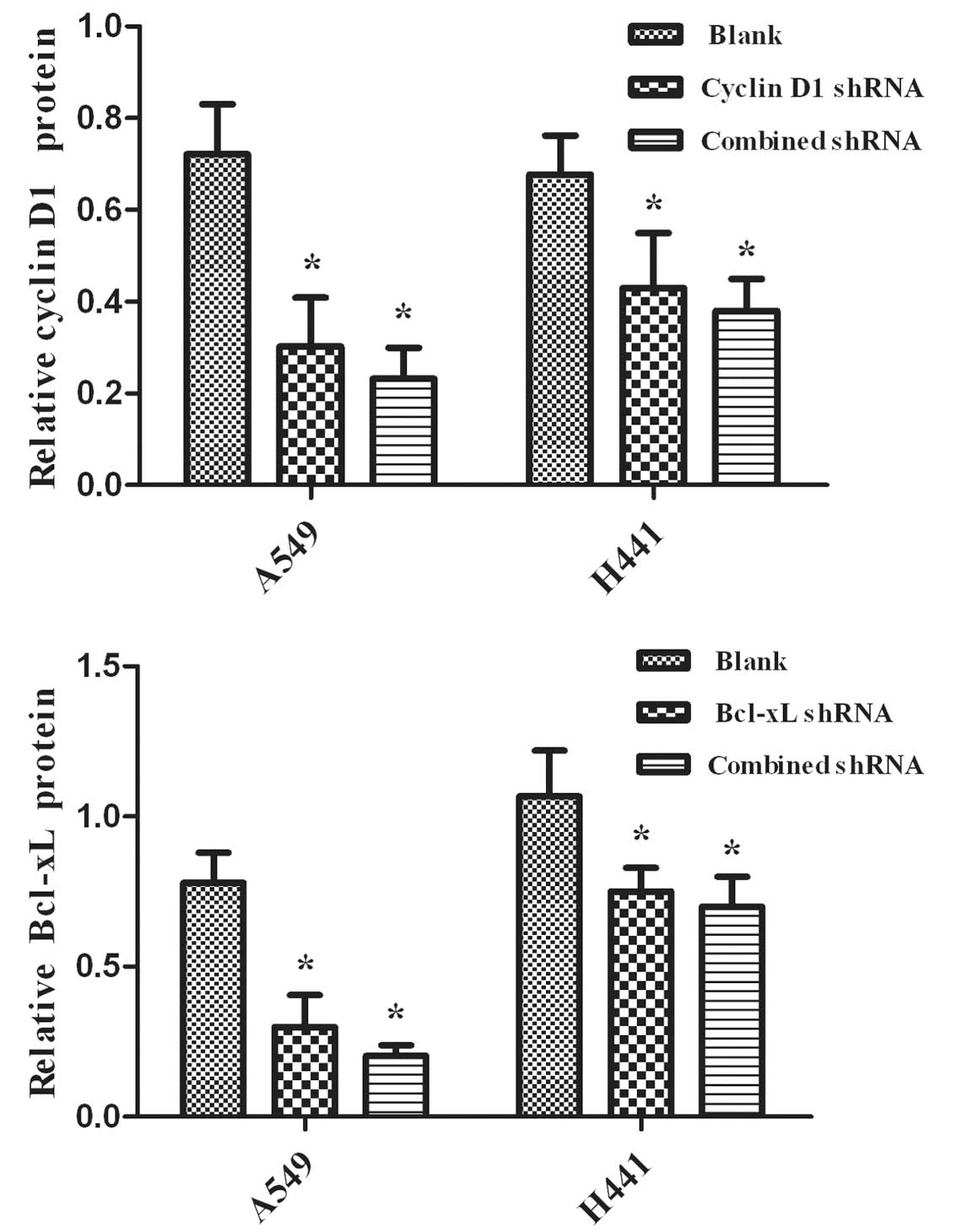
The silencing effects of RNAi were evaluated using
real-time RT-PCR analysis and Western blot analysis to detect the
levels of mRNA and protein, respectively. We observed the
expression of mRNA and the proteins in NSCLC A549 and NCI-H441
cells. Cyclin D1 and Bcl-xL shRNAs are capable of down-regulating
the expression levels of mRNA (Fig.
1) and protein (Fig. 2)
(P<0.05), while the blank control showed no effects (P>0.05).
Additionally, no significant differences were observed between the
combined and single interference groups regarding Cyclin D1 or
Bcl-xL, respectively (P>0.05).
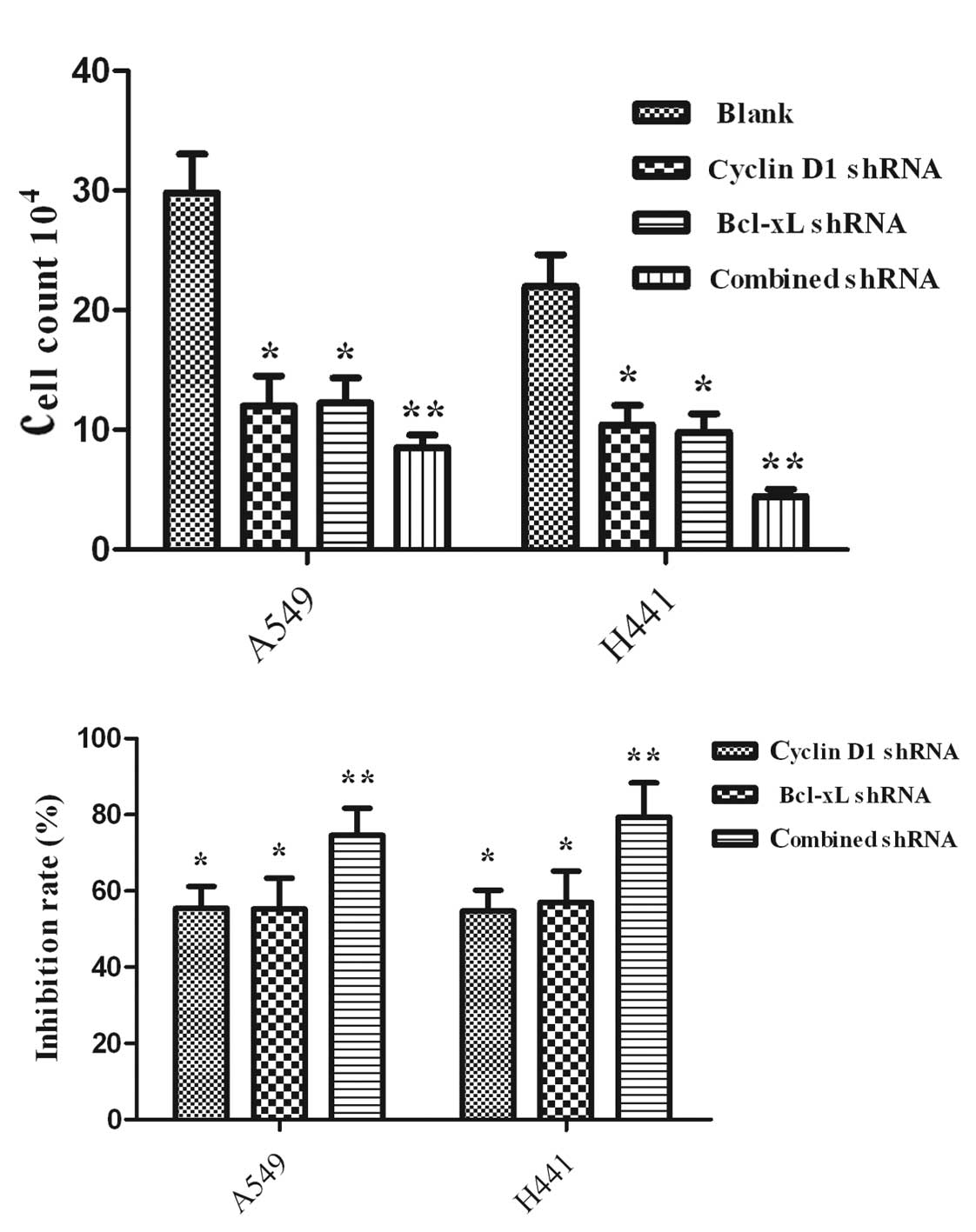
Induction of proliferation in A549 and
NCI-H441 cells by treatment with RNAi
Proliferation was assessed by the cell count
(Fig. 3A) and MTT assay (Fig. 3B). shRNA-treated cells exhibited a
significant decrease compared with the untreated cells (P<0.05).
Furthermore, the proliferation efficacy of the group with combined
interference had a significant decrease compared with the groups of
Cyclin D1 shRNA- and Bcl-xL shRNA-treated cells (P<0.05).
Induction of apoptosis in A549 and
NCI-H441 cells by treatment with RNAi
Apoptosis was assessed by annexin V/PI nuclear
staining. As shown in Fig. 4,
combined targeted interference significantly promoted cell
apoptosis compared with single interference (P<0.05). The three
interference groups also had statistically significant promotion of
cell apoptosis compared with the blank control (P<0.05).
Discussion
Findings of the present study showed that the cell
cycle-regulatory molecule Cyclin D1 and the anti-apoptotic protein
Bcl-xL were overexpressed in A549 and NCI-H441 lung cancer cells.
Thus, Cyclin D1 and Bcl-xL may be potential therapeutic targets in
NSCLC. The down-regulation of Cyclin D1 and Bcl-xL in NSCLC by
single interference has been reported (18,25,26).
We performed a combined intervention of the two genes, and the
results demonstrated that the combined intervention was more
effective in promoting cell apoptosis and reducing cell
proliferation in the NSCLC A549 and NCI-H441 cell lines than single
intervention.
Previous studies have confirmed that combinational
interference with shRNA and chemo/radiation therapy may increase
the efficacy of individual therapy (28,29).
To increase the efficacy of shRNA therapy, combinational therapy
(double or triple therapy) may be a novel strategy for cancer
treatment (30,31).
In the present study, we first successfully
constructed pcDNA6.2-GW/EmGFP-miR vectors expressing Bcl-xL shRNA,
Cyclin D1 shRNA and Cyclin D1-Bcl-xL shRNA, respectively. The
results indicated that Bcl-xL shRNA, Cyclin D1 shRNA and Cyclin
D1-Bcl-xL shRNA efficiently inhibited the expression of the target
genes, as demonstrated by the mRNA and protein levels shown in
Figs. 2 and 3. The protein expression levels of Bcl-xL
and Cyclin D1 were significantly reduced in the groups of Cyclin D1
shRNA, Bcl-xL shRNA and Cyclin D1-Bcl-xL in A549 and NCI-H441 cells
compared with the blank group. The results of quantitative RT-PCR
analysis of Bcl-xL and Cyclin D1 revealed mRNA variations that
generally correlated with the Western blotting data in our study.
RT-PCR and Western blot analysis further indicated that
shRNA-transfected cells had successfully silenced the target
gene.
Cyclin D1 shRNA transfection resulted in marked
changes in the levels of proliferation and apoptosis in A549 and
NCI-H441 cells. A number of gene therapy strategies targeting
Cyclin D1 have been used in vitro and in vivo, and
have been shown to suppress tumor growth and promote tumor
apoptosis in lung cancer (32,33).
Driscoll et al have reported that Cyclin D1 antisense
oligonucleotide-transfected A549 and NCI-H441 cells exhibited a
reduced growth rate with a range of 40–60% at 0–8 day growth
curves, which was consistent with our results (32). However, Huang et al have
reported cell proliferation assay analysis performed at 1-, 3-, 5-
and 7-day time points (18). These
authors found that cells transfected with Cyclin D1-targeted shRNA
exhibited a significant decrease in cell proliferation only at the
7-day time point (P<0.05), and there was no significant
difference at the 1-, 3- and 5-day time points (P>0.05). We
analyzed the possible reasons for these differences. Firstly, the
intervention methods utilized are different. Oligonucleotide
transfection efficacy was higher than plasmid transfection
efficacy. Secondly, cell proliferation and apoptosis were related
to the time of interference. We detected the proliferation and
apoptosis of lung cancer cells within 48 h following
transfection.
Bcl-xL has been found to be overexpressed in various
types of cancer, such as lung and prostate cancer (25,34).
Kim et al have proven that Bcl-xL as an anti-apoptotic
protein may result in a multiple drug-resistant phenotype (35). Antisense oligonucleotide-directed
Bcl-xL has been shown to cause sensitization to chemotherapy and
significant apoptosis in NSCLC (36,37).
Lei et al have reported that inhibition of Bcl-xL small
interfering RNA (shRNA) on the cisplatin (DDP)-resistant human lung
adenocarcinoma cell line A549/DDP was 12.65–58.75% (25). It is thus further confirmed that
Bcl-xL may be a potent intervention target in NSCLC.
Cyclin D1-Bcl-xL shRNA transfection was more
effective in promoting cell apoptosis and reducing cell
proliferation compared with single shRNA transfection. The
inhibition rate of the combined intervention was less than the sum
of the individual Cyclin D1 shRNA and Bcl-xL shRNA groups. However,
apoptosis of the Cyclin D1-Bcl-xL shRNA group was 14.3% more than
the sum of the Cyclin D1 shRNA (4.835%) and Bcl-xL shRNA (5.41%) in
the A549 cell groups. A similar result was achieved in the NCI-H441
cells with Cyclin D1-Bcl-xL shRNA (15.5%), Bcl-xL shRNA (6.2%) and
Cyclin D1 shRNA (5.7%). The change in apoptosis and growth
suppression in the Cyclin D1-Bcl-xL shRNA group was not consistent.
This inconsistency may be due to the fact that Cyclin D1 and Bcl-xL
play roles in carcinogenesis through different molecular mechanisms
with apoptosis inhibitors and cell-cycle regulators, respectively
(7,23). A number of studies have reported
that activation of the signal transducer and activator of
transcription (STAT) 3, a potent transcription factor, is capable
of suppressing apoptosis by mediating survival gene products
including Bcl-xL and may lead to cell proliferation through its
ability to induce the expression of Cyclin D1 in NSCLC (38–40).
Combined interference using Cyclin D1 and Bcl-xL may affect STAT3
signaling, resulting in growth suppression that is less than the
sum of single interference. Additionally, besides its role in cell
cycle regulation, Cyclin D1 is intricately involved in the
regulation of apoptosis. The effect of Cyclin D1 may be
proapoptotic or antiapoptotic, depending on the different state of
the cell (17,27). That may account for the combined
interference using Cyclin D1 and Bcl-xL promoting apoptosis more
effectively than the effect of the sum of single interference.
Whether the two genes are capable of acting synergistically
requires further investigation. Biliran et al have reported
that the expression of cell survival proteins, such as bcl-xL,
remained relatively high in Cyclin D1-overexpressing cells
(27). Therefore, combined
interference using Cyclin D1 and Bcl-xL may be more effective than
targeted single interference.
In conclusion, we have shown that Cyclin D1 and
Bcl-xL were overexpressed in NSCLC A549 and NCI-H441 cells, and
that the molecular-targeted repression of the combined Cyclin D1
and Bcl-xL genes was a more effective therapeutic strategy for
NSCLC than the down-regulation of either single gene in promoting
cell apoptosis and reducing cell proliferation. Combined
interference on Cyclin D1 and Bcl-xL may therefore be a potent
target strategy in NSCLC therapy. Although our findings in
vitro were only from two cell lines, these results may have
significant implications. Further in vivo studies are
required to elucidate whether combined interference of Cyclin D1
and Bcl-xL may also more effectively inhibit tumors. In future, for
effective in vivo delivery, tumor specificity, TNM staging
and non-specific immune responses should be overcome before this
technology may be successfully used in clinical research.
Acknowledgements
This work was supported by the
Ministry of Education Scientific Research Foundation for Returned
Overseas Students (no. 20071108) and the Natural Science Foundation
of Hubei Province (no. 2010CDB09304).
References
|
1.
|
PC HoffmanAM MauerEE VokesLung
cancerLancet355479485200010.1016/S0140-6736(00)82038-310841143
|
|
2.
|
SF AltekruseCL KosaryM KrapchoN NeymanR
AminouW WaldronSEER cancer statistics review 1975–2007Available at:
http://seer.cancer.Gov/csr/1975-2007.
|
|
3.
|
A JemalR SiegelE WardY HaoJ XuT MurrayMJ
ThunCancer statisticsCA Cancer J Clin5871962008
|
|
4.
|
TE StinchcombeMA SocinskiCurrent
treatments for advanced stage none small cell lung cancerProc Am
Thorac Soc6233241200910.1513/pats.200809-110LC
|
|
5.
|
WR SmytheTreatment of stage I non-small
cell lung
carcinomaChest123181S187S200310.1378/chest.123.1_suppl.181S12527578
|
|
6.
|
V HirshSystemic therapies in metastatic
non-small-cell lung cancer with emphasis on targeted therapies: the
rational approachCurr
Oncol171323201010.3747/co.v17i2.54920404973
|
|
7.
|
AS LundbergRA WeinbergControl of the cell
cycle and apoptosisEur J
Cancer35531539199910.1016/S0959-8049(99)00292-010492624
|
|
8.
|
MF BuckleyKJ SweeneyJA HamiltonExpression
and amplification of cyclin genes in human breast
cancerOncogene82127213319938336939
|
|
9.
|
S RamalingamCP BelaniRecent advances in
targeted therapy for non-small cell lung cancerExpert Opin Ther
Targets1124557200710.1517/14728222.11.2.24517227238
|
|
10.
|
M StevensonTherapeutic potential of RNA
interferenceN Engl J
Med35117721777200410.1056/NEJMra04500415496626
|
|
11.
|
CJ SherrG1 phase progression: cycling on
cueCell79551555199410.1016/0092-8674(94)90540-17954821
|
|
12.
|
V BaldinJ LukasMJ MarcoteM PaganoG
DraettaCyclin D1 is a nuclear protein required for cell cycle
progression in G1Genes Dev7812821199310.1101/gad.7.5.8128491378
|
|
13.
|
D ResnitzkyM GossenH BujardSI
ReedAcceleration of the G1/S phase transition by expression of
cyclins D1 and E with an inducible systemMol Cell
Biol141669167919948114703
|
|
14.
|
EK HanSC NgN ArberM BegemannIB
WeinsteinRoles of cyclinD1 and related genes in growth inhibition,
senescence and
apoptosisApoptosis4213219199910.1023/A:100961882414514634283
|
|
15.
|
C GillettV FantlR SmithAmplification and
overexpression of cyclinD1 in breast cancer detected by
immunohistochemical stainingCancer Res541812181719948137296
|
|
16.
|
W JiangSM KahnN TomitaAmplification and
expression of the human cyclinD gene in esophageal cancerCancer
Res522980298319921533816
|
|
17.
|
DC BetticherJ HeighwayPS
HasletonPrognostic significance of CCND1 (cyclinD1) overexpression
in primary resected non-small-cell lung cancerBr J
Cancer73294300199610.1038/bjc.1996.528562333
|
|
18.
|
H HuangYD HuN LiY ZhuInhibition of tumor
growth and metastasis by non-small cell lung cancer cells
transfected with cyclin D1-targeted
siRNAOligonucleotides19151162200910.1089/oli.2008.017419355812
|
|
19.
|
DM BarnesCE GillettCyclin D1 in breast
cancerBreast Cancer Res Treat52115199810.1023/A:1006103831990
|
|
20.
|
J BartkovaJ LukasH MüllerD LützhøftM
StraussJ BartekCyclin D1 protein expression and function in human
breast cancerInt J
Cancer57353361199410.1002/ijc.29105703118168995
|
|
21.
|
S GansaugeF GansaugeM RamadaniH StobbeB
RauN HaradaHG BegerOverexpression of cyclin D1 in human pancreatic
carcinoma is associated with poor prognosisCancer
Res571634163719979134998
|
|
22.
|
DW StaceyCyclin D1 serves as a cell cycle
regulatory switch in actively proliferating cellsCurr Opin Cell
Biol15158163200310.1016/S0955-0674(03)00008-512648671
|
|
23.
|
XM YinSignal transduction mediated by Bid,
apro-death Bcl-2 family proteins, connects the death receptor and
mitochondria apoptosis pathwaysCell
Res10161167200010.1038/sj.cr.729004511032168
|
|
24.
|
B Karczmarek-BorowskaA FilipJ
WojcierowskiA SmolenE KorobowiczI Korszen-PileckaM ZdunekEstimation
of prognostic value of Bcl-xL gene expression in non-small cell
lung cancerLung
Cancer516169200610.1016/j.lungcan.2005.08.01016297499
|
|
25.
|
X LeiZ HuangM ZhongB ZhuS TangD LiaoBcl-xL
small interfering RNA sensitizes cisplatin-resistant human lung
adenocarcinoma cellsActa Biochim Biophys Sin
(Shanghai)39344350200710.1111/j.1745-7270.2007.00286.x17492131
|
|
26.
|
Y SasazawaY FutamuraE TashiroM
ImotoVacuolar H+-ATPase inhibitors overcome Bcl-xL-mediated
chemoresistance through restoration of a caspase-independent
apoptotic pathwayCancer Sci100146014672009
|
|
27.
|
H Biliran JrY WangS BanerjeeH XuH HengA
ThakurA BolligFH SarkarJD LiaoOverexpression of cyclin D1 promotes
tumor cell growth and confers resistance to cisplatin-mediated
apoptosis in an elastase-myc transgene-expressing pancreatic tumor
cell lineClin Cancer
Res1160756086200510.1158/1078-0432.CCR-04-241916115953
|
|
28.
|
X HuangT LinJ GuL ZhangJA RothLC StephensY
YuJ LiuB FangCombined TRAIL and Bax gene therapy prolonged survival
in mice with ovarian cancer xenograftGene
Ther913791386200210.1038/sj.gt.330181012365003
|
|
29.
|
CA SledzBR WilliamsRNA interference and
double stranded-RNA-activated pathwaysBiochem Soc
Trans32952956200410.1042/BST032095215506933
|
|
30.
|
MJ BuenoI Pérez de CastroM
MalumbresControl of cell proliferation pathways by microRNAsCell
Cycle731433148200810.4161/cc.7.20.683318843198
|
|
31.
|
M JovanovicMO HengartnermiRNA and
apoptosis: RNAs to die
forOncogene2561766187200610.1038/sj.onc.120991217028597
|
|
32.
|
B DriscollL WuS BuckleyFL HallKD AndersonD
WarburtonCyclin D1 antisense RNA destabilizes pRb and retards lung
cancer cell growthAm J Physiol273L941L94919979374720
|
|
33.
|
ER SauterM HerlynSC LiuS LitwinJA
RidgeProlonged response to antisense cyclin D1 in a human squamous
cancer xenograft modelClin Cancer Res6654660200010690551
|
|
34.
|
EM BruckheimerBT GjertsenTJ
McDonnellImplications of cell death regulation in the pathogenesis
and treatment of prostate cancerSemin Oncol26382398199910482181
|
|
35.
|
IK KimYK JungDY NohYS SongCH ChoiBH OhES
MasudaYK JungFunctional screening of genes suppressing
TRAIL-induced apoptosis: distinctin hibitory activities of Bcl-XL
and Bcl-2Br J Cancer88910917200310.1038/sj.bjc.660079512644829
|
|
36.
|
SH LeechRA OlieO GautschiInduction of
apoptosis in lung cancer cells following bcl-xl antisense
treatmentInt J
Cancer86570576200010.1002/(SICI)1097-0215(20000515)86:4%3C570::AID-IJC20%3E3.0.CO;2-T10797273
|
|
37.
|
S WangD YangME LippmanTargeting Bcl-2 and
Bcl-XL with nonpeptidic small-molecule antagonistsSemin
Oncol30133142200310.1053/j.seminoncol.2003.08.01514613034
|
|
38.
|
X ZhangJ ZhangL WangH WeiZ TianTherapeutic
effects of STAT3 decoy oligodeoxynucleotide on human lung cancer in
xenograft miceBMC Cancer7149200710.1186/1471-2407-7-14917683579
|
|
39.
|
P WeerasingheGE GarciaQ ZhuP YuanL FengL
MaoN JingInhibition of Stat3 activation and tumor growth
suppression of non-small cell lung cancer by G-quartet
oligonucleotidesInt J Oncol31129136200717549413
|
|
40.
|
X ZhangJ ZhangH WeiZ TianSTAT3-decoy
oligodeoxynucleotide inhibits the growth of human lung cancer via
down-regulating its target genesOncol Rep1713771382200717487394
|