Introduction
Breast cancer is the most common type of cancer that
affects females (1). The interplay
between genetic and epigenetic events, as well as environmental
risk factors, has significant implications in the pathogenesis of
breast cancers (2). Current
research is concerned with identifying new genetic, epigenetic,
prognostic and predictive factors. It has been shown that genetic
factors (mutations in BRCA1/2) and reproductive history account for
one-third of all breast cancer cases. However, in two-thirds of
breast cancer cases the etiology remains unclear. Previous
epidemiological studies have indicated that certain environmental
agents may play an important role in the development of breast
carcinomas (3). Thus, the capacity
to metabolize and detoxify exogenous toxins may correlate with an
individual’s susceptibility to environment-induced breast cancer.
The GSTP1 gene is involved in a wide range of detoxification
reactions which protect cells from carcinogens (3,4).
GSTs provide protection against the electrophilic metabolites of
carcinogens and reactive oxygen species. GSTP1 is a
biotransformation enzyme expressed in normal breast epithelial
cells. High levels of GSTP1 have been associated with a poor
prognosis in breast cancer (3).
Silencing of tumor-suppressor genes through the
hyper-methylation of their promoter regions is a frequent event in
carcinogenesis (5).
Hypermethylation of CpG islands in the gene promoter regions of
numerous tumor suppressor and DNA repair genes has been reported to
be associated with events such as chromatin condensation,
replication delay and gene silencing (6,7).
Identification of epigenetic changes and their correlation with
clinical factors may lead to improvements in breast cancer
diagnosis and treatment. The 5’ region of GSTP1 is rich in
CpG islands and its methylation causes changes in expression levels
in neoplastic cells, as has been reported in a number of published
studies (8–15). GSTP1 promoter
hypermethylation is also associated with a loss of GSTP1 expression
(13,15). Studies have investigated the
methylation status of GSTP1 in invasive breast cancer
(9,16–19)
and a different study revealed GSTP1 promoter methylation to
be an early event in breast cancer (10). GSTP1 promoter methylation
has also been reported to be associated with a poor prognosis in
breast cancer (16).
GSTP1 has a polymorphic site at codon 105 in
exon 5, where an adenosine to guanosine (A>G) transition results
in an Ile to Val substitution (I105V), giving rise to the
GSTP1*B allele (20). Individuals with the valine allele
exhibit significantly lower enzyme activity and a reduced
detoxification ability (21).
There are five classes of GST enzymes (α, µ, π, σ and θ) in humans.
Studies have been published concerning the potential effects of the
changes to the activation and detoxification abilities of GST class
π enzymes on an individual’s risk of breast cancer and have
established an association between the GSTP1 Ile105Val polymorphism
and breast cancer risk (22–24).
Previously (25,26), we demonstrated that homozygous
mutant individuals have a significantly higher risk of breast
cancer. Therefore, we investigated the hypothesis that epigenetic
modification in homozygous mutants with reduced enzymatic activity
increases the risk of breast cancer, which is further modified by
various clinicopathological parameters, lifestyle factors and
dietary habits.
Materials and methods
Subjects and sample collection
A total of 215 breast cancer tissue samples and
corresponding blood samples were obtained from patients who
underwent surgery at the Institute Rotary Cancer Hospital (All
India Institute of Medical Sciences, New Delhi, India) between
September 2006 and May 2009. An additional 215 blood samples (10
ml) from healthy donors were also obtained. Patient and control
subject characteristics were as described previously (25). The study was approved by the
University and Hospital’s Human Ethics Committees. Patients and
controls provided written informed consent prior to their
enrollment in the study. Fresh tissue samples obtained from the
surgical specimens were snap-frozen in liquid nitrogen and stored
at −80°C until further use. The tissue samples were categorized on
the basis of tumor advancement according to the conventional
staging system described previously (27,28).
GSTP1 polymorphism analysis
The exon 5 Ile105Val polymorphism in the
GSTP1 gene was detected using PCR-restriction fragment
length polymorphism (PCR-RFLP) as described previously (25). DNA from the patient and control
blood samples was isolated and amplified using GSTP1 primers
and PCR products were digested with BsmAI. Digested PCR
products were then run on 2% agarose gel and at least 25% of the
samples were repeated to confirm the results.
GSTP1 promoter hypermethylation
analysis
Genomic DNA isolated from tissue biopsies was
modified by sodium bisulfite using an EZ DNA Methylation-Gold kit
according to the manufacturer’s instructions (ZYMO Research
Corporation, Irvine, CA, USA). The primer sequences used in the
present study are listed in Table
I (19). The PCR was set-up by
adding 2 μl of bisulfite modified DNA to a PCR mix containing 1X
PCR buffer, dNTPs, 1 unit Platinum Taq DNA polymerase (Life
Technologies, Inc., Rockville, MD, USA) and primers (300 ng each
per reaction) to make a final volume of 25 μl. Methylation-specific
PCR (MSP) was carried out using the conditions described previously
(25). A negative (no DNA) and
positive control (universal methylated DNA) were included for each
PCR set. PCR products were analyzed by running on a non-denaturing
6% polyacrylamide gel stained with ethidium bromide and visualized
under UV illumination.
 | Table IMethylation-specific primer sequences
for GSTP1. |
Table I
Methylation-specific primer sequences
for GSTP1.
| Oligonucleotide
name | Primer
sequence |
|---|
| GSTP1 Unmethylated
(Forward primer) | 5′-GAT GTT TGG GGT
GTA GTG GTT GTT-3′ |
| GSTP1 Unmethylated
(Reverse primer) | 5′-CCA CCC CAA TAC
TAA ATC ACA ACA-3′ |
| GSTP1 Methylated
(Forward primer) | 5′-TTC GGG GTG TAG
CGC TCG TC-3′ |
| GSTP1 Methylated
(Reverse primer) | 5′-GCC CCA ATA CTA
AAT CAC GAC G-3′ |
Immunohistochemical analysis
Immunohistochemical analysis was performed on tissue
sections placed on poly-L-lysine-coated slides. Sections were
de-paraffinized in xylene and rehydrated in decreasing gradients of
alcohol, followed by the inhibition of endogenous peroxidase
activity (0.3% hydrogen peroxide in methanol) for 45 min. For
antigen retrieval, slides were placed in citrate buffer (pH 6.0)
and boiled at maximum power for 25 min in a microwave oven. The
tissue sections were washed with PBS for 10 min, incubated
overnight at 4°C with primary anti-GSTP1 antibody (1:200 dilution;
#SC-66000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,USA) in a
moist chamber according to the manufacturer’s guidelines,
counterstained with hematoxylin, dehydrated and mounted for
analysis. For the negative controls, sections were treated
similarly with the exception of the addition of the primary
antibody. Assessment of GSTP1 expression was performed under a
microscope at x400 magnification.
Western blot analysis
The amount of GSTP1 was detected by resolving the
cell lysates on 12% SDS-PAGE at a 1:50 dilution. After performing
SDS-PAGE, the gel was transferred to the transfer tank buffer for
20 min, followed by the transfer of proteins onto the
nitrocellulose membrane using semi-dry gel transfer apparatus
(Bio-Rad, Hercules, CA, USA) at 13 V for 40 min. The nitrocellulose
membrane was then placed in blocking buffer (PBS, 1.0% BSA and
0.05% Tween-20) and incubated at room temperature for 1 h with
constant agitation. The membrane was washed three times for 10 min
per wash using wash buffer (PBS, 0.1% BSA and 0.05% Tween-20). The
primary antibody anti-GSTP1was then diluted in wash buffer
(1:5,000) before being added and incubated for 1 h; the membrane
was washed 3 times. Secondary anti-mouse antibody diluted in wash
buffer (1:5,000) was added to the membrane and incubated for 1 h at
room temperature. The membrane was washed three times and developed
until bands appeared. The reaction was stopped by adding 100 mM
EDTA. These western blot membranes were scanned for density
measurements using the Quantity One software of the Gel
Documentation 2000 System (Bio-Rad).
Statistical analysis
The odds ratios (ORs) and 95% confidence intervals
(CIs) were calculated to measure the association between the
GSTP1 genotype and the risk of breast cancer. Associations
between the GSTP1 genotype and methylation status, as well
as any correlation between GSTP1 methylation and GSTP1
expression, were examined using the Chi-square test and Fisher’s
exact test (SAS Institute, Cary, NC, USA).
Results
Polymorphism in GSTP1 genotypes
A total of 37.7% (81/215) of breast cancer patients
were homozygous for the wild-type allele, 41.4% (89/215)
heterozygous (Ile/Val) and 20.9% (45/215) homozygous (Val/Val) for
the mutant allele. Similarly, we found the distribution of the
GSTP1 genotype in the controls to be 47% (101/215) wild-type
homozygous genotypes, 34.8% (75/215) heterozygous and 18.1%
(39/215) homozygous mutant genotypes. However, no significant risk
of breast cancer among individuals carrying the mutant allele was
observed (Table II; OR, 1.48; 95%
CI, 0.97–2.26 for heterozygotes; OR, 1.42; 95% CI, 0.86–2.42 for
homozygous mutants).
| Table IIAssociation between the GSTP1
genotype and breast cancer. |
Table II
Association between the GSTP1
genotype and breast cancer.
| GSTP1
genotype | Cases (n=215), n
(%) | Controls (n=215), n
(%) | OR (95% CI) | P-valuea |
|---|
| Ile/Ile (A/A) | 81 (37.7) | 101 (47) | 1 (Reference) | |
| Ile/Val (A/G) | 89 (41.4) | 75 (34.9) | 1.48
(0.97–2.26) | 0.07 |
| Val/Val (G/G) | 45 (20.9) | 39 (18.1) | 1.44
(0.86–2.42) | 0.19 |
GSTP1 promoter hypermethylation
MSP was employed to study GSTP1 promoter
hypermethylation. Aberrant promoter hypermethylation was observed
in 74 (34.4%) cases and the remaining 141 (65.6%) were unmethylated
(Table III, Fig. 1). A total of 28 of the 74
methylated cases belonged to the early stage of breast cancer group
while 46 were from the locally advanced disease group. Fisher’s
exact test was used to determine the association between
hypermethylation of the GSTP1 gene with the clinical stages
of primary breast cancer patients. In the two disease groups,
GSTP1 methylation was a significant event (in early breast
cancer cases; P= 0.05; in locally advanced breast cancer; P= 0.03;
Table IV). No statistically
significant associations were observed between GSTP1
methylation and various clinicopathological parameters (Table V) such as menopausal status and
progesterone receptor (PR) and estrogen receptor (ER) status. The
association of various lifestyle factors with GSTP1 promoter
hypermethylation was also studied, including age, smoking status,
age of menarche onset, contraceptive use, family history of cancer,
number of children, ER and PR status and alcohol intake.
Methylation of the GSTP1 gene was significantly associated
with ER+/PR+ status (Table VI). The disease risk was increased
by >2.5 times compared with ER−/PR− tissue
(OR, 2.63; 95% CI, 1.18–5.87; P=0.01; Table VI). When the results were analyzed
with regard to ER and PR status with age (<50 or >50 years),
the disease risk was increased by 2.5 times in the <50 years of
age group (OR, 2.47; 95% CI, 0.99–6.19; Table VII). Similarly, when ER/PR status
was considered in females <50 years old, the risk was increased
by nearly three times when both ER and PR status were positive (OR,
2.92; 95% CI, 1.03–8.31; Table
VII). However, none of the other factors were significantly
associated with GSTP1 promoter hypermethylation (data not
shown).
| Table IIIDistribution of methylated status
among various genotypes of GSTP1 in breast cancer cases. |
Table III
Distribution of methylated status
among various genotypes of GSTP1 in breast cancer cases.
| Genotype | Methylated
(n=74) | Unmethylated
(n=141) | OR (95% CI) |
|---|
| GSTP1 A/A
(81) | 29 | 52 | 1.0
(Reference) |
| GSTP1 A/G
(89) | 30 | 59 | 0.91
(0.48–1.72) |
| GSTP1 G/G
(45) | 15 | 30 | 0.89
(0.42–1.93) |
| Table IVStage-specific methylation patterns
of GSTP1 gene. |
Table IV
Stage-specific methylation patterns
of GSTP1 gene.
| Tumor stage | Methylated | Unmethylated | Total (%) | P-valuea |
|---|
| Early | 28 | 89 | 117 (54.4) | 0.05 |
| Locally
advanced/metastatic | 46 | 52 | 98 (45.6) | 0.03 |
| Total | 74 | 141 | 215 | |
| Table VGSTP1 methylation status
stratified on the basis of various clinicopathological
features. |
Table V
GSTP1 methylation status
stratified on the basis of various clinicopathological
features.
| Clinicopathological
features | Total | Methylated | Unmethylated | P-valuea |
|---|
| Menopause
status | | | | |
|
Premenopausal | 132 | 50 | 82 | 0.19 |
|
Postmenopausal | 83 | 24 | 59 | |
| PR status | | | | |
| Positive | 121 | 45 | 76 | 0.39 |
| Negative | 94 | 29 | 65 | |
| ER status | | | | |
| Positive | 140 | 54 | 86 | 0.09 |
| Negative | 75 | 20 | 55 | |
| Table VIAssociation of GSTP1
methylation with ER and PR status |
Table VI
Association of GSTP1
methylation with ER and PR status
| ER and PR
status | Total | Methylated | Unmethylated | OR (95% CI) |
|---|
| ER− and
PR− | 48 | 11 | 37 | 1 (Reference) |
| ER− and
PR+ | 38 | 9 | 29 | 1.04
(0.38–2.86) |
| ER+ and
PR− | 47 | 18 | 29 | 2.09
(0.85–5.11) |
| ER+ and
PR+ | 82 | 36 | 46 | 2.63
(1.18–5.87)a |
| Table VIIGSTP1 methylation as per ER
and PR status and age distribution. |
Table VII
GSTP1 methylation as per ER
and PR status and age distribution.
| ER and PR
status | Age (years) | Methylated | Unmethylated | OR (95% CI) |
|---|
| ER+ and
PR+ | >50 | 11 | 24 | 1 (Reference) |
| ≤50 | 25 | 22 | 2.47
(0.99–6.19)a |
| ER+ and
PR+ | ≤50 | 7 | 18 | 1 (Reference) |
| ER+ and
PR+ | >50 | 25 | 22 | 2.92
(1.03–8.31)a |
In order to test the hypothesis that somatic
epigenetic modification in homozygous mutants, combined with
reduced enzymatic activity, increases the risk of breast cancer an
attempt was made to determine the role of the GSTP1
polymorphism in promoter hypermethylation. However, no significant
associations of any of the three genotypes with promoter
hypermethylation were observed (Table
III). Therefore, it was concluded that promoter
hypermethylation in homozygous mutants with reduced enzymatic
activity due to the substitution mutation did not elevate the the
risk of breast cancer.
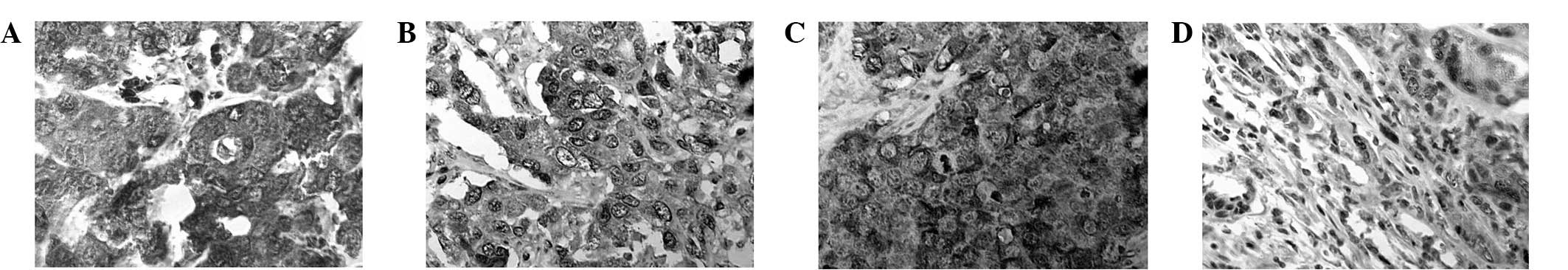
Immunohistochemical and western blot
analysis of GSTP1
Immunohistochemistry and western blot analysis were
performed on all the tissue samples to determine the correlation
between GSTP1 hypermethylation and GSTP1 expression.
Immunohistochemistry was employed to localize the protein and
western blotting to quantify protein expression. Representative
results of the GSTP1 immunohistochemistry and western blot analysis
are shown in Figs. 2 and 3, respectively, and the results obtained
in these two assays complemented each other. The
immunohistochemistry results did not contradict those of the
western blotting in any cases. A total of 67 tumors lacking GSTP1
expression exhibited GSTP1 promoter hypermethylation (100%)
and 7 tumors showed methylation but expressed GSTP1 (Table VIII; P<0.0005). The results
indicate that methylation does not always result in a loss of GSTP1
activity and there may be other mechanisms that contribute to its
activity. GSTP1 was expressed in all unmethylated samples. Strong
staining and high expression levels were found in all samples of
non-tumorous breast tissues.
| Table VIIIExpression analysis of GSTP1
gene in breast cancer patients according to GSTP1
methylation status. |
Table VIII
Expression analysis of GSTP1
gene in breast cancer patients according to GSTP1
methylation status.
| | GSTP1
immunoexpression
| |
|---|
| Breast
carcinoma | n | Present, n (%) | Absent, n (%) | P-value |
|---|
| GSTP1
methylated | 74 | 7 (9.5) | 67 (90.5) | 0.0005 |
| GSTP1
unmethylated | 141 | 141 (100) | 0 | |
Discussion
Glutathione S-transferases are phase II
metabolizing enzymes involved in the biotransformation of exogenous
substances, including mutagens, carcinogens and other poisonous
chemicals, and play a crucial role in the detoxification process,
thereby protecting cells from these compounds (29). GSTP1 codes for a GST π
family enzyme and is located on human chromosome 11q13.
Polymorphism in exon 5 (A105G) results in amino acid sequence
variation which affects enzyme activity as well as its substrate
specificity. Therefore, a detailed investigation of the effect of
this polymorphism on an individual’s susceptibility to various
environmentally generated tumors is required (30). Since a variety of environmental
factors may play a decisive role in causing breast cancer in the
majority of cases, the role of this polymorphism in breast cancer
appears to be important. In the present study, no evidence of an
increased risk of breast cancer in females carrying the variant
allele (Val) at codon 105 of GSTP1 was observed. Previously,
we demonstrated that the GSTP1 polymorphism is associated
with an increased risk of breast cancer (25) and therefore, the current hypothesis
was tested. Tissue samples were not available from all breast
cancer cases so the present study was carried out using 215 of the
413 cases reported previously and accordingly matched controls. It
is likely that the polymorphism results obtained in the present
study may not represent the real scenario since the patients’
menopausal status may have affected the present results compared
with the overall results obtained previously (25). In the present investigation, tissue
samples were obtained from 132 pre-menopausal patients and 83
post-menopausal patients. GSTP1 variants encode the change
at position 105 from isoleucine to valine. It is predicted that
this change may limit free access of the substrate to the H-site of
the enzyme which is required for conjugation with glutathione, as
well as thermostability (31,32).
This would result in reduced activity of the polymorphic protein.
However, the exact underlying biological mechanism is not
completely understood (3).
Since gene methylation generally mimics the mutation
pattern (33), the question of
whether methylation plays a role in genetic susceptibility to
breast cancer similar to that of the mutation (34) was also examined in the present
study. A previous study revealed an association between
GSTP1 methylation and tumor invasion, size, sentinel lymph
node metastasis and progression in breast cancer (19). In the present study, it was also
demonstrated that hypermethylation of GSTP1 occurs more
frequently in advanced stage cancer cases than in early disease
status cases. These findings suggest that in addition to a role in
tumorigenesis, GSTP1 methylation may also be associated with
disease progression.
GSTP1 is considered to act as a
tumor-suppressor gene, leading to tumor growth when it is
inactivated. It has been reported that GSTP1 interferes with
N-terminal c-Jun kinase signaling (35). If inactivated, GSTP1 may act
as a caretaker gene leading to additional somatic genome
alterations that promote tumor growth (36). GSTP1 binds non-covalently to
steroids, allowing it to act as an intracellular buffer to minimize
short-term fluctuations in steroid levels. The breast is an
important organ of the body which is continuously exposed to these
steroids and it is therefore highly likely that estrogens act as
endogenous tumor initiators in the breast tissue when GSTP1
is inactivated by promoter methylation. GSTP1 promoter
region inactivation by hypermethylation is a common event in cancer
and this epigenetic modification is often linked with a loss of
GSTP1 expression (9,12,18,37,38).
We observed that GSTP1 promoter hypermethylation is present
in one-third (34.4%) of human breast carcinomas, which was well
within the previous reported frequency range (13 to 38.9%)
(9,10,16–19).
Statistical analysis revealed an inverse correlation between
GSTP1 methylation and GSTP1 expression. Methylated samples
exhibited a loss of gene expression, suggesting that the silencing
of the GSTP1 gene by CpG island DNA methylation plays an
important role in the development of breast cancer. In unmethylated
samples an intense expression signal was obtained for all samples
using expression analysis techniques (see Materials and methods).
In previous studies, GSTP1 promoter hypermethylation has
been linked with gene silencing and reduced GSTP1 expression
(11,38) in breast cancer cells, whereas
normal mammary epithelial cells have always been found to express
GSTP1 (9). In certain cases, the
presence of GSTP1 expression despite GSTP1 methylation may
be attributed to a heterogeneous cell population in the tumor.
Based on our findings, we propose that genotyping and methylation
analysis may be considered together as a sensitive and accurate
diagnostic biomarker (39).
No association was found between GSTP1
promoter hypermethylation and the GSTP1 genotype, although
these mechanisms separately demonstrated a predisposition to breast
cancer in the present study and our previous study (25). In the present study, it is notable
that the methylation pattern of the GSTP1 gene was found to
be independent of age and menopausal status as reported in a
previous study (40).
It has long been recognized that estrogens are
linked to breast carcinogenesis (41) and that they cause genotoxic damage
(42). Methylation-mediated
silencing of the GSTPl gene may alleviate the role of
estrogen as an endogenous tumor initiator. In the present study
ER+/PR+ status was revealed to be
significantly associated with GSTP1 methylation in breast
cancer patients from northern parts of India. Significant
differences between ethnicities have been reported only in
ER−/PR− individuals aged less than 50 years
old in African-American and Caucasian females (43). Methylation frequencies were also
reported to be significantly higher in certain genes in Caucasian
females who are more than 50 years old with
ER+/PR+ than in their Korean counterparts.
These differences may be explained on the basis of the ethnicity of
the studied populations and possibly the associated risk factors.
The present study reported similar findings, as age was
significantly associated with methylation, as well as with ER and
PR status, even though the sample size was small. It is known that
estrogen action in normal and abnormal mammary cells is mediated by
ER and that ER is overexpressed in more than two-thirds of breast
cancer cases. Epigenetic modifications are potentially reversible
and therefore, identification of ethnicity-specific methylation
markers may be essential for cancer patient management and
therapeutic intervention. Our findings suggest that ER/PR-positive
tumors provide the possibility of developing biomarkers for
therapeutically targeting this population (44).
In the present study, promoter hypermethylation was
revealed to be the major factor underlying the loss of GSTP1
expression in breast carcinomas. However, our findings did not
demonstrate that epigenetic modification in homozygous mutant
individuals with reduced enzymatic activity increases the risk of
breast cancer. Our previous results and those of the present study
indicate that polymorphism and methylation are independent events
that individually affect the risk of breast cancer. These finding
may have implications for future chemotherapy regimens based on
epigenetic changes and ER/PR status.
Acknowledgements
The authors would like to thank the
Council of Scientific and Industrial Research, New Delhi (CSIR;
Grant No. 9=466 [87]-EMR-II) for financial assistance.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar
|
|
2
|
Widschwendter M and Jones PA: DNA
methylation and breast carcinogenesis. Oncogene. 21:5462–5482.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pongtheerat T, Pakdeethai S, Purisa W,
Chariyalertsak S and Petmitr S: Promoter methylation and genetic
polymorphism of glutathione S-transferase P1 gene (GSTP1) in Thai
breast-cancer patients. Asian Pac J Cancer Prev. 12:2731–2734.
2011.PubMed/NCBI
|
|
4
|
Lu S, Wang Z, Cui D, Liu H and Hao X:
Glutathione S-transferase P1 Ile105Val polymorphism and breast
cancer risk: a meta-analysis involving 34,658 subjects. Breast
Cancer Res Treat. 125:253–259. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fraga MF, Herranz M, Espada J, et al: A
mouse skin multistage carcinogenesis model reflects the aberrant
DNA methylation patterns of human tumours. Cancer Res.
64:5527–5534. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esteller M: Epigenetics in cancer. N Eng J
Med. 358:1148–1159. 2008. View Article : Google Scholar
|
|
8
|
Silvestrini R, Veneroni S, Benini E, et
al: Expression of p53 glutathione S-transferase-pi and Bcl-2
proteins and benefit from adjuvant radiotherapy in breast cancer. J
Natl Cancer Inst. 89:639–645. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esteller M, Corn PG, Urena JM, Gabrielson
E, Baylin SB and Herman JG: Inactivation of glutathione
S-transferase P1 gene by promoter hypermethylation in human
neoplasia. Cancer Res. 58:4515–4518. 1998.PubMed/NCBI
|
|
10
|
Lee JS: GSTP1 promoter hypermethylation is
an early event in breast carcinogenesis. Virchows Arch.
450:637–642. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Millar DS, Ow KK, Paul CL, Russell PJ,
Molloy PL and Clark SJ: Detailed methylation analysis of the
glutathione S-transferase pi (GSTP1) gene in prostate cancer.
Oncogene. 18:1313–1324. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jerónimo C, Varzim G, Henrique R, et al:
I105V polymorphism and promoter methylation of the GSTP1 gene in
prostate adeno-carcinoma. Cancer Epidemiol Biomarkers Prev.
11:445–450. 2002.PubMed/NCBI
|
|
13
|
Chan QK, Khoo US, Chan KY, et al: Promoter
methylation and differential expression of pi-class glutathione
S-transferase in endometrial carcinoma. J Mol Diagn. 7:8–16. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tokumaru Y, Harden SV, Sun DI, Yamashita
K, Epstein JI and Sidransky D: Optimal use of a panel of
methylation markers with GSTP1 hypermethylation in the diagnosis of
prostate adenocarcinoma. Clin Cancer Res. 10:5518–5522. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong S, Tang MW, Yeo W, Liu C, Lo YM and
Johnson PJ: Silencing of GSTP1 gene by CpG island DNA
hypermethylation in HBV-associated hepatocellular carcinomas. Clin
Cancer Res. 8:1087–1092. 2002.PubMed/NCBI
|
|
16
|
Arai T, Miyoshi Y, Kim SJ, Taguchi T,
Tamaki Y and Noguchi S: Association of GSTP1 CpG islands
hypermethylation with poor prognosis in human breast cancers.
Breast Cancer Res Treat. 100:169–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hoque MO, Feng Q, Toure P, et al:
Detection of aberrant methylation of four genes in plasma DNA for
the detection of breast cancer. J Clin Oncol. 24:4262–4269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parrella P, Poeta ML, Gallo AP, et al:
Nonrandom distribution of aberrant promoter methylation of
cancer-related genes in sporadic breast tumors. Clin Cancer Res.
10:5349–5354. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shinozaki M, Hoon DS, Giuliano AE, Hansen
NM, Wang HJ, Turner R and Taback B: Distinct hypermethylation
profile of primary breast cancer is associated with sentinel lymph
node metastasis. Clin Cancer Res. 11:2156–2162. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Henderson CJ, McLaren AW, Moffat GJ, Bacon
EJ and Wolf CR: Pi-class glutathione S-transferase: regulation and
function. Chem Biol Interact. 111–112:69–82. 1998.PubMed/NCBI
|
|
21
|
Hu X, Xia H, Srivastava SK, Pal A, Awasthi
YC, Zimniak P and Singh SV: Catalytic efficiencies of allelic
variants of human glutathione S-transferase P1-1 toward
carcinogenic anti-diol epoxides of benzo[c]phenanthrene and
benzo[g]chrysene. Cancer Res. 58:5340–5343. 1998.PubMed/NCBI
|
|
22
|
Antognelli C, Del Buono C, Ludovini V,
Gori S, Talesa VN, Crinò L, Barberini F and Rulli A: CYP17, GSTP1,
PON1 and GLO1 gene polymorphisms as risk factors for breast cancer:
an Italian case-control study. BMC Cancer. 9:1152009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reding KW, Weiss NS, Chen C, et al:
Genetic polymorphisms in the catechol estrogen metabolism pathway
and breast cancer risk. Cancer Epidemiol Biomarkers Prev.
18:1461–1467. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MARIE-GENICA Consortium on Genetic
Susceptibility for Menopausal Hormone Therapy Related Breast Cancer
Risk: Genetic polymorphisms in phase I and phase II enzymes and
breast cancer risk associated with menopausal hormone therapy in
postmenopausal women. Breast Cancer Res Treat. 119:463–474. 2010.
View Article : Google Scholar
|
|
25
|
Saxena A, Dhillon VS, Raish M, et al:
Detection and relevance of germline genetic polymorphisms in
glutathione S-transferases (GSTs) in breast cancer patients from
northern Indian population. Breast Cancer Res Treat. 115:537–543.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang BL, Sun T, Zhang BN, et al:
Polymorphisms of GSTP1 is associated with differences of
chemotherapy response and toxicity in breast cancer. Chin Med J
(Engl). 124:199–204. 2011.PubMed/NCBI
|
|
27
|
Singletary SE, Allred C, Ashley P, et al:
Revision of the American Joint Committee on Cancer staging system
for breast cancer. J Clin Oncol. 20:3628–3636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sobin LH and Fleming ID: TNM
classification of Malignant Tumors, 5th edition (1997). Union
Internationale Contre le Cancer and the American Joint Committee on
Cancer. Cancer. 80:1803–1804. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Strange RC, Spiteri MA, Ramachandran S and
Fryer AA: Glutathione-S-transferase family of enzymes. Mutat Res.
482:21–26. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kopps S, Angeli-Greaves M, Blaszkewicz M,
et al: Glutathione S-transfrease P1 Ile105Val polymoprphism in
occupationally exposed bladder cancer cases. J Toxicol Environ
Health A. 71:898–901. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bernardini S, Bellincampi L, Ballerini S,
et al: Glutathione S-transferase P1 *C allelic variant increases
susceptibility for late-onset Alzheimer disease: association study
and relationship with apolipoprotein E epsilon4 allele. Clin Chem.
51:944–951. 2005.
|
|
32
|
Johansson AS, Stenberg G, Widersten M and
Mannervik B: Structure activity relationships and thermal stability
of human glutathione transferase P1-1 governed by the H-site
residue 105. J Mol Biol. 278:687–698. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Baylin SB, Belinsky SA and Herman JG:
Aberrant methylation of gene promoters in cancer - concepts,
misconcepts, and promise. J Natl Cancer Inst. 92:1460–1461. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gazzoli I, Loda M, Garber J, Syngal S and
Kolodner RD: A hereditary nonpolyposis colorectal carcinoma case
associated with hypermethylation of the MLH1 gene in normal tissue
and loss of heterozygosity of the unmethylated allele in the
resulting microsatellite instability-high tumor. Cancer Res.
62:3925–3928. 2002.
|
|
35
|
Adler V, Yin Z, Fuchs SY, et al:
Regulation of JNK signaling by GSTp. EMBO J. 18:1321–1334. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kinzler KW and Vogelstein B:
Cancer-susceptibility genes. Gatekeepers and caretakers. Nature.
386:761–763. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee WH, Morton RA, Epstein JI, et al:
Cytidine methylation of regulatory sequences near the pi-class
glutathione S-transferase gene accompanies human prostatic
carcinogenesis. Proc Natl Acad Sci USA. 91:11733–11737. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brooks JD, Weinstein M, Lin X, et al: CG
island methylation changes near the GSTP1 gene in prostatic
intraepithelial neoplasia. Cancer Epidemiol Biomark Prev.
7:531–536. 1998.PubMed/NCBI
|
|
39
|
Tserga A, Michalopoulos NV, Levidou G,
Korkolopoulou P, Zografos G, Patsouris E and Saetta AA: Association
of aberrant DNA methylation with clinicopathological features in
breast cancer. Oncol Rep. 27:1630–1638. 2012.PubMed/NCBI
|
|
40
|
Matuschek C, Bölke E, Lammering G, et al:
Methylated APC and GSTP1 genes in serum DNA correlate with the
presence of circulating blood tumor cells and are associated with a
more aggressive and advanced breast cancer disease. Eur J Med Res.
15:277–286. 2010.PubMed/NCBI
|
|
41
|
Fishman J, Osborne MP and Telang NT: The
role of estrogen in mammary carcinogenesis. Ann NY Acad Sci.
768:91–100. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cavalieri EL, Stack DE, Devanesan PD, et
al: Molecular origin of cancer: catechol estrogen-3,4-quinones as
endogenous tumor initiators. Proc Natl Acad Sci USA.
94:10937–10942. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee JS, Lo PK, Fackler MJ, Argani P, Zhang
Z, Garrett-Meyer E and Sukumar S: A comparative study of Korean
with Caucasian breast cancer reveals frequency of methylation in
multiple genes correlates with breast cancer in young, ER,
PR-negative breast cancer in Korean women. Cancer Biol Ther.
6:1114–1120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cho YH, Shen J, Gammon MD, Zhang YJ, et
al: Prognostic significance of gene-specific promoter
hypermethylation in breast cancer patients. Breast Cancer Res
Treat. 131:197–205. 2012. View Article : Google Scholar : PubMed/NCBI
|