Introduction
Intestinal drug transporters have great potential
for drug absorption (1,2). They may serve as either drug targets
or drug delivery systems. It is generally assumed that at least 5%
of all human genes are associated with transporters which is
consistent with the biological significance of transporters and
their roles in cell homeostasis. The identification and
characterization of drug transporters has provided a scientific
basis for understanding drug delivery and disposition, the
molecular mechanisms of drug interactions and
inter-individual/inter-species differences (3). Various types of xenobiotic or drug
transporters have been identified as being important as barriers
against toxic compounds and influx pumps to take up nutrients into
the body. Since these xenobiotic transporters generally have a wide
range of recognition specificities and accept various types of
compounds as substrates, the localization and functional expression
of such transporters may be a critical factor in the disposition
and subsequent biological activity of therapeutic agents.
Dogs have emerged as a primary species for the study
of biology and human diseases (4).
The organization of the dog genome has been studied extensively in
the last ten years. With advances in genomics, microarray
technology has been used to identify tissue-specific genes,
including intestinal transporters. It is now accepted that the
process of drug absorption in the intestine is highly associated
with the functional expression of intestinal transporters (5). The current study was therefore
carried out to generate a gene expression database of transporters
in the canine duodenum.
Materials and methods
Materials
The TRIzol® reagent and SuperScript
Choice System cDNA synthesis kit were purchased from Invitrogen
(Carlsbad, CA, USA). The BioArray high-yield RNA transcript
labeling kit was obtained from Enzo Biochem (New York, NY, USA).
The RNeasy kit was supplied by Qiagen (Valencia, CA, USA). The
Canine 2.0 and Mouse 430A 2.0 GeneChips were provided by Affymetrix
(Santa Clara, CA, USA). GeneChip hybridization and scanning were
performed at the Seoulin Molecular Biology Technique Center (Seoul,
Korea).
Animals
Beagle dogs (4 males, 5.5–7.6 kg; Marshall
BioResource, Beijing, China) were housed in a controlled
semi-barrier system room at Chemon Co. (Yongin, Korea). The animals
were considered to be healthy based on clinical examination (Korea
Food and Drug Administration Guide for the Care and Use of
Laboratory Animals, 2009). ICR mice (30–40 g) were obtained from
Orient Bio Co. (Seoul, Korea) and housed in a controlled animal
room at Konkuk University (Seoul, Korea). The animals were fed
solid pellets and provided with water ad libitum. All
procedures were approved by the Konkuk University Institutional
Animal Care and Use Committee.
RNA isolation
Mucosal tissues obtained from dog and mouse
duodenums were immediately scraped with a clean glass slide,
transferred to a new frozen vial and dipped into liquid
N2. Tissue (∼100 mg) was added to 1 ml TRIzol reagent
and homogenized with a razor on ice. The homogenate was transferred
to a new tube and then 200 μl chloroform was added to the TRIzol
mixture. Following centrifugation at 12,500 rpm for 15 min at 4°C,
the aqueous phase was transferred to a new tube. The RNA was then
precipitated with 500 μl isopropanol and washed with 80% ethanol.
The RNA was further purified with an RNeasy Mini kit (Qiagen)
according to the manufacturer’s instructions. The concentration of
the purified RNA was measured at 260 nm and 5 μg purified RNA was
mixed with RNA loading buffer and heated at 75°C for 15 min. After
cooling on ice for 5 min, the RNA was loaded onto 1%
agarose/formaldehyde gel in 1X MOPS buffer. The gel was run at
80–100 V for 50 min and the presence of two sharp 18S and 28S bands
was confirmed under UV light.
Microarray assay
Once the total RNA samples were prepared, probe
synthesis, hybridization, detection and scanning were performed
according to the standard instructions of the manufacturer
(Affymetrix, Inc.). The cDNA was synthesized using the One-Cycle
cDNA Synthesis kit. Single-stranded cDNA was synthesized using
Superscript II reverse transcriptase and T7-oligo(dT) primers at
42°C for 1 h. Double-stranded (ds) cDNA was obtained through a
reaction using DNA ligase, DNA polymerase I and RNase H at 16°C for
2 h, followed by T4 DNA polymerase at 16°C for 5 min. After clean
up with a Sample Cleanup Module (Affymetrix, Inc.), ds-cDNA was
used for in vitro transcription (IVT). cDNA was transcribed
using the GeneChip IVT Labeling kit (Affymetrix, Inc.) in the
presence of biotin-labeled CTP and UTP. The resulting
biotin-labeled IVT-RNA was again purified with a Sample Cleanup
Module (Affymetrix, Inc.) and subsequently fragmented. Fragmented
cRNA was hybridized at 45°C for 16 h according to the
manufacturer’s instructions. Following hybridization, the arrays
were washed in a GeneChip Fluidics Station 450 with a non-stringent
wash buffer at 25°C and then by a stringent wash buffer at 50°C.
The arrays were then stained with a streptavidin-phycoerythrin
complex. After staining, the intensities were determined with a
GeneChip scanner. The duodenal mRNA expression profile obtained
from microarray data analyses for SLC15A1 was validated using
semiquantitative RT-PCR. The RT-PCR assay was performed as
described previously (6). The
pattern of SLC15A1 mRNA expression in the individual biopsies
determined by RT-PCR was similar to that observed in the microarray
data.
Data analysis
Official symbols and gene names were used in
accordance with the symbol and name lists approved by the Human
Genome Organization (HUGO) Gene Nomenclature Committee (http://www.genenames.org). Data analysis was performed
using GeneSpring 7.2 software (Silicon Genetics, Redwood City, CA,
USA). The numeric data were extracted from DAT images and
normalized using Microarray Suite software. Gene function analysis
was performed using the gene ontology-mining tool of NetAffx, which
is based on the Gene Ontology database (http://www.geneontology.org). GeneSpring also uses
data from public genomics databases to build gene ontologies based
on annotation information. For the present GeneChip probe array
study, the data for each gene represented data from 11–20 probe
pairs, each ∼25 bp in length. The overall target-specific intensity
was measured as the difference between the intensity of the
perfectly matched and mismatched probes. For normalization, data
from each expression array were scaled so that the overall
fluorescence intensity across each chip was equivalent (average
target intensity set at 500). The One-Sided Wilcoxon Signed Rank
test was employed to generate the detection P-value. If the overall
intensity of the perfect match was significantly larger than that
of the mismatch, the detection P-value was small. The probed gene
set was considered to be present if the P-value was <0.04. If
the P-value was >0.06, the probe set was considered to be
absent. The change algorithm generated a change P-value and an
associated fold-change value. The second algorithm gave a
quantitative estimate of the change in gene expression in the form
of a signal log ratio. The level of gene expression was considered
to be increased if its change P-value was <0.0025 and the gene
expression was considered to be decreased if its change P-value was
>0.9975.
Results
Sequence analysis
A total of 43,035 sequences from the dog duodenum
were analyzed and 60% exhibited >1-fold changes. As shown in
Table I, the total numbers of
detected transporter genes were 124 in the dog duodenum and 130 in
the mouse intestine. Among the transporter groups, the expression
levels of fatty acid, peptide, amino acid, glucose and multidrug
resistance/multidrug resistance-associated protein (MDR/MRP)
transporter genes were relatively higher than those of other
transporter gene groups in the two species.
 | Table INumbers of transporter genes expressed
in dog and mouse intestines. |
Table I
Numbers of transporter genes expressed
in dog and mouse intestines.
| Dog
| Mouse
|
|---|
| Transporter
cluster | Number | Expression level
(mean ± SD) | Number | Expression level
(mean ± SD) |
|---|
| Glucose
transporters | 18 | 8.1±12.8 | 11 | 19.5±37.0 |
| Amino acid
transporters | 7 | 12.6±31.9 | 15 | 7.1±12.7 |
| Peptide
transporters | 4 | 46.3±62.1 | 3 | 25.8±37.5 |
| Fatty acid
transporter | 12 | 75.0±167.8 | 13 | 41.9±85.0 |
| Mitochondrial solute
carrier | 10 | 16.6±50.5 | 16 | 23.8±42.3 |
| Nucleoside
transporters | 5 | 1.7±0. 8 | 6 | 3.6±3.4 |
| Nucleobase
transporter | 2 | 1.3±1.1 | 3 | 3.7±7.6 |
| MDRs/MRPs | 7 | 10.8±17.3 | 17 | 4.5±7.1 |
| Organic anion
transporters | 13 | 1.7±2.9 | 14 | 0.8±1.5 |
| Organic cation
transporters | 8 | 1.3±1.2 | 9 | 3.7±4.5 |
| Phosphate
transporters | 11 | 3.5±3.8 | 5 | 5.2±5.56 |
| Sodium/hydrogen
exchangers | 8 | 1.0±0.8 | 2 | 1.8±0. 5 |
| Sulfate
transporter | 2 | 6.1±7.3 | 2 | 1.6±1.1 |
| Zinc
Transporters | 17 | 2.9±4.2 | 14 | 6.7±5.2 |
| Total | 124 | – | 130 | – |
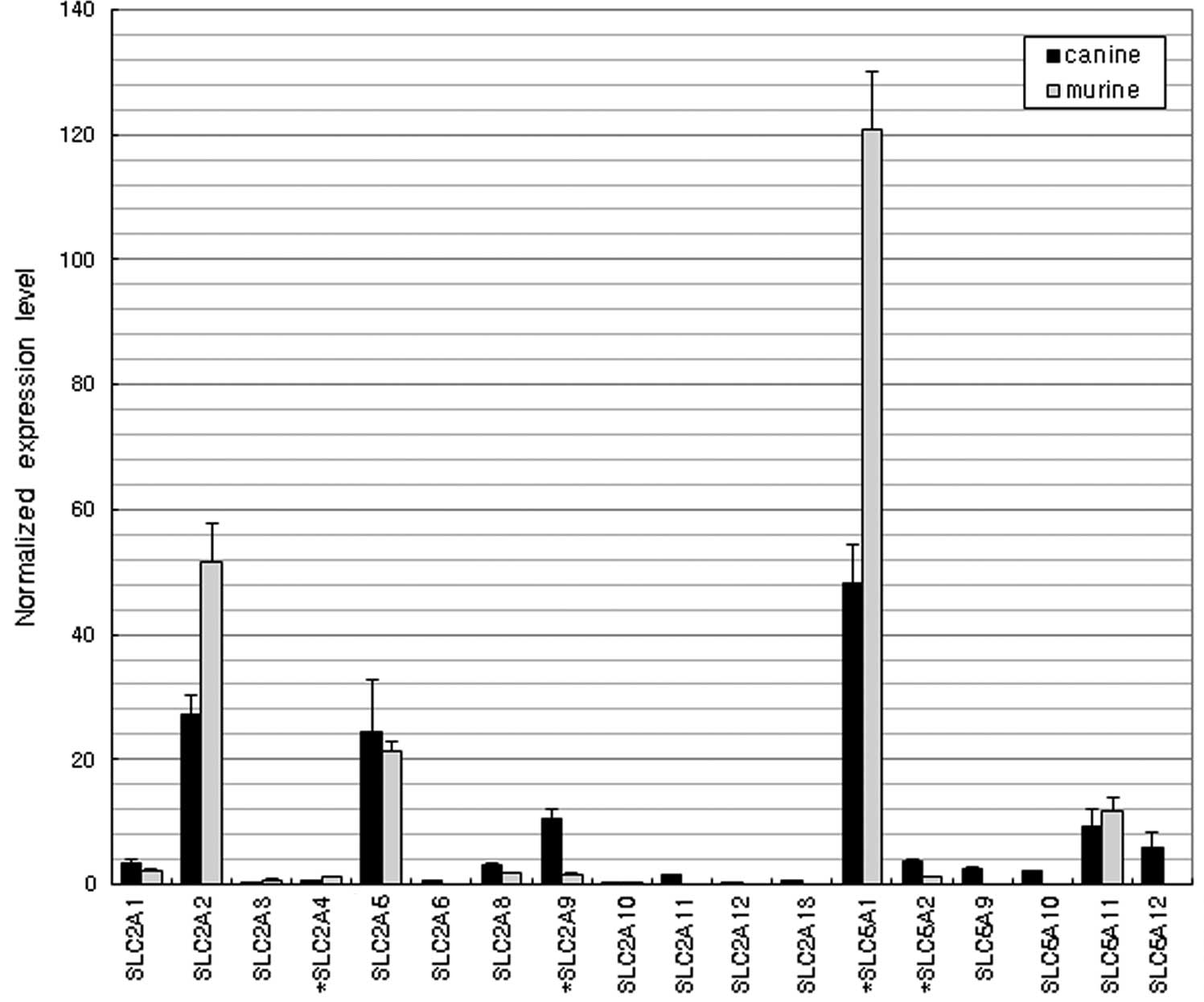
Glucose transporters
Fig. 1 shows the
similar expression profiles of the glucose transporter genes in the
dogs and mice. The majority of the SLC2 family (facilitated glucose
transporters, GLUTs) and SLC5 family (sodium/glucose
cotransporters, SGLTs) genes were expressed. SLC2A2, SLC2A5 and
SLC5A1 were the dominantly expressed genes in the two species.
Amino acid and peptide transporters
Fig. 2 shows the
expression levels of various amino acid and peptide transporter
genes in the duodenums of the dogs and mice. SLC3A1 and SLC15A1
were the dominantly expressed transporters in dogs, while SLC7A9
and SLC7A7 were highly expressed in mice. The expression of SLC15A1
was >15-fold higher in dogs than in mice (P<0.01).
Fatty acid transporters
The expression levels of fatty acid transporter
genes are shown in Fig. 3. The
overall expression levels of fatty acid transporter genes were
relatively high compared with those of the other transporter groups
(Table I). Among the fatty acid
binding proteins (FABPs), FABP1 and FABP2 were the most dominantly
expressed in both species.
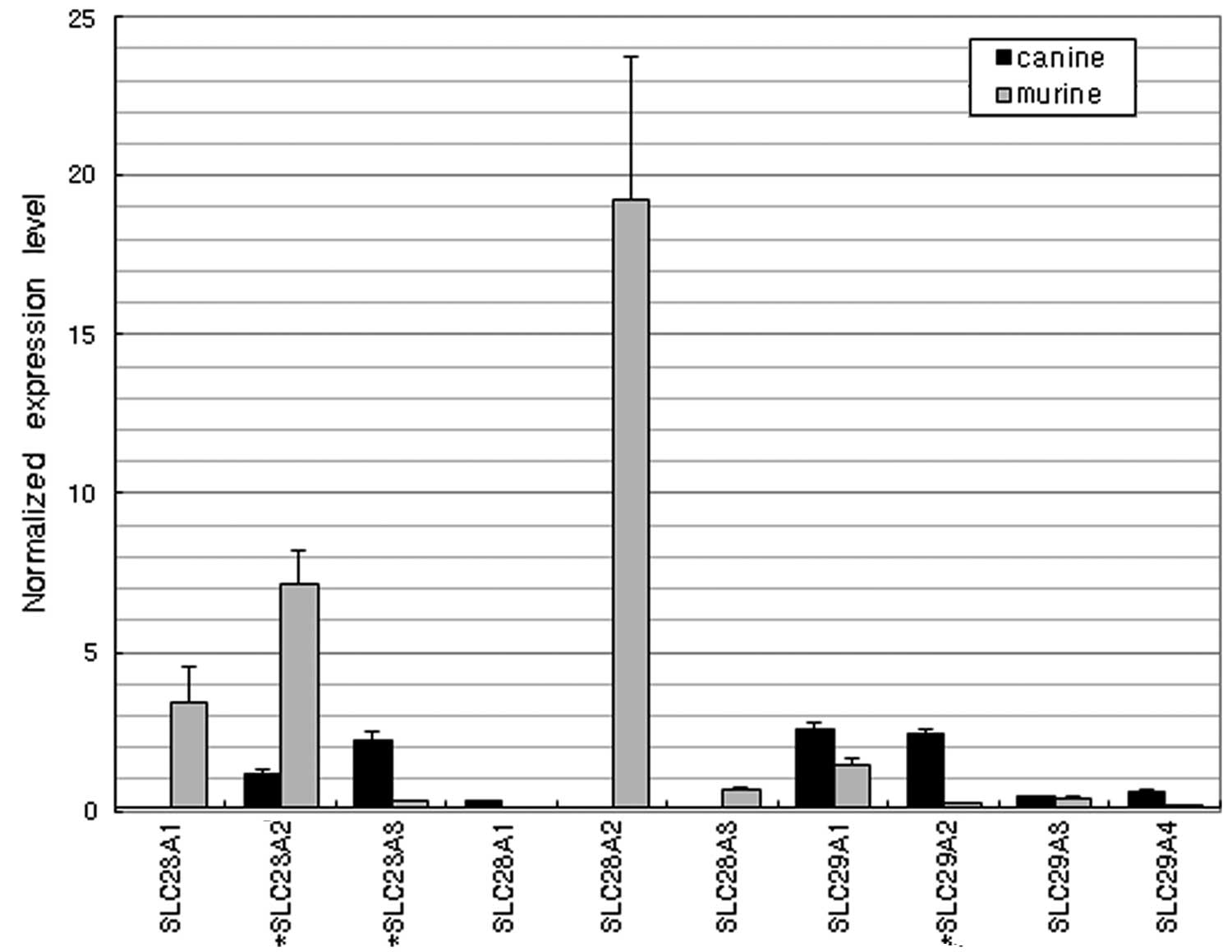
Nucleobase and nucleoside
transporters
As nucleobase transporters, SLC23A1 and SLC23A2 were
highly expressed in mice but poorly expressed in dogs (Fig. 4). Among the nucleoside transporter
genes, SLC28A2 (a sodium-coupled nucleoside transporter) was highly
expressed in mice but not in dogs. Another family of nucleoside
transporters, including SLC29A1 and SLC29A2, was more highly
expressed in dogs than in mice.
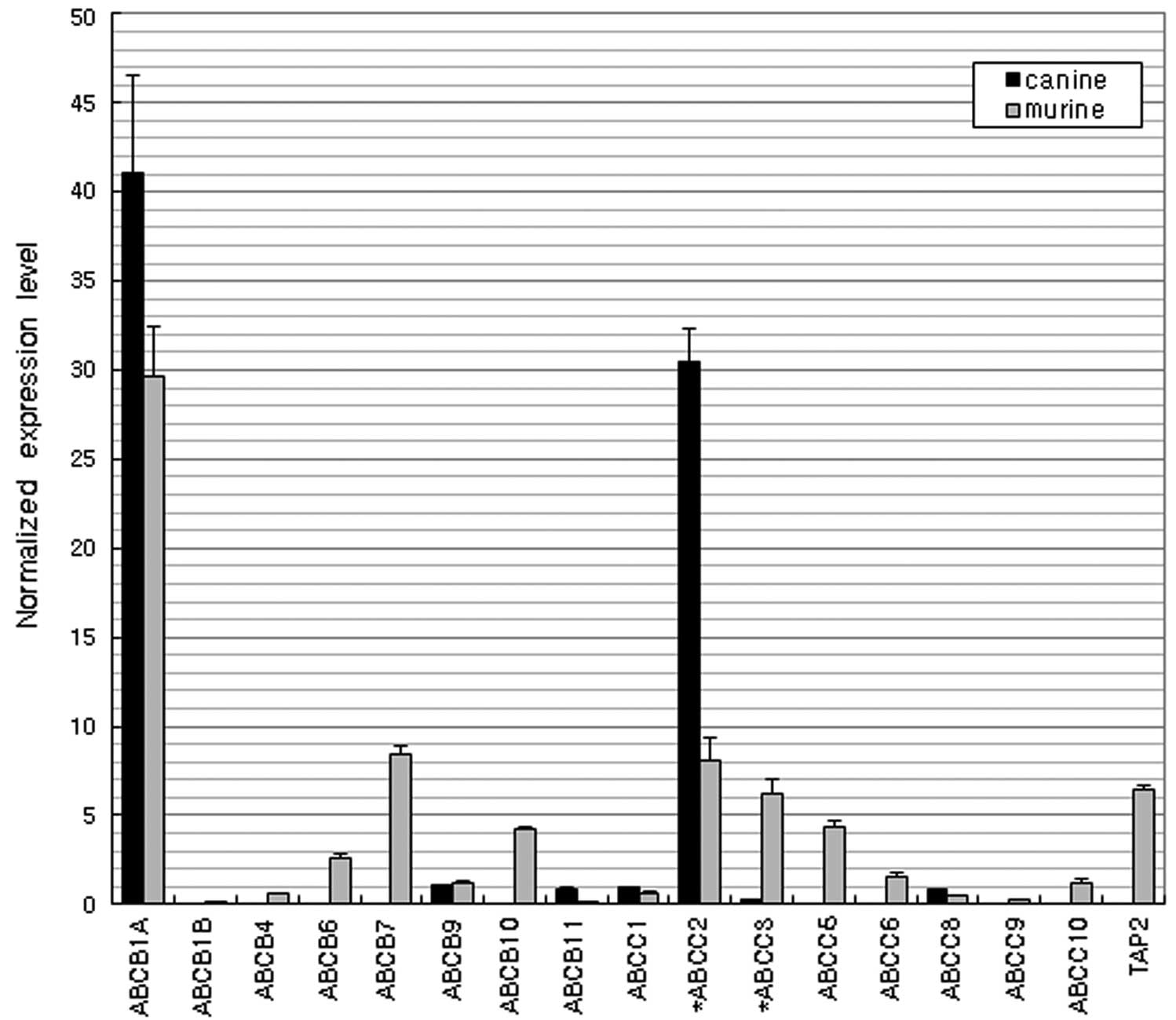
MDR and MRPs
Fig. 5 shows the
expression levels of ATP-binding cassette (ABC) genes. Similarly
high expression of ABCB1A (MDR/TAP) and ABCC2 (CFTR/MRP) were
observed in both species.
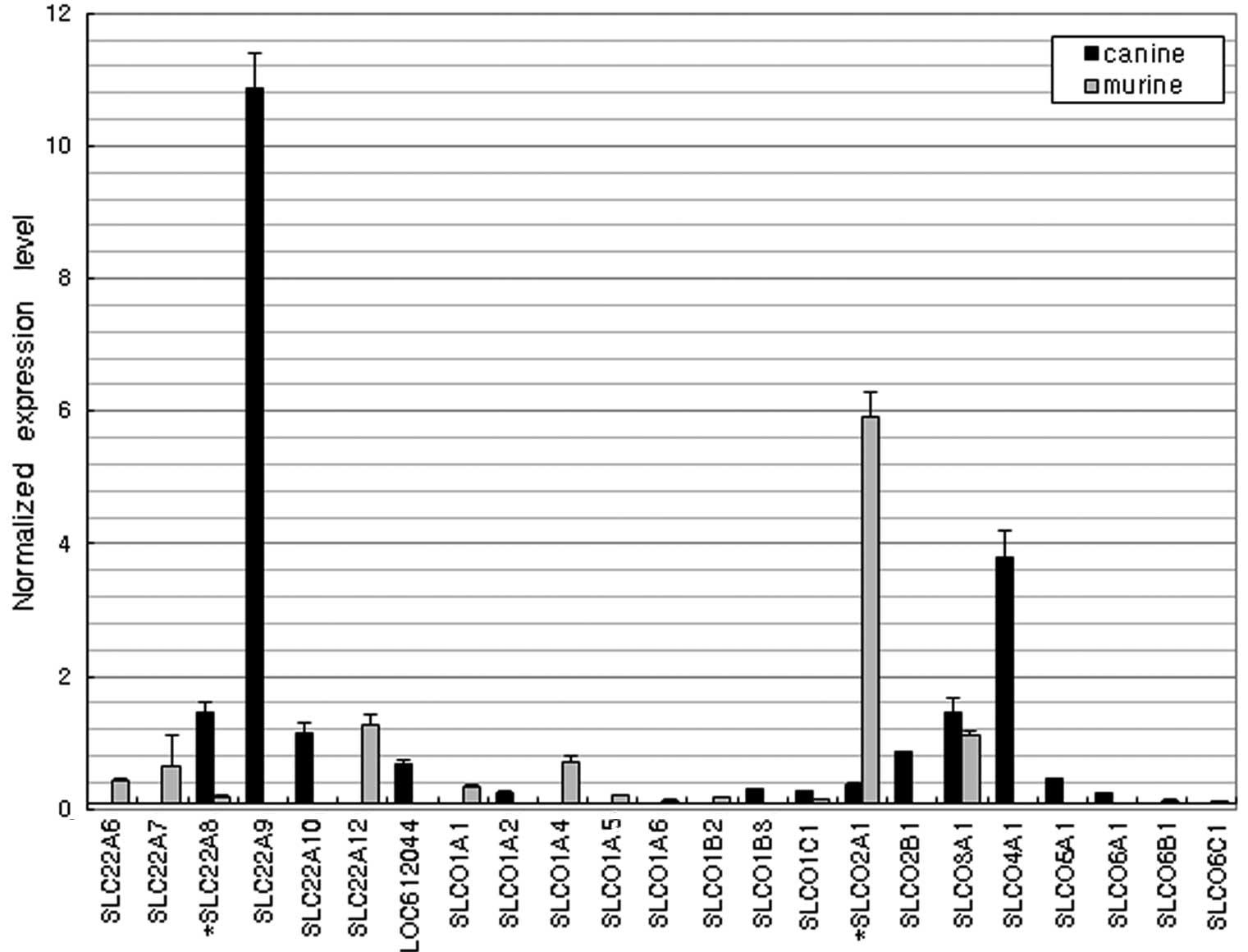
Organic anion and cation
transporters
Fig. 6 shows an
interspecies variation in the expression levels of organic anion
transporters. The most highly expressed anion transporter genes in
the dog duodenum were SLC22A9 and SLCO4A1 and the most strongly
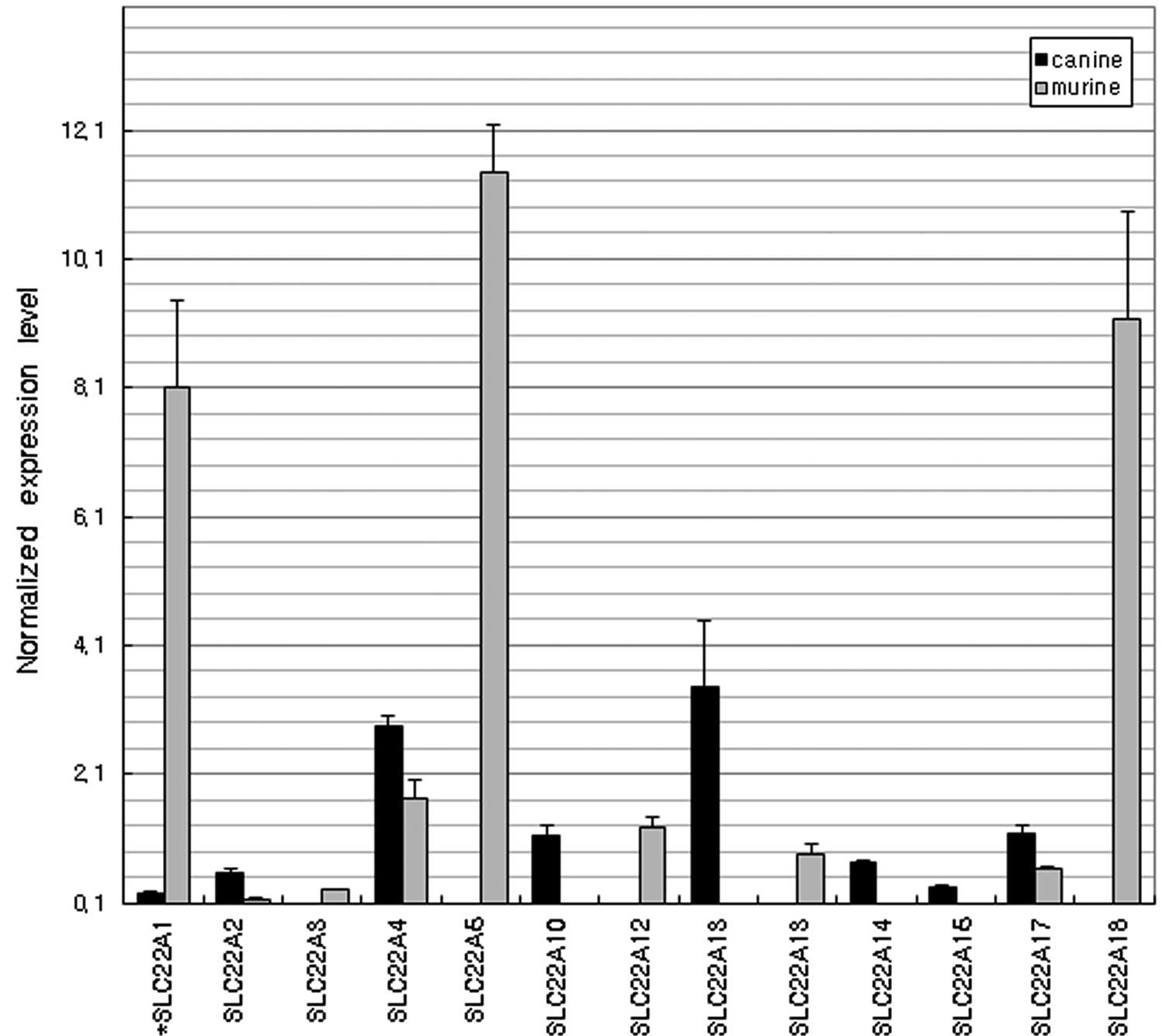
expressed gene in mice was SLCO2A1. Of the organic cation
transporters (Fig. 7), SLC22A1,
SLC22A5 and SLC22A18 were dominantly expressed in the mice although
their expression levels were negligible in the dogs. However, the
expression of SLC22A13 in dogs was >30-fold higher than that in
mice (P<0.01).
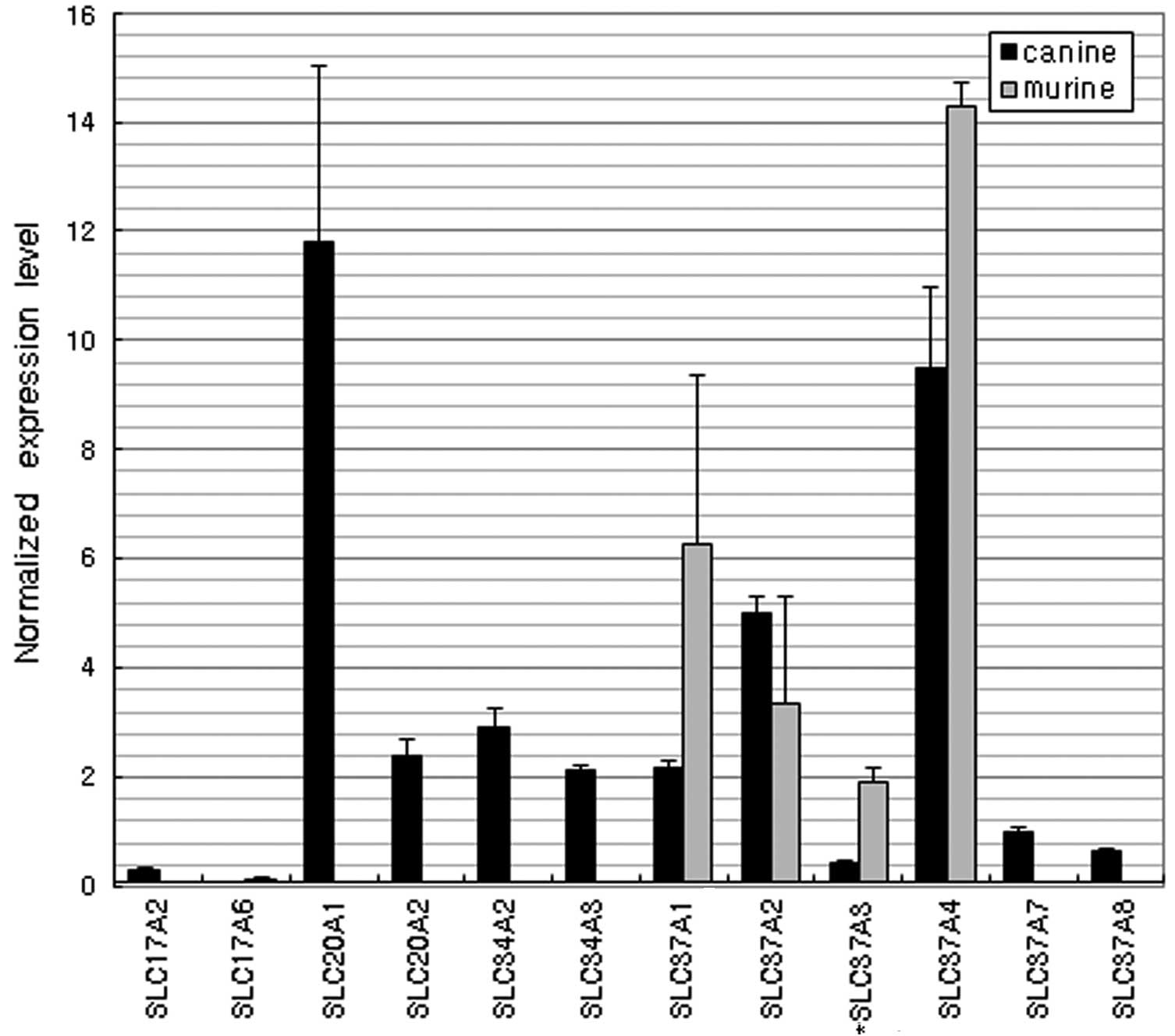
Phosphate transporters
Among the various phosphate transporters, the SLC17
(sodium phosphate transporter), SLC20 (phosphate transporter),
SLC34 (sodium phosphate transporter) and SLC37
(glycerol-3-phosphate transporter) family genes were detected in
the dogs (Fig. 8). Only 4 SLC37
family genes, SLC37A1, SLC37A2, SLC37A3 and SLC37A4, were expressed
in the mice. The most highly expressed genes were SLC20A1 in dogs
and SLC37A4 in mice (P<0.01).
Discussion
The completion of the DNA sequencing of the human,
mouse, dog and rat genomes and knowledge of cross-species gene
homologies enables the study of differential gene expression in
animal models (7). The
characterization of tissue-specific genes, such as intestinal
transporters, has the potential to greatly enhance the
understanding of the bioavailability of oral drugs (8–13).
It is now accepted that the process of drug absorption in the
intestine is highly associated with the functional gene expression
of intestinal transporters (8).
However, little is known about the transporter genes that are
transcribed in the dog intestine.
Membrane transport is a critical process for the
intestinal absorption of glucose, one of the primary energy sources
for various physiological and pharmacological functions (14). All glucose transporters have
significant homologies but they function differently to transport
various sugars or sugar-associated compounds from the intestine to
the systemic circulation with interspecies diversities (15). The data of the present study
revealed that the three glucose transporters SLC2A2 (GLUT2), SLC2A5
(GLUT5) and SLC5A1 (SGLT1) are similarly dominantly expressed in
dogs and mice. In humans, SLC5A1 transports glucose from the
intestinal lumen to the cytosol and SLC2A2 transports glucose from
the cytosol to the blood. Intestinal glucose absorption by the
apical SLC2A2 pathway may be 3- to 5-times greater than that by
SGLT1 at high concentrations of sugar (16). SLC2A5 preferentially transports
fructose rather than glucose (17).
The classification of amino acid transporters is
well defined according to their substrate specificity and tissue
distribution (18). The amino acid
transporters have highly restrictive substrate specificities
(19). The intestinal uptake of
certain anticancer drugs, such as the toxicants gabapentin,
pregabalin, melphalan, baclofen, D-cycloserine and methyl-mercury
L-cysteine complex, and other drugs is known to be mediated
specifically by these transporters. In the present study, a
significant difference was observed between the expression of the
transporters in the two species. SLC3A1 was dominantly expressed in
dogs and SLC7A9 and SLC7A7 (cationic amino acid transporters) were
detected at high levels in mice. Previous studies have demonstrated
that amino acid transporters are good targets for improving the
oral bioavailability of amino acid-associated drugs. The peptide
transporter SLC15A1 (PEPT1) was expressed in both species but its
expression level was 15-fold higher in dogs than in mice. The
proton-coupled peptide transporter is responsible for the
absorption of small peptides arising from the digestion of dietary
proteins (20). It is well known
that SLC15A1 is responsible for the uptake of a number of
peptide-like drugs, including β-lactam antibiotics, angiotensin
converting enzyme inhibitors, renin inhibitors, antitumor or
antiviral agents, thrombin inhibitors, a dopamine receptor
antagonist and amino acid prodrugs (21). Kim et al(22) also reported the notable expression
of SLC15A1 in the rat intestine.
Fatty acids are absorbed by passive diffusion.
However, numerous studies have demonstrated that various
transporters are involved in fatty acid absorption (23). Intestinal enterocytes contain high
concentrations of two cytosolic FABPs, FABP1 and FABP2, which are
hypothesized to be involved in cellular fatty acid trafficking. It
was suggested that FABP2, but not FABP1, may directly extract fatty
acids from membranes (24). The
present results reveal that the two FABP genes are the principal
fatty acid transporters expressed in the intestines of dogs and
mice with similar expression trends in both species. Schaffer and
Lodish (25) identified a membrane
protein which they termed fatty acid transport protein (FATP) from
mouse adipocytes. However, in the present study, the expression of
FATP was not particularly strong in either species.
Aside from the crucial roles of purines and
pyrimidines in DNA and RNA synthesis, they are also significant
components of a number of important biomolecules (26). Therefore, there is a specific
transport system associated with active transporters for delivering
nucleobases or nucleosides into cells. In the present study, a
considerable difference in the expression of nucleobase and
nucleoside transporters was observed between dogs and mice. In
mice, the nucleobase transporters SLC23A1 and SLC23A2 and
nucleoside transporter SLC28A2 (sodium-coupled nucleoside
transporters) were highly expressed but these genes were poorly
expressed in dogs. By contrast, the nucleoside transporters SLC29A1
and SLC29A2 were highly expressed in dogs. Nucleobase and
nucleoside analogs are widely used in the treatment of neoplasms
and viral infections (27,28).
Typical efflux transporters involve MRPs and MDRs,
which are members of the ABC superfamily. These proteins
translocate a wide variety of substrates, including sugars, amino
acids, nucleosides, lipids, bile salts, metal ions, peptides,
proteins and a large number of hydrophobic compounds and
metabolites across extra- and intracellular membranes (29,30).
In the present study, ABCB1A and ABCC2 (MRP2) were the most highly
expressed genes in the two species. Previous studies revealed that
ABCC2 accepts glutathione conjugates, sulfate conjugates,
glucuronides and non-conjugated organic anions, pravastatin,
vinblastine, temocaprilat, BQ-123, methotrexate, irinotecan and HIV
protease inhibitors, including saquinavir, ritonavir and indinavir.
ABCC3 (MRP3) mediates the transport of glucuronide conjugates,
taurocholate, glycocholate and methotrexate. Differential
expression of ABCC3 was observed between the two species but the
expression levels of ABCC3 were lower than those of ABCC2.
Organic anion transporters serve as the efflux
system for a number of endogenous compounds, anionic drugs,
environmental substances and their metabolic products, which are
usually harmful to the body. In particular, these families are
known to affect the pharmacokinetics and drug-drug interactions of
various drugs (31). In the
present study, SLC22A9 and SLCO4A1 in dogs and SLCO2A1 in mice were
dominantly expressed and the overall levels of expression were
higher in dogs than in mice. SLCO4A1 mediates the
sodium-independent transport of organic anions, including the
thyroid hormones triiodo-L-thyronine and thyroxine. Organic cation
transporters in the liver, kidney and intestine are critical for
the absorption and elimination of a number of endogenous amines as
well as a wide range of drugs and environmental toxins. In the
present study, the SLC22 family gene expression profile was
markedly different between the dogs and mice. The expression of
SLC22A13 was more than 30-fold higher in dogs than in mice.
SLC22A13 was originally known as an organic cation transporter but
Bahn et al(32) renamed it
OAT10 (organic anion transporter) since it is able to transport
various anionic compounds, including nicotinate, p-aminohippurate
and urate.
In the analysis of phosphate transporters, the data
reveal that members of the SLC17, SLC20, SLC34 and SLC37 families
were expressed in the dogs, while only SLC37 family genes,
including SLC37A1, SLC37A2, SLC37A3 and SLC37A4, were expressed in
the mice. The most highly expressed genes in the dogs and mice were
SLC20A1 and SLC37A4, respectively. SLC20A1 is a ubiquitously
expressed sodium-phosphate symporter that plays a fundamental
housekeeping role in the maintenance of phosphate homeostasis.
SLC20A1 may also function as a retroviral receptor as it confers
susceptibility to certain viral infections in human cells.
Although similar profiles were observed for several
transporters in both species, including the glucose transporters
SLC5A1 and SLC2As, the fatty acid transporters FABP1 and FABP2, the
efflux transporters ABCB1A and ABCC2 and the phosphate transporter
SLC37A4, overall, the data of the present study reveal markedly
different transcriptomic profiles for the intestinal transporters
of dogs and mice. The dog is an extremely important animal species,
not only as a laboratory animal, but also as a major animal in
veterinary medicine. However, genetic information has not been
explored extensively for the dog in comparison with other model
animals. Therefore, the database generated in the present study may
be useful for comparing the intestinal xenobiotic transport systems
of dogs and other animals.
Acknowledgements
This study was supported by a grant
(10162KFDA995) from the Korea Food & Drug Administration in
2012.
References
|
1
|
Varma MV: Role of intestinal transporters
and metabolism in the oral absorption of drug and prodrugs. Curr
Drug Metab. 11:7152010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katsura T and Inui K: Intestinal
absorption of drugs mediated by drug transporters: mechanisms and
regulation. Drug Metab Pharmacokinet. 18:1–15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sugiura T, Kato Y and Tsuji A: Role of SLC
xenobiotic transporters and their regulatory mechanisms PDZ
proteins in drug delivery and disposition. J Control Release.
116:238–246. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rankin KS, Starkey M, Lunec J, et al: Of
dogs and men: comparative biology as a tool for the discovery of
novel biomarkers and drug development targets in osteosarcoma.
Pediatr Blood Cancer. 58:327–333. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang W, Jiang J, Ballard CE and Wang B:
Prodrug approaches to the improved delivery of peptide drugs. Curr
Pharm Des. 5:265–287. 1999.PubMed/NCBI
|
|
6
|
Shin HC, Kim HR, Cho HJ, et al:
Comparative gene expression of intestinal metabolizing enzymes.
Biopharm Drug Dispos. 30:411–421. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang H, Tong W, Perkins R, et al:
Bioinformatics approaches for cross-species liver cancer analysis
based on microarray gene expression profiling. BMC Bioinformatics.
6(Suppl 2): S62005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Landowski CP, Sun D, Foster DR, et al:
Gene expression in the human intestine and correlation with oral
valacyclovir pharmacokinetic parameters. J Pharmacol Exp Ther.
306:778–786. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takara K, Ohnishi N, Horibe S and Yokayama
T: Expression profiles of drug-metabolizing enzyme CYP3A and drug
efflux transporter multidrug resistance 1 subfamily mRNAS in small
intestine. Drug Metab Dispos. 31:1235–1239. 2003. View Article : Google Scholar
|
|
10
|
Rushmore TH and Kong AN: Pharmacogenomics,
regulation and signaling pathways of phase I and II drug
metabolizing enzymes. Curr Drug Metab. 3:481–490. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mizuno N, Niwa T, Yotsumoto Y and Sugiyama
Y: Impact of drug transporter studies on drug discovery and
development. Pharmacol Rev. 55:425–461. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Beaumont K: The importance of gut wall
metabolism in determining drug bioavailability. Drug
Bioavailability/Estimation of Solubility, Permeability and
Absorption. van de Waterbeemd H, Lennernas H and Artursson P:
Wiley-VCH; Weinheim: pp. 311–328. 2003
|
|
13
|
Tsuji A: Transporter-mediated drug
interactions. Drug Metab Pharmacokinet. 17:253–274. 2002.
View Article : Google Scholar
|
|
14
|
Mizuma T, Ohta K and Awazu S: The
beta-anomeric and glucose preferences of glucose transport carrier
for intestinal active absorption of monosaccharide conjugates.
Biochim Biophys Acta. 1200:117–122. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao FQ and Keating AF: Expression and
regulation of glucose transporters in the bovine mammary gland. J
Dairy Sci. 90(Suppl 1): E76–E86. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stelmańskan E: The important role of GLUT2
in intestinal sugar transport and absorption. Postepy Biochem.
55:385–387. 2009.(In Polish).
|
|
17
|
Phay JE, Hussain HB and Moley JF: Cloning
and expression analysis of a novel member of the facilitative
glucose transporter family, SLC2A9 (GLUT9). Genomics. 66:217–220.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tate BJ, Witort E, McKenzie IF and Hogarth
PM: Expression of the high responder/non-responder human Fc gamma
RII. Analysis by PCR and transfection into FcR-COS cells. Immunol
Cell Biol. 70(Pt 2): 79–87. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wells RG and Hediger MA: Cloning of a rat
kidney cDNA that stimulates dibasic and neutral amino acid
transport and has sequence similarity to glucosidases. Proc Natl
Acad Sci USA. 89:5596–5600. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seal CJ and Parker DS: Isolation and
characterization of circulating low molecular weight peptides in
steer, sheep and rat portal and peripheral blood. Comp Biochem
Physiol B. 99:679–685. 1991.PubMed/NCBI
|
|
21
|
Lee VH, Chu C, Mahlin ED, et al:
Biopharmaceutics of transmucosal peptide and protein drug
administration: role of transport mechanisms with a focus on the
involvement of PepT1. J Control Release. 62:129–140. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim HR, Park SW, Cho HJ, et al:
Comparative gene expression profiles of intestinal transporters in
mice, rats and humans. Pharmacol Res. 56:224–236. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abumrad N, Harmon C and Ibrahimi A:
Membrane transport of long-chain fatty acids: evidence for a
facilitated process. J Lipid Res. 39:2309–2318. 1998.PubMed/NCBI
|
|
24
|
Thumser AE and Storch J: Liver and
intestinal fatty acid-binding proteins obtain fatty acids from
phospholipid membranes by different mechanisms. J Lipid Res.
41:647–656. 2000.PubMed/NCBI
|
|
25
|
Schaffer JE and Lodish HF: Expression
cloning and characterization of a novel adipocyte long chain fatty
acid transport protein. Cell. 79:427–436. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Traut TW: Physiological concentrations of
purines and pyrimidines. Mol Cell Biochem. 140:1–22. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
el Kouni MH: Trends in the design of
nucleoside analogues as anti-HIV drugs. Curr Pharm Des. 8:581–593.
2002.PubMed/NCBI
|
|
28
|
Damaraju VL, Damaraju S, Young JD, et al:
Nucleoside anti-cancer drugs: the role of nucleoside transporters
in resistance to cancer chemotherapy. Oncogene. 22:7524–7536. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sandusky GE, Mintze KS, Pratt SE and
Dantzig AH: Expression of multidrug resistance-associated protein 2
(MRP2) in normal human tissues and carcinomas using tissue
microarrays. Histopathology. 41:65–74. 2002. View Article : Google Scholar
|
|
30
|
Tang F and Borchardt RT: Characterization
of the efflux transporter(s) responsible for restricting intestinal
mucosa permeation of an acyloxyalkoxy-based cyclic prodrug of the
opioid peptide DADLE. Pharm Res. 19:780–786. 2002. View Article : Google Scholar
|
|
31
|
Miyazaki H, Sekine T and Endou H: The
multispecific organic anion transporter family: properties and
pharmacological significance. Trends Pharmacol Sci. 25:654–662.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bahn A, Hagos Y, Reuter S, et al:
Identification of a new urate and high affinity nicotinate
transporter, hOAT10 (SLC22A13). J Biol Chem. 283:16332–16341. 2008.
View Article : Google Scholar : PubMed/NCBI
|