Introduction
Peritoneal dialysis (PD) is the main treatment for
patients with end-stage renal disease. However, peritoneal fibrosis
(PF) caused by long-term PD eventually leads to ultrafiltration
failure, which limits the application of PD (1,2).
Human peritoneal mesothelial cells (HPMCs) constitute the largest
cell population in the peritoneum. They maintain the peritoneal
structure and stable function, preventing abdominal infection and
peritoneal sclerosis. The epithelial-mesenchymal transition (EMT)
is an important cause of PF in the PD process (3,4). The
loss of epithelial cell adhesion and phenotypic transformation of
fibroblasts to muscle are key steps in EMT (5).
Traditional HPMC isolation and culture is difficult
to perform. In this study, the HPMCs shed from the peritoneal
effluent of PD patients were separated and cultured. The HPMCs of
PD patients reflect the function of peritoneal cells and may be
analyzed for changes in the gene expression of cellular proteins
E-cadherin, vimentin, fibronectin (FN) and COL-1. This is an
effective method for investigating the physiological functions and
changes in HPMCs in PD patients, and provides clinical evidence for
reversing or preventing ultrafiltration failure.
A microRNA (miRNA) is a functional non-coding small
molecule RNA existing in plant and animal genomes, with 21–25
nucleotides. miRNA binds with the untranslated region of the 3′
end, the coding region or the untranslated region of 5′ end of the
target mRNA. This inhibits translation or triggers the degradation
of mRNA and therefore inhibits translation following gene
transcription. It has been demonstrated that miRNA directly
regulates 30% of the transcription of protein-coding genes in the
mammalian genome. miRNA plays an important role in cells and is
involved in almost all biological processes (6,7).
Previously, it has been shown that target miRNA may effectively
reverse EMT and inhibit fibrosis of organs, including the heart,
liver, lung and kidney. (8–11).
It has also been shown that miRNA is a therapeutic target and a
diagnostic marker (12–14). However, the role of miRNA in
peritoneal mesothelial cell EMT and PF is unclear.
MicroRNA-200 (miRNA-200) is a recently discovered
family that is closely associated with a variety of fibrotic
diseases, and includes miR-200a, miR-200b, miR-200c, miR-141 and
miR-429. The abnormal expression of the miRNA-200 gene cluster or a
single gene has been observed in a variety of fibrotic diseases,
including liver fibrosis, pulmonary fibrosis, kidney fibrosis and
systemic sclerosis (15–17). Regulation of its expression may
inhibit or reduce fibrosis, suggesting that miRNA-200 may be a
potential anti-fibrotic target. In the present study, the changes
in miR-200c expression levels in the HPMCs of PD patients were
studied.
Materials and methods
Patients
The study included 28 patients with continuous
ambulatory peritoneal dialysis (CAPD) without peritonitis,
including 13 males and 15 females, aging from 41 to 73 (54.37±9.33)
years old. There were 12 patients with newly inserted tubes
(dialysis time ≤15 days; PD start group) and 16 patients who had
been undergoing PD for >6 months (the PD >6 months group)
with a dialysis time of 6–22 (10.80±4.38) months and a dialysate
concentration of 2.5% (Baxter, Deerfield, IL, USA). There were 13
patients with chronic glomerulonephritis, 8 patients with diabetic
nephropathy and 7 patients with hypertensive nephropathy. Prior
written and informed consent were obtained from every patient and
the study was approved by the ethics review board of Central South
University (Changsha, China).
Reagents
Fetal calf serum (15% FCS; Hangzhou Evergreen
Biotechnology Co., Hangzhou, China) was added to DMEM/F12 medium
(Gibco, Carlsbad, CA, USA). Goat E-cadherin antibody, mouse
vimentin antibody, rat anti-human factor VIII antibody, leukocyte
CD45, rabbit FN antibody, rabbit COL-1 antibody, mouse GAPDH
antibody and goat anti-mouse horseradish peroxidase (HRP) labeled
secondary antibody was purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). TRIzol reagent was purchased from
Invitrogen (Carlsbad, CA, USA). The Revert Aid™ First strand cDNA
Synthesis kit was purchased from Fermentus (Vilnius, Lithuania).
SYBR GreenER™ qPCR SuperMix, the miRNeasy Mini kit and miRNA Q-PCR
Detection kit were purchased from Invitrogen (USA).
Isolation and culture of HPMCs
Samples of sterile abdominal effluent (~2,000 ml)
were collected from the CAPD patients at night. The samples were
centrifuged at 500 × g and 4°C for 5 min and the supernatant was
discarded. The precipitated cells were washed twice with D-Hank’s
balanced salt solution. Cells resuspended in 15% FCS DMEM/F12
medium were added to adjust the cell number to 1×106
cells/ml and inoculated on gelatin-coated 25 mm2
(Corning, New York, NY, USA). The cells were cultured at 37°C in a
5% carbon dioxide incubator and the culture medium was changed
every 72 h. The cultured cells were detected under an inverted
phase contrast microscope (Eclipse TS100-F; Nikon, Tokyo, Japan)
and by immunofluorescence and scanning electron microscopy (Philips
CM 120 transmission electron microscope, Amsterdam, The
Netherlands).
Immunofluorescence
The cells were washed with PBS three times and fixed
with 2–4% formaldehyde for 15 min. Non-immune goat serum containing
0.3% Triton X-100 was added and the cells were kept at room
temperature for 60 min. The primary antibody was added at 4°C
overnight. The primary antibodies used were anti-E-cadherin
antibody, anti-Vimentin, anti-A cyclase VIII Antibody (B-6) and
anti-CD45 antibody. The cells were washed three times with PBS for
5 min. The secondary antibody (50 μl; 1:200) was added and
the cells were kept at room temperature for 60 min. The secondary
antibodies were Alexa Fluor® 488 goat anti-mouse IgG2b
(H+L) and Alexa Fluor® 488 mouse anti-goat IgG (H+L).
Finally, the cells were stained with DAPI. After washing three
times with PBS, the cells were observed under a fluorescence
microscope.
Electron microscopy
The cells were washed three times with cold PBS and
fixed with 2.5% glutaraldehyde for 30 min. After washing three
times with PBS for 5 min, the cells were fixed with 1% osmium
tetroxide for 4 h at 4°C. The cells were dehydrated with ethanol.
After drying and gilding by ion sputter, the cells were detected by
scanning electron microscopy.
qPCR
Total cell RNA was extracted with TRIzol reagent.
The cDNA synthesis was performed according to the manufacturer’s
instructions for the RevertAid™ H Minus FirstStrand cDNA Synthesis
kit. The PCR amplification was performed using the SYBR GreenER
qPCR SuperMix according to the manufacturer’s instructions.
Deionized water was the negative control for each PCR. The relative
expression amount of target gene mRNA=2−ΔΔCt. The Ct
value was the cycle number before fluorescence reached the
threshold value. ΔΔCt=(Ct target gene - Ct housekeeper gene)
experimental group - (Ct target gene - Ct housekeeper gene) control
group. In this experiment, the PD start group was considered the
control group. The relative expression level of mRNA in the more
than 6 months PD group was expressed as 2-ΔΔCt in the PD start
group. The primers used in this study are shown in Table I.
 | Table I.The primers used in this study. |
Table I.
The primers used in this study.
| Primers | Sequences | Product size
(bp) |
|---|
| E-cadherin
forward |
5′-TCATGAGTGTCCCCCGGTAT | 240 |
| E-cadherin
reverse |
5′-TCTTGAAGCGATTGCCCCAT | |
| Vimentin forward |
5′-GCTACGTGACTACGTCCACC | 265 |
| Vimentin reverse |
5′-TAGTTGGCGAAGCGGTCATT | |
| FN forward |
5′-AACTGGTAACCCTTCCACACCC | 266 |
| FN reverse |
5′-AGCTTCTTGTCCTACATTCGGC | |
| COL-1 forward |
5′-GCCAAGACGAAGACATCCCA | 156 |
| COL-1 reverse |
5′-GGCAGTTCTTGGTCTCGTCA | |
| GAPDH forward |
5′-CAATGACCCCTTCATTGACC | 106 |
| GAPDH reverse |
5′-GACAAGCTTCCCGTTCTCAG | |
| hsa-miR-200c
forward |
5′-TAATACTGCCGGGTAATGATGGA | 75 |
| hsa-miR-200c
reverse |
5′-TGGTGTCGTGGAGTCG | |
| U6 forward |
5′-GCTTCGGCAGCACATATACTAAAAT | 81 |
| U6 reverse |
5′-CGCTTCACGAATTTGCGTGTCAT | |
Western blotting
Total protein was extracted for 12% SDS-PAGE
electrophoresis. The primary antibodies against E-cadherin (1:500),
vimentin (1:800), FN (1:500) and COL-1 (1:500) were used and
incubated at room temperature for 1 h. Secondary antibodies used in
this study were goat anti-mouse IgG-HRP (Cat# sc-2005, 1:10,000;
Santa Cruz Biotechnology, Inc.). After development by enhanced
chemiluminescence (ECL), gel pro 4.0 software (Media Cybernetics,
Rockville, MD, USA) was used for image analysis.
Statistical analysis
All data were analyzed by SPSS 11.0 statistical
software (SPSS, Inc., Chicago, IL, USA). The experimental data are
expressed as mean ± standard deviation. A Student’s t-test was used
to compare values. P<0.05 was considered to indicate a
statistically significant result.
Results
Morphological changes of HPMCs in
patients
To determine the morphological changes in the HPMCs
of patients, the cells were detected using microscopy,
immunofluorescence labeling, electron microscopy and transmission
electron microscopy. The results of the PD start group is shown in
Fig. 1A–G and the results of the
PD >6 months is shown in Fig.
H. Effluent HPMCs were fusiform after 2 days of primary culture
and were not easily differentiated from fibroblasts (Fig. 1A). At 14 days, cells were fused
(Fig. 1B). In order to determine
the sources of the HPMCs, immunofluorescence detection was
performed. This showed that cytoplasmic vimentin (Fig. 1C) and cytokeratin (Fig. 1D) antigens were positively detected
in the cells. A visible green fluorescent area in the cytoplasm was
considered a positive result. The cell nucleus was restained with
DAPI in order to show the complete nucleus (Fig. 1C and D). There was no fluorescent
signal for factor VIII and leukocyte CD45, suggesting that there
was no factor VIII and leukocyte CD45 expression in the cell and
the cell purity was >95%. Scanning electron microscopy showed
filamentous microvilli of different lengths on the cell surface
(Fig. 1E). Transmission electron
microscopy showed a large number of microvilli on the cell surface
and abundant endoplasmic reticulum and mitochondria in the
cytoplasm without Weibel-Palade bodies (Fig. 1F).
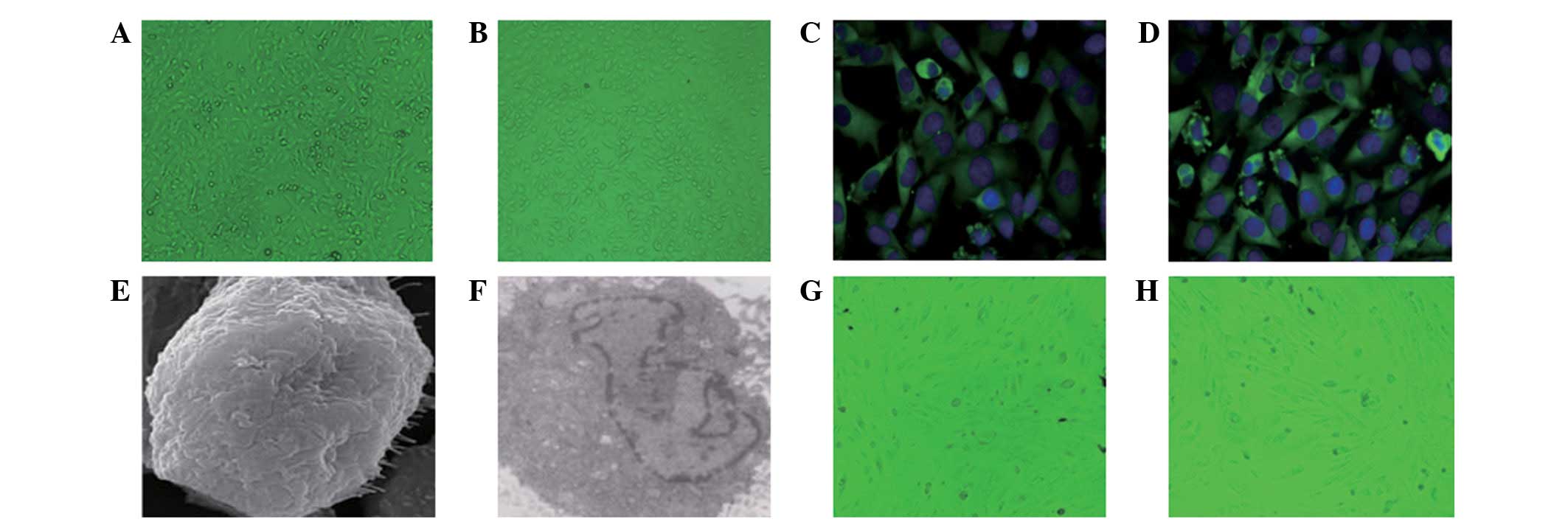 | Figure 1.Observation of HPMCs. Observation of
HPMCs under a microscope at (A) day 2 after primary culture
(magnification, ×200; observed under light microscope, no staining)
and (B) day 14 after primary culture (magnification, ×200; observed
under light microscope, no staining). Immunofluorescence showed (C)
vimentin-positive HPMCs (magnification, ×400; immunofluorescence
staining) and (D) cytokeratin-positive HPMCs (magnification, ×400;
immunofluorescence staining). (E) Electron microscopy showed an
HPMC in PD effluent culture (magnification, ×10,000; gilded with
ion sputter). (F) Transmission electron microscopy of an HPMC in PD
effluent culture (magnification, ×4,000; gilded with ion sputter).
(G) Mixed mesothelium (magnification, ×200; observed under light
microscope, no staining). (H) Fibroblast-like mesothelium
(magnification, ×200; observed under light microscope, no
staining). (A–G) HPMCs were from the PD start group; (H) HPMCs were
from the PD >6 months group. HPMCs, human peritoneal mesothelial
cells; PD, peritoneal dialysis. |
Through a light microscope, the cells in the PD
effluent were observed to be round or oval epithelioid cells,
spindle fiber-like cells or mixed HPMCs. Certain HPMCs of the PD
start group were round or oval-shaped epithelioid cells. After
fusion, cells were cobblestone-like, similar to omentum-cultured
HPMCs. However, the majority of epithelioid cells and spindle cells
were mixed (Fig. 1G). Spindle
fiber-like cells were mainly observed in patients who had been
undergoing PD for >6 months (Fig.
1H). The cell morphological changes indicate that the cells
from the peritoneal dialysate effluent fluids are HPMCs, which
suggests the occurrence of EMT.
E-cadherin, vimentin, FN and COL-1
expression of peritoneal mesothelial cells in PD patients
To determine the expression levels of proteins,
HPMCs were harvested from the patients in the PD start group and
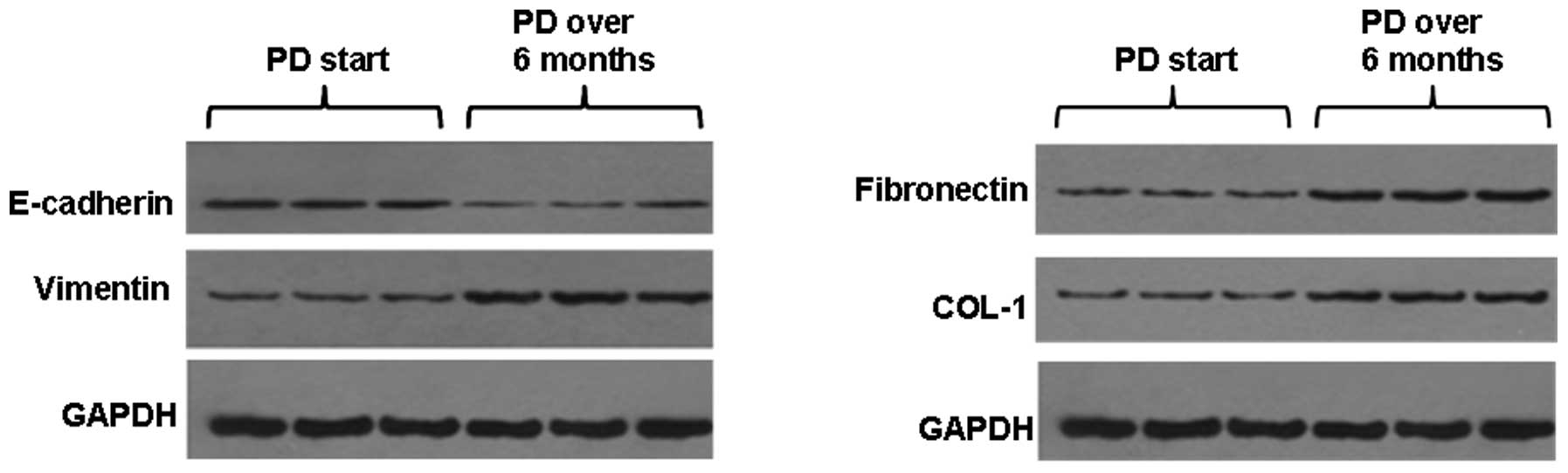
the PD >6 months group. The western blot analysis showed that
E-cadherin protein expression levels in the peritoneal mesothelial
cells were significantly reduced in the PD >6 months group
(P<0.05) and the vimentin, FN and COL-1 protein expression
levels were significantly increased (P<0.05) compared with those
in patients in the PD start group (Fig. 2).
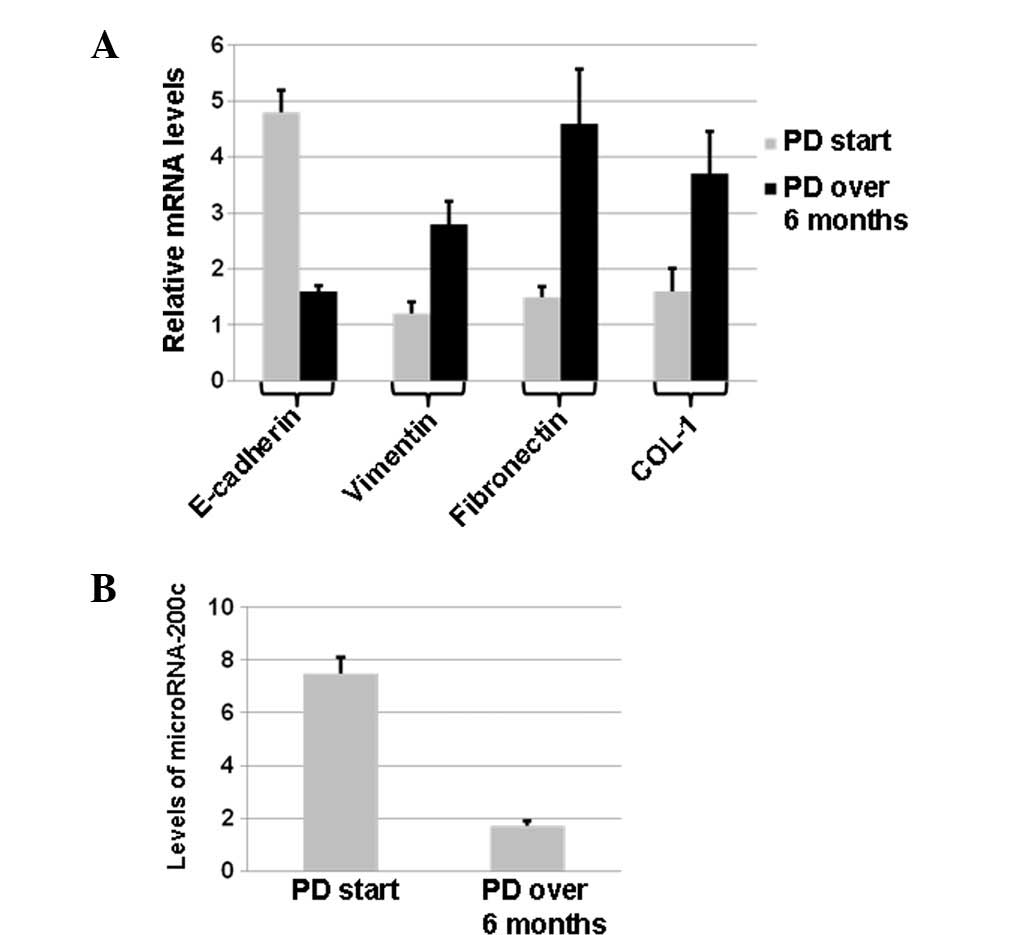
qPCR also showed that E-cadherin mRNA levels in the
peritoneal mesothelial cells were significantly decreased in the PD
>6 months group (P<0.05), and vimentin, COL-1 and FN mRNA
levels were significantly increased (P<0.05) (Fig. 3A). These results suggest that the
expression of these molecular marker proteins was altered in
different ways at the transcriptional levels.
miRNA-200c expression changes in the
effluent peritoneal mesothelial cells of PD patients
To determine the changes in miRNA-200c expression,
qPCR was performed. With U6 as a reference gene, miRNA-200c
expression in the peritoneal mesothelial cells of PD patients was
determined by qPCR Taqman probe assay. The results showed that the
relative expression of miRNA-200C was 7.546±0.341 in the PD start
group and 1.701±1.070 in the PD >6 months group; the expression
level was significantly lower in the PD >6 months group than in
the PD start group (P<0.05; Fig.
3B). These results suggest that changes in miRNA-200c levels
may be associated with EMT.
Discussion
EMT is an important source of formation of
myofibroblasts (18,19). In 2003, morphological changes in
the HPMCs of PD effluents were first reported, including in the
epithelial cells and the fiber cells (20). In the present study, HPMCs were
separated and cultured from the PD effluent with a cell purity of
>95%. At one and two days after primary culture, the effluent
HPMCs were fusiform and could not be differentiated from
fibroblasts until 10-4 days. Immunofluorescence and scanning
electron microscopy analysis of the effluent HPMCs was positive for
vimentin, cytokeratin and microvilli on the cell surface. By
contrast, myofibroblasts were positive for vimentin and negative
for cytokeratin on the cell surface. This indicates that the
myofibroblasts fiber-like cells were derived from mesothelial
cells, which indicated the occurrence of EMT.
Previous in vivo experiments have indicated
that are were fiber-like cells in the peritoneal tissues of
long-term PD patients, which not only express typical epithelial
cell markers but also expressed fibroblast markers, such as α-SMA
(21). It has also been
demonstrated that EMT-related changes in peritoneal mesothelial
cells may present in early peritoneal dialysis (20). As the time of PD is extended,
epithelial cells gradually change to a shuttle type and the
expression of epithelial cell marker E-cadherin decreases, while
the expression of mesenchymal marker vimentin increases (22,23).
Previously it was shown that in PD patients with 2 years of
dialysis, 74% lost mesothelial cells in the peritoneum, 46% had PF
and 17% showed evidence of in situ EMT in the peritoneum (24). There were myofibroblasts in all
peritonea with EMT changes. In this experiment, it was observed
that in the PD start group, the mesothelial cells in the peritoneal
dialysis effluent were round, oval and mixed epithelioid cells. For
the mixed cells, EMT of the peritoneal mesothelial cells was
detected in early peritoneal dialysis. However, as the dialysis
time was extended, the cell morphology changed significantly, and
the cells in the PD more than 6 months group were long spindle
fiber-like. We observed that the expression level of E-cadherin was
decreased, whereas the expression of vimentin, Col-1 and FN was
significantly increased in the PD >6 months group, suggesting
that the protein expression was changed significantly. In the early
stage of CAPD, when cells remain cubic, the expression levels of
E-cadherin, vimentin, FN and COL-1 changed, indicating EMT is the
starting point of PF. When the peritoneum is exposed to a
mechanical exfoliation environment for a long time, the phenotype
inevitably becomes abnormal and mesothelial cells are completely
transformed due to injury. This promotes fibrosis and
ultrafiltration failure. These molecular protein markers provide
evidence for further study of the molecular mechanisms of PF. We
also observed that the level of microRNA-200c was significantly
reduced in the PD >6 months group. This finding suggests that
microRNA-200c may be involved in the EMT process.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (No. 30871169/c140105)
and the Chinese Ministry of Health clinical disciplines key project
(No. 2007-353).
References
|
1.
|
Smit W, Schouten N, van den Berg N,
Langedijk MJ, Struijk DG and Krediet RT; Netherlands
Ultrafiltration Failure Study Group: Analysis of the prevalence and
causes of ultrafiltration failure during long-term peritoneal
dialysis: a cross-sectional study. Perit Dial Int. 24:562–570.
2004.
|
|
2.
|
Kim YL: Update on mechanisms of
ultrafiltration failure. Perit Dial Int. 29(Suppl 2): S123–S127.
2009.PubMed/NCBI
|
|
3.
|
Selgas R, Bajo A, Jiménez-Heffernan JA,
Sánchez-Tomero JA, Del Peso G, Aguilera A and López-Cabrera M:
Epithelialto-mesenchymal transition of the mesothelial cell - its
role in the response of the peritoneum to dialysis. Nephrol Dial
Transplant. 21(Suppl 2): ii2–ii7. 2006.PubMed/NCBI
|
|
4.
|
Aroeira LS, Aguilera A, Sánchez-Tomero JA,
et al: Epithelial to mesenchymal transition and peritoneal membrane
failure in peritoneal dialysis patients: pathologic significance
and potential therapeutic interventions. J Am Soc Nephrol.
18:2004–2013. 2007. View Article : Google Scholar
|
|
5.
|
Elsurer R, Afsar B, Sezer S and Ozdemir
FN: Peritoneal cells at admission: do they have prognostic
significance in peritonitis? Ren Fail. 32:335–342. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Wienholds E, Kloosterman WP, Miska E, et
al: MicroRNA expression in zebrafish embryonic development.
Science. 309:310–311. 2005. View Article : Google Scholar
|
|
7.
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Kato M, Zhang J, Wang M, et al:
MicroRNA-192 in diabetic kidney glomeruli and its function in
TGF-beta-induced collagen expression via inhibition of E-box
repressors. Proc Natl Acad Sci USA. 104:3432–3437. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Ogawa T, Iizuka M, Sekiya Y, et al:
Suppression of type I collagen production by microRNA-29b in
cultured human stellate cells. Biochem Biophys Res Commun.
391:316–321. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Kong D, Li Y, Wang Z, et al: miR-200
regulates PDGF-D-mediated epithelial-mesenchymal transition,
adhesion, and invasion of prostate cancer cells. Stem Cells.
27:1712–1721. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Sabatel C, Cornet AM, Tabruyn SP, et al:
Sprouty1, a new target of the angiostatic agent 16K prolactin,
negatively regulates angiogenesis. Mol Cancer. 9:2312010.
View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Jeffrey SS: Cancer biomarker profiling
with microRNAs. Nat Biotechnol. 26:400–401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Cortez MA, Bueso-Ramos C, Ferdin J, et al:
MicroRNAs in body fluids - the mix of hormones and biomarkers. Nat
Rev Clin Oncol. 8:467–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Ge Y, Xiao L, Chen X, et al: MicroRNAs in
peritoneal dialysis effluent are promising biomarkers for
peritoneal fibrosis in peritoneal dialysis patients. Med
Hypotheses. 78:155–156. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Wang B, Koh P, Winbanks C, et al: miR-200a
prevents renal fibrogenesis through repression of TGF-β2
expression. Diabetes. 60:280–287. 2011.PubMed/NCBI
|
|
16.
|
Yang S, Banerjee S, de Freitas A, et al:
Participation of miR-200 in pulmonary fibrosis. Am J Pathol.
180:484–493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Iwano M, Plieth D, Danoff TM, et al:
Evidence that fibroblasts derive from epithelium during tissue
fibrosis. J Clin Invest. 110:341–350. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Williams JD, Craig KJ, Topley N and
Williams GT: Peritoneal dialysis: changes to the structure of the
peritoneal membrane and potential for biocompatible solutions.
Kidney Int Suppl. 84:S158–S161. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Williams JD, Craig KJ, Topley N, et al:
Peritoneal Biopsy Study Group: Morphologic changes in the
peritoneal membrane of patients with renal disease. J Am Soc
Nephrol. 13:470–479. 2002.PubMed/NCBI
|
|
20.
|
Yáñez-Mó M, Lara-Pezzi E, Selgas R, et al:
Peritoneal dialysis and epithelial-to-mesenchymal transition of
mesothelial cells. N Engl J Med. 348:403–413. 2003.
|
|
21.
|
Jiménez-Heffernan JA, Aguilera A, Aroeira
LS, et al: Immunohistochemical characterization of fibroblast
subpopulations in normal peritoneal tissue and in peritoneal
dialysis-induced fibrosis. Virchows Arch. 444:247–256. 2004.
|
|
22.
|
Schilte MN, Loureiro J, Keuning ED, ter
Wee PM, Celie JW, Beelen RH and van den Born J: Long-term
intervention with heparins in a rat model of peritoneal dialysis.
Perit Dial Int. 29:26–35. 2009.PubMed/NCBI
|
|
23.
|
Margetts PJ, Bonniaud P, Liu L, Hoff CM,
Holmes CJ, West-Mays JA and Kelly MM: Transient overexpression of
TGF-{beta}1 induces epithelial mesenchymal transition in the rodent
peritoneum. J Am Soc Nephrol. 16:425–436. 2005.
|
|
24.
|
Del Peso G, Jiménez-Heffernan JA, Bajo MA,
et al: Epithelial-to-mesenchymal transition of mesothelial cells is
an early event during peritoneal dialysis and is associated with
high peritoneal transport. Kidney Int Suppl. 108:S26–S33.
2008.PubMed/NCBI
|