Introduction
Bladder carcinoma is the most common malignancy of
the urinary tract in China, while transitional cell carcinoma is
the most commonly diagnosed urothelial tumor (1). The prognosis of patients with
non-muscle invasive bladder cancer is good, with five-year survival
rates of 82–100%; however, patients with metastatic urothelial
cancer have a poorer prognosis, with two-year survival rates of
only 5–10% (2). The tumor cells
develop a high tolerance for intrinsic and extrinsic defense
systems and therapeutic procedures. Furthermore, tumor cells may
infiltrate into the adjacent tissues and metastasize to remote
organs and tissues and cause bleeding, infection and dystrophy, in
addition to disrupting important organ functions. Ultimately, tumor
cells migrate and invade various organs, which leads to the
mortality of the patient. At present, an effective therapy for
metastatic urothelial cancer remains unavailable.
Temperature-sensitive transient receptor potential
vanilloid (TRPV) channels are critical contributors to normal pain
and temperature sensations. These channels are
Ca2+-permeable and contribute to intracellular
Ca2+ homeostasis. However, the regulatory mechanism and
the role of the TRPV2 channel in carcinogenesis has not yet been
elucidated. TRPV2, the second member of the TRPV superfamily, was
initially known as vanilloid receptor-like protein 1 and shares 50%
homology with TRPV1 (3). TRPV2
contains six transmembrane domains that consist of a putative
pore-loop region, a cytoplasmic amino terminus with three
ankyrin-repeat domains, and a cytoplasmic carboxy terminus. As a
nonselective cation channel with high Ca2+ permeability,
it also acts as a heat sensor, with a temperature threshold of
50–52°C (4) and may be activated
by 2-aminoethoxydiphenyl borate (5) and insulin-like growth factor-1
(6). TRPV2 is widely distributed
in human organs and tissues, such as the brain, vascular smooth
muscle cells, the gastrointestinal tract, macrophages and the
urothelial tract (7). Furthermore,
TRPV2 has a wide range of physiological and pathological functions
(8). Previous studies have shown
that TPRV2 may be clinically associated with cancer (9–11),
particularly urinary tract tumors (3,12,13).
TRPV2 expression levels have been directly correlated with the
tumor stage and grade of urothelial carcinoma (UC) of the human
bladder (14). It has also been
demonstrated that TRPV2 activation induces apoptotic cell death in
human T24 bladder cancer cells (15). However, the role of TRPV2 in
bladder cancer development and progression remains unclear.
The aim of this study was to investigate the effects
of TRPV2 on the proliferation, migration and invasiveness of 5637
bladder cancer cells, which are characterized by low TRPV2
expression.
Materials and methods
Cell culture
Human 5637 bladder carcinoma cells were obtained
from the American Type Culture Collection (Manassas, VA, USA) and
cultured in RPMI-1640 medium (Gibco-BRL, Grand Island, NY, USA)
supplemented with 100 IU ml−1 penicillin G sodium, 100
μg ml−1 streptomycin sulfate and 10% fetal bovine
serum (FBS; Gibco-BRL) in a humidified 95% air and 5%
CO2 atmosphere at 37°C.
Permanent transfection of 5637 cells with
TRPV2 cDNA
The 5637 cells were plated on a six-well plate and
transfected at ~85% confluence with the rat TRPV2 encoding vector,
pcDNA3.1 (+), using Lipofectamine® 2000 (Invitrogen Life
Technologies, Carlsbad, CA, USA), in accordance with the
manufacturer’s instructions. The stably transfected clones were
selected using Geneticin® G418 (Sigma, St. Louis, MO,
USA) at 400 μg ml−1. Seven clones were identified
using reverse transcription-polymerase chain reaction (RT-PCR) and
western blot analysis. The selected clones were subcloned and
maintained under selection pressure for an additional week.
RT-PCR
Total mRNA was isolated from cells using TRIzol
reagent (Invitrogen Life Technologies), in accordance with the
manufacturer’s instructions. Briefly, 2 μg total RNA was
reverse-transcribed with oligo-d(T) (Invitrogen Life Technologies)
and ThermoScript™ reverse transcriptase (Invitrogen Life
Technologies) in a final reaction volume of 20 μl.
Subsequently, 5% of the samples were amplified by PCR, using the
primers listed in Table I. The
primer sequences were designed using Primer Express Software (PE
Biosystems, Foster City, CA, USA) and synthesized by Invitrogen
(Shanghai, China). Two pairs of TRPV2 primers, which are absent in
human TRPV2, were designed using the rat TRPV2 mRNA as a template
to confirm whether the plasmid was successfully transfected and
expressed at the mRNA level. Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was used for the quantification of the sample
DNA amplification. The DNA amplification conditions included an
initial denaturation step at 95°C for 5 min; 30 cycles at 95°C for
30 sec, 60°C for 30 sec, 72°C for 30 sec; and a final extension
step at 72°C for 7 min.
 | Table I.List of primers used for RT-PCR
amplification. |
Table I.
List of primers used for RT-PCR
amplification.
| Primer | 5′-forward-3′ | 5′-reverse-3′ | Product size
(bp) | Accession no. |
|---|
| TRPV2 (a) |
GTGACGGAACAGCCCACGGT |
CAGTGATGCCTGGCCCTGATGG | 475 | NM_001270797.1 |
| TRPV2 (b) |
AACAAGGGGAAGCAGGAACCGC |
GGCATTGACGAGGGGCTTGGG | 390 | NM_001270797.1 |
| GAPDH |
CGCTCCTGGAAGATGGTGAT |
ACGGATTTGGTCGTATTGGG | 214 | NM_002046.4 |
Western blot assay
The protein expression of TRPV2, matrix
metalloproteinase 2 (MMP2), and GAPDH was assayed by western blot
analysis. Equal quantities of the protein (30 μg) were
separated using 10% sodium dodecyl sulfate polyacrylamide gel
electrophoresis and transferred onto enhanced chemiluminescence
nitrocellulose membranes (Amersham Biosciences, Piscataway, NJ,
USA). Following this, anti-TRPV2-specific antibodies (code:
sc-30155; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
[1:250 (v/v) with non-fat milk], MMP2 antibodies (code: 4022, Cell
Signaling Technology, Inc., Danvers, MA, USA) [1:400 (v/v) with
non-fat milk], and anti-GAPDH-specific antibodies (code: sc-137179,
Santa Cruz Biotechnology, Inc.) [1:500 (v/v) with non-fat milk]
were used for the analysis. Western blot analysis was performed as
previously described (16). Each
experiment was repeated three times with similar results. One
representative experiment is shown.
Cell proliferation assay
A
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
colorimetric assay was used to measure the cell proliferation.
Briefly, the cells were plated at the initial density of 500 per
well in 96-well plates (Corning Life Sciences, Corning, NY, USA),
and the medium was changed 24 h later (day 0). Thereafter, until
day seven, the medium was changed daily. The MTT assay was
performed in accordance with the manufacturer’s instructions
(Sigma). The absorbance at 570 nm was quantified on a microplate
spectrophotometer (ASYS-Hitech GmbH, Municipality of Eugendorf,
Austria).
Cell cycle assay
The cells (~5×105 per well) were
incubated until 85% confluence and digested with 0.25% trypsin
(Gibco-BRL). The cells were subsequently harvested and fixed
overnight with 70% ethanol in phosphate-buffered saline (PBS; added
dropwise) at 4°C and then resuspended in PBS containing 40
μg ml−1 propidium iodide, 0.1 mg ml−1
RNase, and 0.1% Triton X-100 in a dark room. Following incubation
at 37°C for 30 min, the cells were analyzed using a flow cytometer
(Becton-Dickinson, San Jose, CA, USA) equipped with an argon ion
laser at a wavelength of 488 nm. The cell cycle stage was then
determined and analyzed.
Scratch motility assay
The cells were cultured for 24 h as confluent
monolayers in complete medium and then wounded by moving them
across the well with a standard 200 μl pipette tip. The
wounded monolayers were then washed twice to remove non-adherent
cells. Wound closure was monitored for 24 h from initial wounding
using an inverted phase contrast microscope (Leica, Wetzlar,
Germany). Wound closure was monitored for 24 h, as this was shorter
than the doubling time of the 5637 cells. The distance between
borders was estimated using four different fields from each sample.
Four equidistant points in each image were measured to obtain a
better estimate of the true width of the wounded area. The
migration rate was expressed as a percentage of the control (5637
cells, 0 h) and calculated as the proportion of the mean distance
between the borderlines caused by scratching and the distance that
remained cell-free following regrowth. Three independent series of
experiments were performed in quadruplicate.
Transwell assay
The cells were seeded on the top of 8.0-μm
pore Transwell cell culture inserts (Corning Life Sciences), which
were paved with Matrigel glue (diluted 1:4 with serum-free
RPMI-1640 medium; Millipore, Billerica, MA, USA) at a density of
50,000 cells per well (24-well plate) in serum-free culture medium
containing 0.1% bovine serum albumin. Subsequent to culture, the
cells were stimulated to migrate across the filters using 10% FBS
as the chemoattractant in the assay chambers. Following 24 h of
incubation at 37°C, the noninvading cells on the Transwell plates
were scraped off with a cotton swab, whereas the cells that
migrated through the filter pores to the lower surface of the
inserts were fixed for 30 min with 4% paraformaldehyde in PBS and
stained with 0.1% crystal violet for 20 min. The cells under each
filter were counted on five random examination fields
(magnification, ×200) using an inverted phase contrast microscope
(Leica). The data are expressed as the mean of four wells ±
standard error of the mean.
Statistical analysis
SPSS statistical software for Windows version 17.0
(SPSS, Inc., Chicago, IL, USA) was used to conduct the statistical
analysis. All data are presented as the mean ± standard error of
the mean. Each experiment was repeated at least three times. ‘n’
indicates the number of the cells per experiment, whereas ‘N’
indicates the number of experiments performed. A Tukey-Kramer test
was used for statistical comparisons of the means and differences
and P<0.05 was considered to indicate a statistically
significant difference.
Results
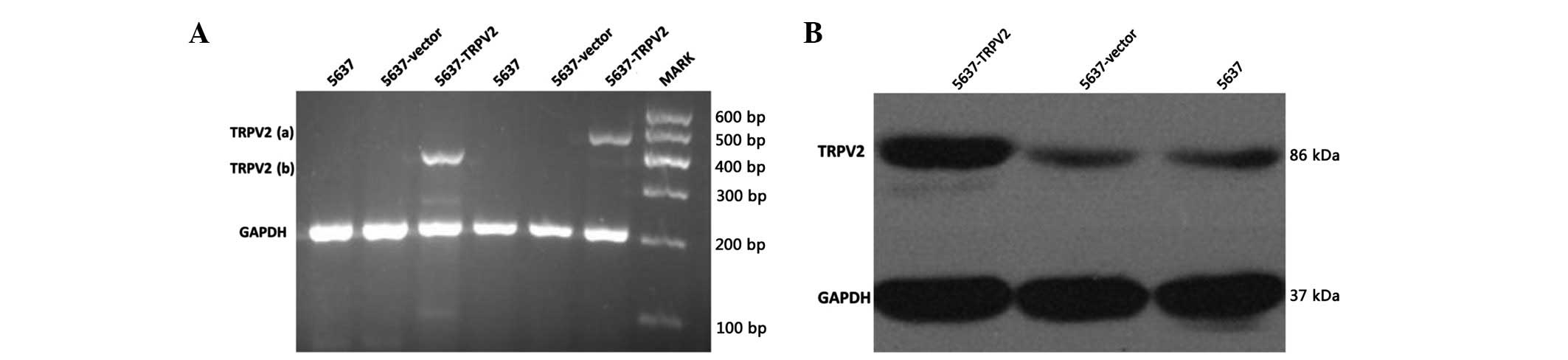
Detection of TRPV2 protein in 5637-TRPV2,
5637-vector, and 5637 cells
The two expected bands were detected in 5637-TRPV2
cells via an RT-PCR assay using specific primers (Fig. 1A). The result demonstrated that the
plasmid was successfully transfected into the 5637 cells. The TRPV2
protein expression level was determined using western blot analysis
(Fig. 1B). The TRPV2 protein
expression levels in the 5637-TRPV2 cells were significantly higher
than in the other cells, which indicated that the transfected
plasmid was expressed at both the mRNA and protein levels.
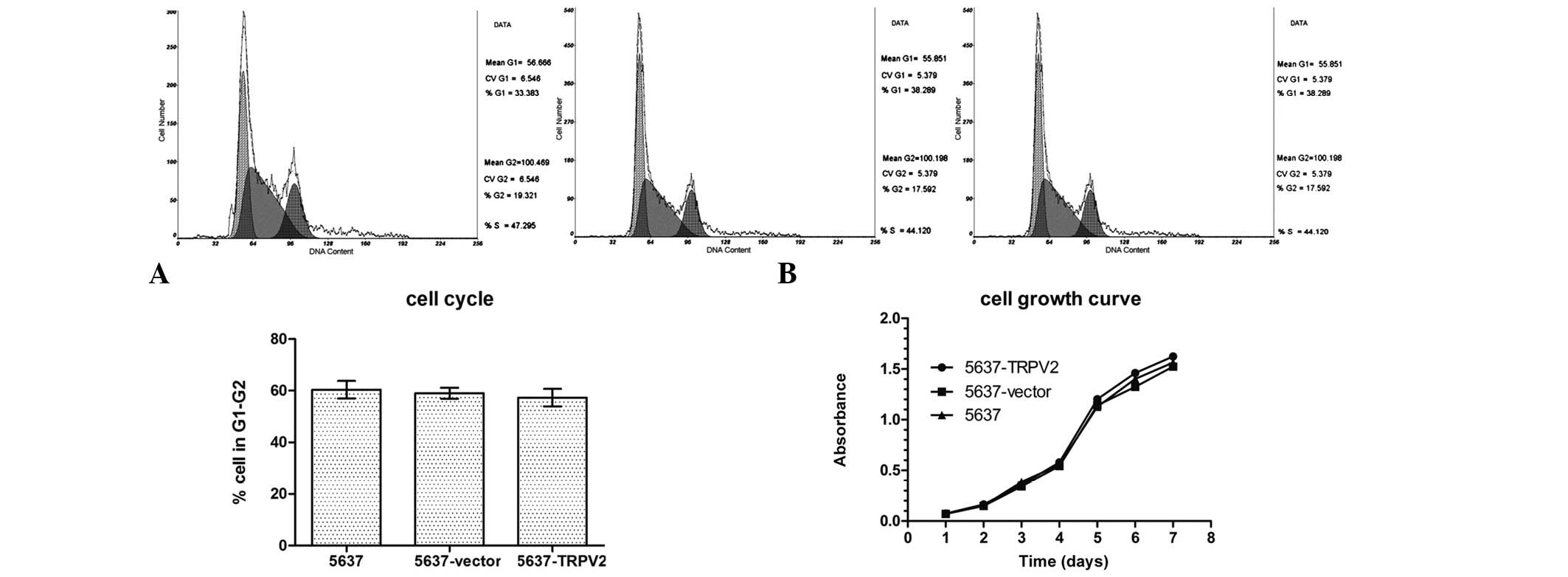
Effects of TRPV2 on 5637 cell
proliferation
Cell proliferation was evaluated in terms of cell
cycle distribution using flow cytometry. The percentage of cells in
the G1–G2 stage was 57.32±5.89% for the 5637-TRPV2 group,
59.04±3.72% for the 5637-vector group, and 60.36±5.89% for the 5637
group. These results did not indicate any significant differences
among the three cell groups (Fig.
2A). The results of the MTT assay also indicated a lack of
significant differences among the cell growth curves of the three
groups. These results suggested that the growth rate of the 5637
cells was unaffected by the TRPV2 channels (Fig. 2B) and that TRPV2 channels did not
affect 5637 cell proliferation.
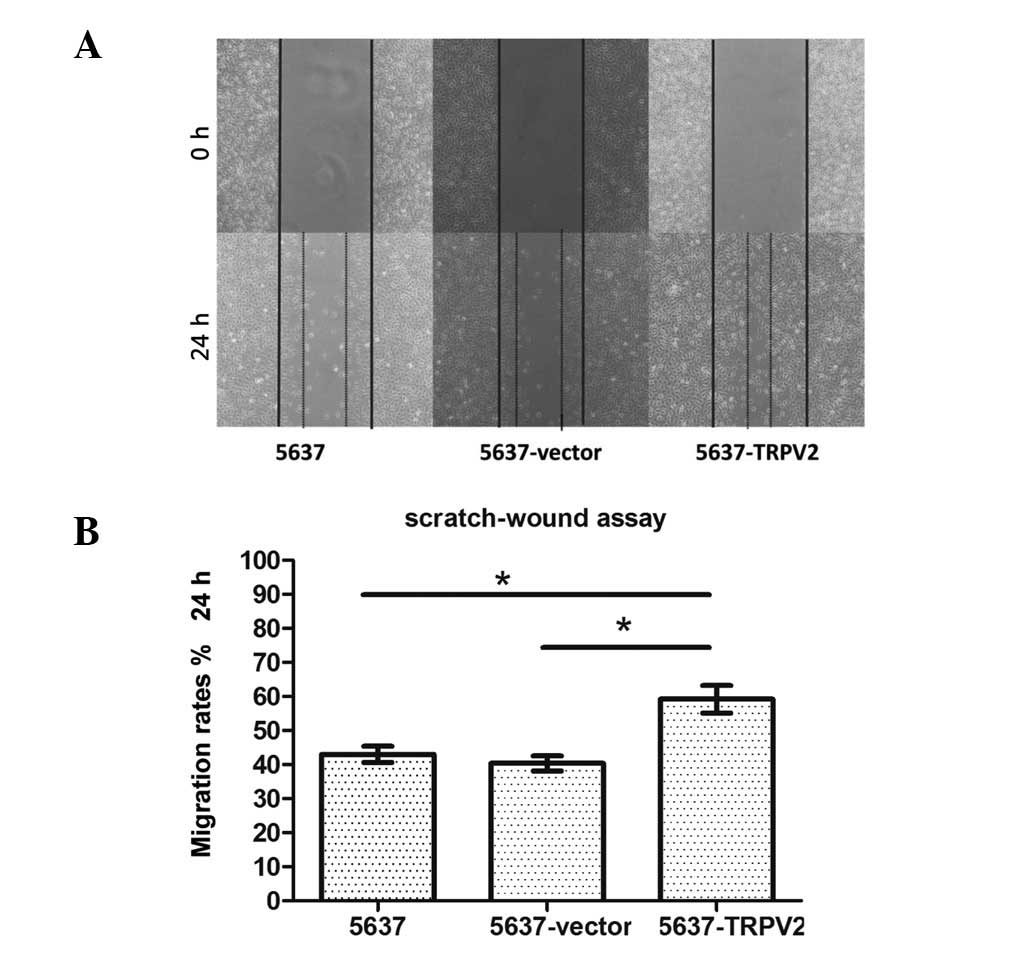
Effects of TRPV2 on 5637 cell
migration
A scratch-wound assay was used to observe the
effects of TRPV2 on cell migration. Following 24 h of incubation,
the motility of the 5637-TRPV2 cells was significantly higher than
that of the cells in the 5637 and 5637-vector groups (Fig. 3A). Wound areas were measured and
normalized relative to the control values (5637 at 0 h), which were
assumed to be 100%. The results showed that the cell migration rate
of the 5637-TRPV2 group (59.21±4.04%) was significantly enhanced
compared with that of the 5637 (42.99±2.37%) and 5637-vector
(40.34±2.24%) groups (n=4, N=3; P<0.05; Fig. 3B).
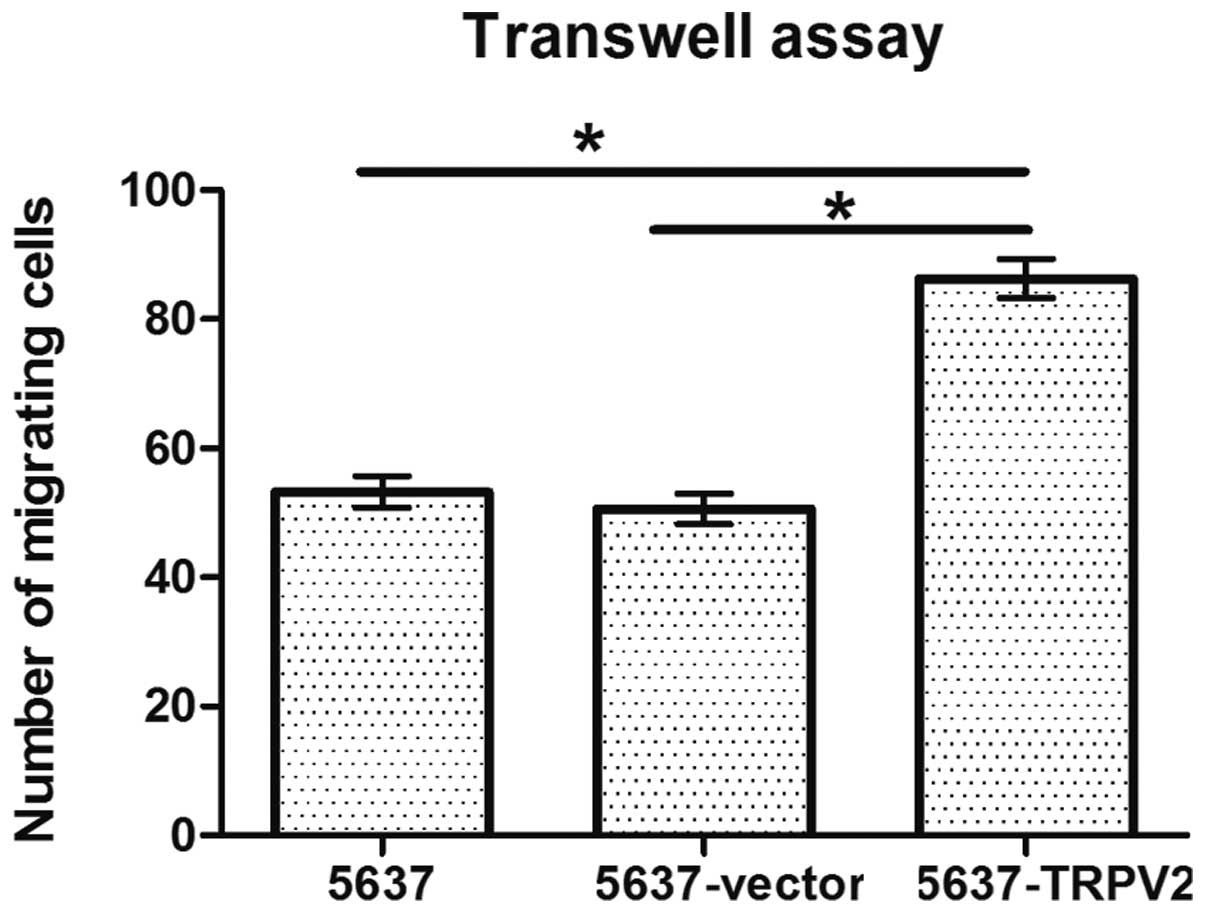
Effects of TRPV2 on 5637 cell
invasion
A Transwell assay was used to observe cell
penetration into the Matrigel glue. The number of migrating cells
in the 5637-TRPV2 group (86.31±7.04) was significantly higher than
that in the 5637 (53.22±4.94) and 5637-vector (50.59±5.91) groups
(n=4, N=3; P<0.05; Fig. 4).
Detection of MMP2 protein in 5637-TRPV2,
5637-vector and 5637 cells
MMP2, a protein closely correlated with tumor cell
migration, was detected using a western blot assay. MMP2 expression
was higher in the 5637-TRPV2 group than in the other two groups
(Fig. 5).
Discussion
The TRPV family is a subclass of the TRP channel
family. It contains six types of calcium-permeable channels that
have unique channel properties (4,17,18).
The TPRV family is expressed in a number of different tissues and
functions to regulate cellular calcium metabolism and calcium
signaling. Members of the TPRV family may also participate in cell
proliferation, differentiation and apoptosis. As one of the
important members of the TPRV superfamily, TRPV2 is highly
expressed in sensory dorsal root ganglion neurons and the
hypothalamus (7). It has been
shown that TRPV2 is closely associated with the urinary system. For
instance, TRPV2 is localized in the superficial cells, whereas
there is negligible or poor expression in the basal and club-shaped
cells in the urothelial cells. TRPV2 channels are expressed in
normal human urothelial cells and in UC of the bladder. However,
the expression level of TRPV2 channels in UC is significantly
higher than in normal urothelial cells and is positively correlated
with the clinical grade and stage of the tumor (14). It has been indicated that TRPV2
channels may be critical in the development and progression of
bladder cancer, but their specific role remains unknown.
Yamada et al (15) studied functional TRPV2 expression
in UC T24 and RT4 cells. Caprodossi et al (14) observed that the TRPV2 expression
levels in UC cells were correlated with high-grade disease.
However, unlike the present study, they focused on the regulation
of calcium influx through TRPV2 channels, which induces apoptotic
cell death in T24 cells. Nevertheless, they did not describe the
changes in the proliferation and migration of the T24 cells under
specific siRNA treatment. In the present study, the
rat-TRPV2-encoding vector was transfected into 5637 cells, which
are characterized by low levels of TRPV2 expression and relatively
weak cell aggression, to verify the precise role of TRPV2 channels
on bladder cancer (Fig. 1B).
Following the selection of the monoclonal cell lines, no
significant difference was observed in cell cycle distribution
using flow cytometric analysis (Fig.
2A). Similar results were obtained from the cell growth curve
in a week (Fig. 2B). These results
indicated that TRPV2 exerted no effect on the cell growth rate and
proliferation. The study also evaluated cell migration ability,
with the results of the scratch-wound assay and the Transwell assay
showing that TRPV2 enhanced 5637 cell migration.
TRPV2 channels contribute to intracellular
Ca2+ homeostasis. Activation of the channels increases
the intracellular free Ca2+ levels, which enables the
Ca2+ to participate in various cellular processes, such
as proliferation, metabolism and gene transcription.
Ca2+ also has a multifunctional role in directional
sensing, cytoskeleton redistribution, traction force generation and
relocation of focal adhesions in tumor cells (19). However, the precise role of TRPV2
in bladder cancer remains unclear. Based on previous studies
(20–22), we inferred that TRPV2 activity may
be mediated by the direct regulation of key proteins, such as MMP2,
which are used by cancer cells for invasion. MMP2 is associated
with bladder cancer invasiveness (23) and its expression is often used to
measure the migration ability of tumor cells. It was observed that
MMP2 expression was significantly higher in 5637-TRPV2 cells than
in the cells of the other two groups (Fig. 5). MMP2 is a
Zn2+-dependent type IV collagenase with a molecular mass
of 72 kDa. It is activated by biochemical interaction with a
transmembrane MMP, called membrane-type (MT)-MMP, or by binding
with integrin αVβl cell surface adhesion receptors. Numerous
studies have demonstrated that MMP2 is critical in cancer
development and progression (21,24–27).
Cell migration is a complex process that requires the coordinated
regulation of cell-cell attachment, cell-matrix attachment and
matrix remodeling. MMP2 directly modulates cell-matrix adhesion by
removing adhesion sites or by exposing binding sites to induce cell
migration (28), and it affects
tumor cell behavior in vivo, due to the ability to cleave
growth factors, cell surface receptors, cell adhesion molecules and
chemokines/cytokines, which promotes tumor metastases (29–31).
Furthermore, MMP2 selects more aggressive phenotypes by generating
apoptosis-resistant cells via the cleavage of proapoptotic factors
(32), in addition to
collaborating with other MMPs to promote cancer-related
angiogenesis. As a result of these functions and roles, MMP2 is an
extremely important protein in bladder cancer development and
progression. The results of the present study suggest that MMP2
expression is increased during TRPV2 overexpression in 5637 cells,
which is consistent with the previously described inference.
In conclusion, the nonselective cationic TRPV2
channel enhances bladder cancer cell migration, but does not affect
cell proliferation in vitro. Furthermore, TRPV2 activity,
which may be mediated by direct MMP2 regulation, is important in
bladder tumor development and progression. These results suggest
that TRPV2 channels are a potential target for therapeutic
approaches to bladder carcinoma. However, the precise role of TRPV2
in bladder cancer in vivo requires further study.
Abbreviations:
|
TRP
|
transient receptor potential
channel;
|
|
MMP2
|
matrix metalloproteinase 2
|
Acknowledgements
This study was supported by the
Fundamental Research Funds for the Central Universities (grant no.
201130302020009).
References
|
1.
|
Chen WQ, Zeng HM, Zheng RS, Zhang SW and
He J: Cancer incidence and mortality in china, 2007. Chin J Cancer
Res. 24:1–8. 2012. View Article : Google Scholar
|
|
2.
|
Elsobky E, El-Baz M, Gomha M, Abol-Enein H
and Shaaban AA: Prognostic value of angiogenesis in
schistosoma-associated squamous cell carcinoma of the urinary
bladder. Urology. 60:69–73. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Prevarskaya N, Flourakis M, Bidaux G,
Thebault S and Skryma R: Differential role of TRP channels in
prostate cancer. Biochem Soc Trans. 35:133–135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Caterina MJ, Rosen TA, Tominaga M, Brake
AJ and Julius D: A capsaicin-receptor homologue with a high
threshold for noxious heat. Nature. 398:436–441. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Neeper MP, Liu Y, Hutchinson TL, Wang Y,
Flores CM and Qin N: Activation properties of heterologously
expressed mammalian TRPV2: evidence for species dependence. J Biol
Chem. 282:15894–15902. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Kanzaki M, Zhang YQ, Mashima H, Li L,
Shibata H and Kojima I: Translocation of a calcium-permeable cation
channel induced by insulin-like growth factor-I. Nat Cell Biol.
1:165–170. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Everaerts W, Gevaert T, Nilius B and De
Ridder D: On the origin of bladder sensing: Tr(i)ps in urology.
Neurourol Urodyn. 27:264–273. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Nilius B, Owsianik G, Voets T and Peters
JA: Transient receptor potential cation channels in disease.
Physiol Rev. 87:165–217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Prevarskaya N, Zhang L and Barritt G: TRP
channels in cancer. Biochim Biophys Acta. 1772:937–946. 2007.
View Article : Google Scholar
|
|
10.
|
Lehen’kyi V and Prevarskaya N: Study of
TRP channels in cancer cells. TRP Channels. Zhu MX: CRC Press; Boca
Raton, FL: Chapter 17. 2011, PubMed/NCBI
|
|
11.
|
Santoni G and Farfariello V: TRP channels
and cancer: new targets for diagnosis and chemotherapy. Endocr
Metab Immune Disord Drug Targets. 11:54–67. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Gkika D and Prevarskaya N: TRP channels in
prostate cancer: the good, the bad and the ugly? Asian J Androl.
13:673–676. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Ziglioli F, Frattini A, Maestroni U,
Dinale F, Ciufifeda M and Cortellini P: Vanilloid-mediated
apoptosis in prostate cancer cells through a TRPV-1 dependent and a
TRPV-1-independent mechanism. Acta Biomed. 80:13–20.
2009.PubMed/NCBI
|
|
14.
|
Caprodossi S, Lucciarini R, Amantini C, et
al: Transient receptor potential vanilloid type 2 (TRPV2)
expression in normal urothelium and in urothelial carcinoma of
human bladder: correlation with the pathologic stage. Eur Urol.
54:612–620. 2008. View Article : Google Scholar
|
|
15.
|
Yamada T, Ueda T, Shibata Y, et al: TRPV2
activation induces apoptotic cell death in human T24 bladder cancer
cells: a potential therapeutic target for bladder cancer. Urology.
76:509.e1–509.e7. 2010. View Article : Google Scholar
|
|
16.
|
Yang ZH, Wang XH, Wang HP and Hu LQ:
Effects of TRPM8 on the proliferation and motility of prostate
cancer PC-3 cells. Asian J Androl. 11:157–165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Caterina MJ, Schumacher MA, Tominaga M,
Rosen TA, Levine JD and Julius D: The capsaicin receptor: a
heat-activated ion channel in the pain pathway. Nature.
389:816–824. 1997. View
Article : Google Scholar
|
|
18.
|
Liedtke W, Choe Y, Martì-Renom MA, et al:
Vanilloid receptor-related osmotically activated channel (VR-OAC),
a candidate vertebrate osmoreceptor. Cell. 103:525–535. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Ridley AJ, Schwartz MA, Burridge K, et al:
Cell migration: integrating signals from front to back. Science.
302:1704–1709. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Monet M, Gkika D, Lehen’kyi V, et al:
Lysophospholipids stimulate prostate cancer cell migration via
TRPV2 channel activation. Biochim Biophys Acta. 1793:528–539. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Chuang CK, Pang ST, Chuang TJ and Liao SK:
Profiling of matrix metalloproteinases and tissue inhibitors of
metalloproteinases proteins in bladder urothelial carcinoma. Oncol
Lett. 1:691–695. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Monet M, Lehen’kyi V, Gackiere F, et al:
Role of cationic channel TRPV2 in promoting prostate cancer
migration and progression to androgen resistance. Cancer Res.
70:1225–1235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Kader AK, Liu J, Shao L, et al: Matrix
metalloproteinase polymorphisms are associated with bladder cancer
invasiveness. Clin Cancer Res. 13:2614–2620. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Nakajima M, Welch DR, Wynn DM, Tsuruo T
and Nicolson GL: Serum and plasma M(r) 92,000 progelatinase levels
correlate with spontaneous metastasis of rat 13762NF mammary
adenocarcinoma. Cancer Res. 53:5802–5807. 1993.PubMed/NCBI
|
|
25.
|
Stearns ME and Wang M: Type IV collagenase
(M(r) 72,000) expression in human prostate: benign and malignant
tissue. Cancer Res. 53:878–883. 1993.PubMed/NCBI
|
|
26.
|
Kanayama H, Yokota K, Kurokawa Y, Murakami
Y, Nishitani M and Kagawa S: Prognostic values of matrix
metalloproteinase-2 and tissue inhibitor of metalloproteinase-2
expression in bladder cancer. Cancer. 82:1359–1366. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Coussens LM, Fingleton B and Matrisian LM:
Matrix metalloproteinase inhibitors and cancer: trials and
tribulations. Science. 295:2387–2392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Giannelli G, Falk-Marzillier J, Schiraldi
O, Stetler-Stevenson WG and Quaranta V: Induction of cell migration
by matrix metal-loprotease-2 cleavage of laminin-5. Science.
277:225–228. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Noë V, Fingleton B, Jacobs K, et al:
Release of an invasion promoter E-cadherin fragment by matrilysin
and stromelysin-1. J Cell Sci. 114:111–118. 2001.PubMed/NCBI
|
|
31.
|
Mañes S, Llorente M, Lacalle RA, et al:
The matrix metalloproteinase-9 regulates the insulin-like growth
factor-triggered autocrine response in DU-145 carcinoma cells. J
Biol Chem. 274:6935–6945. 1999.PubMed/NCBI
|
|
32.
|
Stetler-Stevenson WG: Matrix
metalloproteinases in angiogenesis: a moving target for therapeutic
intervention. J Clin Invest. 103:1237–1241. 1999. View Article : Google Scholar : PubMed/NCBI
|