Introduction
Common traumatic brain injury models are mainly
divided into two types: the impact injury and the acceleration
injury models (1–3). The impact injury model is achieved by
keeping the animal in a stationary state and drilling a window into
the skull. Focal and diffuse brain injuries may be generated by the
impact of a liquid or the steel body of the drill on the dura
mater, resulting in damage to the brain tissue via the bone window.
The model demonstrates good experimental control and
reproducibility. The acceleration injury model (3) is further subdivided into two models:
simple inertial acceleration and acceleration impact injuries. The
former entails fixing the animal in a controllable acceleration
device and causing a linear or rotational acceleration motion
injury, according to the design requirements, rather than by
directly impacting the brain. The significant feature of this model
is that it enables independent investigation of the inertia damage
mechanism. The acceleration impact injury is characterized by the
traditional Feeney’s falling body method (4). In this model, the head of the animal
is kept stationary and a heavy object is dropped from a high
position to impact and damage the head. The main features of the
acceleration impact injury model include the fact that it is
possible to adjust the extent of the injury by altering the height
that the object falls from and the weight of the object.
Furthermore, it is easy and simple to repeat the acceleration brain
injury.
However, these animal models only consider brain
injuries caused by the transmission of external forces to the
brain, and ignore the fact that brain is composed of multiple
tissues rather than being one homogeneous body. The mechanical
properties of the tissues comprising the brain are inconsistent and
there is a certain volume of cerebrospinal fluid in the brain
ventricles. The cerebrospinal fluid and the surrounding brain
tissue are two different types of substance. Furthermore, the
cerebrospinal fluid is incompressible (5,6). In
the deceleration injury process, the brain tissue impacts the skull
over a short stretch of time, while the cerebrospinal fluid impacts
the ventricular wall more slowly. The impact caused by the
cerebrospinal fluid moving with a certain energy results in
injuries to the periventricular structures.
Cell apoptosis may be observed in various traumatic
brain injury models, and studies have suggested that cell apoptosis
is involved in the whole process of secondary pathophysiological
evolution following craniocerebral injury (7,8).
However, to the best of our knowledge, there have not been any
studies investigating whether cerebrospinal fluid moving with a
certain energy causes injuries to periventricular structures,
resulting in cell apoptosis in the deceleration injury process.
Therefore, the authors designed a ventricular fluid impact model,
in order to simulate the situation of the cerebrospinal fluid
impacting the ventricular wall during the deceleration injury
process. The aim of this was to observe the changes in various
indicators of the animal at different levels of energy and to
investigate the occurrence of apoptosis of the nerve cells in the
periventricular structures, the hippocampus and
thalamencephalon.
Materials and methods
Grouping
A total of 88 New Zealand rabbits with a mean body
weight of 2.5±0.3 kg (provided by the Experimental Zoology
Department of Central South University, Changsha, China) were
randomly divided into normal control, surgical control and injury
groups. For the former two groups, each group comprised eight
rabbits, whereas the injury group comprised 72 rabbits. According
to the principle of randomization, the injury group was further
divided into the following three subgroups: mild, moderate and
severe injury (24 rabbits in each subgroup). Eight rabbits were
randomly selected from each injury group to determine consciousness
recovery time following trauma (in the mild, moderate and severe
injury groups there were eight, seven and three survival cases
used, respectively, to determine the consciousness recovery time
following trauma). Variations in respiration, blood pressure and
heart rate were monitored. In each group, the remaining 16 rabbits,
respectively, were used for pathological examinations following
brain perfusion fixation conducted at 12 h and 1, 2, 3 and 7 days
subsequent to trauma (in the mild, moderate and severe injury
groups, there were 15, 13 and 10 survival cases, respectively, used
for the determination of the apoptosis-positive cell count). This
study was carried out in strict accordance with the recommendations
in the 8th Edition of the Guide for the Care and Use of Laboratory
Animals (January 1, 2012). The animal use protocol was reviewed and
approved by the Institutional Animal Care and Use Committee (IACUC)
of Xiangya Hospital of Central South University.
Device used for animal brain injury
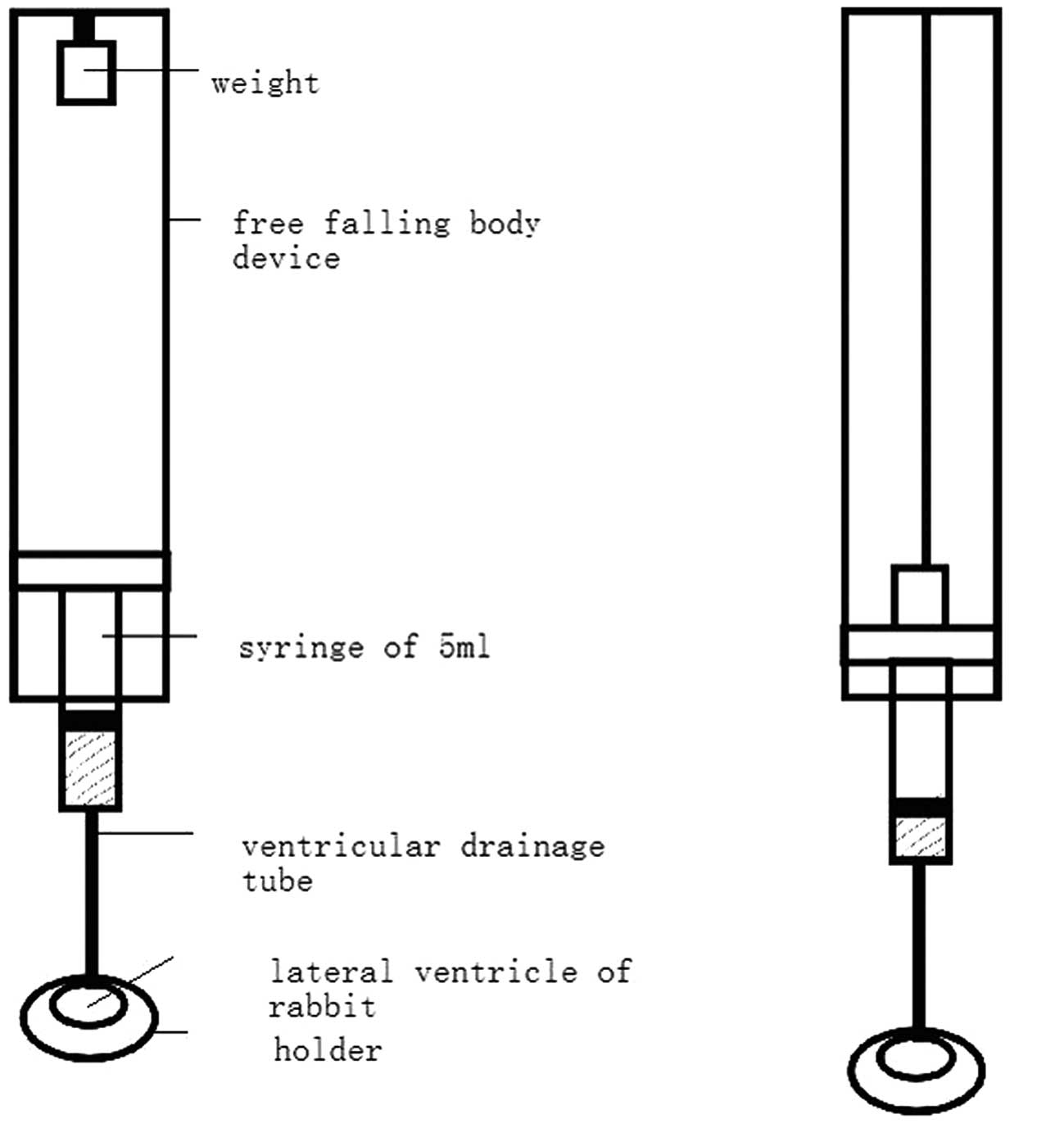
The device used for the animal brain injury
consisted of five components: a weight, a free falling body device
(reference to Feeney’s method), a 5-ml syringe (adequately
lubricated), a ventricular drainage tube and a holder. The lower
part of the free falling body device was connected to the syringe,
while the lower part of the syringe was connected to the lateral
ventricle through the ventricular drainage tube. The top part of
the weight was fixed with a threadlet onto the upper part of the
device. The threadlet length was adjusted. As the weight dropped to
impact the upper syringe core and move it down by 0.8 cm (~1.0 ml
liquid flowed out from the syringe), the threadlet over the weight
was tightened, and the weight did not drop again. Therefore, the
liquid entered the lateral ventricle through the ventricular
drainage tube. Immediately subsequent to this, the holder was moved
away (Fig. 1).
Surgical technique and experimental
procedure
Nembutal solution (30 g/l) was slowly injected at a
dose of 33 mg/kg via an intravenous injection to the marginal ear
vein. Following successful anaesthesia, a rabbit was fixed onto the
holder in prone position and, under aseptic conditions, the sutura
was exposed. In accordance with the method by Kockro et
al(9), a small hole with a
diameter of 1.5 mm was drilled by a dental drill on the right side
of 4 mm of the middle line at the crown drill joint, prior to the
venous indwelling needle (outer diameter of 1.1 mm, replacing the
lateral ventricular drainage tube) being inserted by 4–6 mm.
Following the removal of the internal core, the cerebrospinal fluid
rose in the silicone tube, indicating the success of the puncture,
and it was fixed with dental resin. The right femoral artery
catheter was connected to a blood pressure monitor and the femoral
vein catheter was connected to the infusion set. Heart rate and
respiration were monitored by a physiological recorder. A weight of
100 g fell from heights of 23, 53 and 94 cm for the mild, moderate
and severe injury groups, respectively, to impact the upper syringe
core (according to the energy conservation law mgh=½mv2,
the converted impact energy was equivalent to the energy generated
by 15 ml cerebrospinal fluid in a human unilateral ventricle
momentarily impacting the ventricle wall at speeds of 20, 30 and 40
km/h, respectively, prior to slowing and stopping). Subsequently,
the holder was immediately moved away and the ventricular drainage
tube was removed. An injection of gentamycin sulfate (40,000 U) was
dripped onto the incision for 4–5 days, prior to the bone window
being sealed with bone wax and the scalp being sutured. For the
surgical control group, only the lateral ventricle puncture was
conducted, without the impact procedure. For the normal control
group, no intervention was conducted.
Specimen acquisition
The groups of animals were conventionally
anaesthetized, respectively, at the preset time-points. Following
perfusion fixation with 4% paraformaldehyde, brain tissues were
extracted, conventionally dehydrated, hyalinized and embedded with
paraffin wax for hematoxylin and eosin (H&E) and
immunohistochemical staining.
Analysis of results
Cell apoptosis was evaluated with the cell apoptosis
detection kit provided by Boster Biotecimology Ltd., Co. (Wuhan,
China) according to the terminal deoxynucleotidyl transferase dUTP
nick end labeling (TUNEL) method. If brown granules were present in
the cell nucleus, the cell was positive for apoptosis. In
combination with positive H&E staining results, where cells
that were positive for apoptosis displayed circular and shrinkage
morphology characteristics, cells with nuclear pyknosis, deep cell
staining and structural integrity were considered to be apoptotic
nerve cells. Under the light microscope, five fields of view were
randomly selected to conduct the cell count. The mean positive cell
count of 100 cells was calculated and taken as the apoptosis index
(10).
Statistical analysis
All data are expressed as the mean ± standard
deviation. For a comparison of the same indicator at different
time-points, a t-test was used. One-way analysis of variance
(ANOVA) was used for comparisons of data from different injury
groups, and a non-parameter Kruskal-Wallis test was used to
evaluate inter-group heterogeneity of variance. SPSS 13.0
statistical software (SPSS, Inc., Chicago, IL, USA) was used for
statistical processing. P<0.05 was considered to indicate a
statistically significant difference.
Results
Pathophysiological changes
The mortality rates for the mild, moderate and
severe injury groups were 4.16, 16.67 and 45.83%, respectively.
There were significant differences between the three groups
(P<0.05). The time of death of all cases was within 1 h
subsequent to trauma, and there were no deaths in the normal or
surgical control groups.
In contrast to the normal and surgical control
groups, the mild and moderate injury groups immediately presented
with exaggerated and deep respiration following trauma, while the
severe injury group immediately presented with apnea. Subsequent to
the respirator being connected, the majority of the surviving
animals regained autonomous respiration within 10–15 min following
the trauma. Compared with the normal and surgical control groups,
the differences at 2 h subsequent to trauma were not
significant.
All the traumatized animals immediately demonstrated
an elevation in mean arterial pressure following the trauma, with
the pressure reaching a peak at 5 sec subsequent to the trauma. The
animals then presented with hypopiesia for some time. Within 15–30
min, the mean arterial pressure returned to the level prior to
trauma, whereas the blood pressure of the fatally injured animals
continually dropped until death.
The mean heart rate of the animals that survived
following trauma rapidly decreased from 238±39 beats/min to 160±61
beats/min and gradually returned to the level prior to trauma
within 15–30 min. By contrast, the mean heart rate of the fatally
injured animals decreased to 50% of the level prior to trauma in 2
min and then continually declined until death.
The corneal reflex was normal in the normal control
and the surgical control groups. In the case of acupuncture, the
animals presented evasive actions and were able to crawl by
themselves. It was indicated that consciousness returned to the
normal level (11) (Table I).
 | Table IConsciousness recovery time of rabbits
following brain injury. |
Table I
Consciousness recovery time of rabbits
following brain injury.
| Group | n | Consciousness
recovery time (h) |
|---|
| Normal control | 8 | 2.00±0.39 |
| Surgical control | 8 | 2.11±0.40 |
| Mild injury | 8 | 2.43±1.20 |
| Moderate injury | 7 | 5.85±1.23 |
| Severe injury | 3 | 7.83±0.40 |
Pathological changes
Examinations of the normal control and surgical
control groups under a light microscope were normal. H&E
staining of the hippocampus showed that the morphological
structures of the hippocampus in the mild injury group were normal
at various time-points subsequent to trauma. By contrast, the
morphological structures of the hippocampus in the moderate injury
group at 24 h subsequent to trauma underwent significant changes;
the tissues were loose and hemorrhagic foci were visible. Similar
results were observed in the severe injury group at 8 h subsequent
to trauma (Fig. 2A), i.e. the
morphological structures of the hippocampus were significantly
changed; however, the changes were more marked than those of the
moderate injury group. H&E staining of the thalamencephalon
showed that the morphological structures of the thalamencephalon in
the mild injury group were normal at various time-points subsequent
to trauma, although slight intercellular edema was present at 24 h.
The cellular structures of the moderate injury group at 24 h were
extensively changed; intercellular edema was present and few or no
hemorrhagic foci were visible; by contrast, the changes in the
cellular structures of the severe injury group at 8 h were more
evident, nuclear pyknosis was visible and there was marked
intercellular edema (Fig. 2B).
TUNEL staining showed that very few cells were
positive for apoptosis in the normal control and surgical control
groups. With regard to the injury groups, TUNEL staining of the
hippocampus showed that there was a small number of cells positive
for apoptosis in the mild injury group at 48 h subsequent to trauma
and the peak number was reached at 72 h subsequent to trauma. In
the moderate and severe injury groups, apoptotic cells were present
at 24 h subsequent to trauma and the peak number was reached at 72
h (Fig. 3). At 7 days subsequent
to trauma, levels of apoptotic cells had dropped markedly (Table II).
| Table IIComparison of the apoptosis index in
the hippocampal CA1 region of rabbits following trauma. |
Table II
Comparison of the apoptosis index in
the hippocampal CA1 region of rabbits following trauma.
| | Time after
trauma |
|---|
| |
|
|---|
| Group | n | 12 h | 1 day | 2 days | 3 days | 7 days |
|---|
| Normal control | 8 | 1.03±1.32 | - | - | - | - |
| Surgical control | 8 | 1.01±0.90 | - | - | - | - |
| Mild injury | 15 | 1.21±1.21 | 1.13±0.27 | 8.19±2.13 | 12.18±1.96 | 7.56±2.74 |
| Moderate injury | 13 | 1.97±0.88 | 6.21±2.37 | 12.15±2.19 |
18.44±3.97a | 8.93±2.51 |
| Severe injury | 10 | 1.13±1.12 | 8.23±1.81 | 15.35±3.11 |
27.18±5.05b | 9.12±3.04 |
The trend revealed by the TUNEL staining of the
thalamencephalon was consistent with that of the hippocampal
staining. In the mild injury group a small number of apoptotic
cells were present at 48 h subsequent to trauma, and the number
peaked at 72 h subsequent to trauma. Apoptotic cells were observed
in the moderate and severe injury groups at 24 h, with the number
peaking at 72 h (Fig. 4) and
markedly declining at 7 days (Table
III).
| Table IIIComparisons of the apoptosis index in
the thalamencephalon of rabbits following trauma. |
Table III
Comparisons of the apoptosis index in
the thalamencephalon of rabbits following trauma.
| | Time subsequent to
trauma |
|---|
| |
|
|---|
| Group | n | 12 h | 1 day | 2 days | 3 days | 7 days |
|---|
| Normal control | 8 | 1.72±0.86 | - | - | - | - |
| Surgical control | 8 | 1.84±0.73 | - | - | - | - |
| Mild injury | 8 | 1.89±0.82 | 1.88±0.87 | 16.38±5.92 | 27.29±8.93 | 13.92±4.21 |
| Moderate injury | 8 | 1.91±0.76 | 17.45±6.73 | 19.68±8.81 |
34.25±6.25a | 12.88±3.27 |
| Severe injury | 8 | 1.80±0.72 | 20.17±8.84 | 29.11±9.60 |
43.47±10.29b | 13.15±6.78 |
Discussion
The majority of common traumatic brain injury animal
models only consider brain injuries caused by the transmission of
external forces to the brain (12–14);
however, these models ignore the fact that the brain is composed of
multiple tissues rather than being one homogeneous substance. The
mechanical properties of these tissues differ and there is a
certain volume of cerebrospinal fluid in the brain ventricles. The
cerebrospinal fluid and the surrounding brain tissue are two
different types of substance (15,16).
Furthermore, the cerebrospinal fluid is incompressible. In the
deceleration injury process, the brain tissue impacts the skull
over a short stretch of time, while the cerebrospinal fluid impacts
the ventricular wall more slowly. The impact caused by the
cerebrospinal fluid moving with a certain energy results in
injuries to the periventricular structures. In this study, based on
the previously mentioned considerations, the mathematical principle
was closely combined with the physical principle and a free falling
body device was used to generate a constant and reproducible graded
energy by energy conservation law calculation, and the energy was
transmitted to the brain ventricle via a ventricular drainage tube
to simulate the situation of the cerebrospinal fluid impacting the
ventricular wall during the deceleration injury process. The aim of
this was to observe the changes in various vital indicators of the
animal at different levels of energy and to investigate the
occurrence of nerve cell apoptosis in periventricular structures,
specifically, in the hippocampus and thalamencephalon.
In the current study, TUNEL positive cell
determination in the hippocampus revealed that injury extent was
correlated with the time of the occurrence of cell apoptosis; the
earlier the occurrence of apoptosis, the greater the number of
apoptotic cells. This suggests that, within a certain energy range,
the greater the energy of the ventricle trauma, the more severe the
extent of hippocampal injury and the greater the number of
hippocampal cells positive for TUNEL staining are likely to be. If
the energy exceeded the energy limit, it is likely to induce the
necrosis of a number of cells. Light microscopy and electron
microscopy studies have demonstrated that, in the case of cerebral
concussion, the cerebral cortex and hippocampus show diffuse
neuronal morphological changes (13,17).
This model indicated that simple ventricular fluid impact induced
neuronal cell apoptosis in the hippocampus constituting the wall of
the ventricular temporal horn. This type of pathological change is
likely to be one of the anatomical bases for cerebral concussion
presenting with dysmnesia. In addition, functional disorder
manifestations of the respiratory cycle of the animals in this
model were similar to the transient brainstem responses following
cerebral concussion. It is possible that, in the case of
deceleration injury, the impact of intraventricular cerebrospinal
fluid on periventricular structures is just a complementary
explanation among cerebral concussion pathogeneses.
Previous studies have suggested that persistent
ischemia and degeneration in the thalamencephalon following trauma
may be important in severe brain trauma, particularly in the case
of animal survival (12,18,19),
since the thalamencephalon is located at the sides of the three
ventricles and the lateral ventricle. In the current study, the
possible impact of ventricular liquid on the lateral ventricle
during deceleration injury was considered, and it was demonstrated
using the TUNEL immunohistochemical staining method that cell
apoptosis was persistent in the thalamencephalon. In this model,
there was no significant difference in the animal consciousness
recovery time between the mild injury and the control groups. The
animal consciousness recovery time of the moderate and severe
injury groups were significantly extended compared with those of
the control group, with the mean animal consciousness recovery time
of the severe injury group reaching 7.83±4.0 h. However, cell
apoptosis in the thalamencephalon occurred relatively early, at 24
h subsequent to trauma. Therefore, extension of the animal
consciousness recovery time in this study was not observed to
correlate with the levels of cell apoptosis. H&E staining
showed that the moderate and severe injury groups presented with
hemorrhagic necrotic changes in the thalamencephalon at 2 h
subsequent to trauma. This suggests that the extension in animal
consciousness recovery time at this time was due to cell necrosis
rather than cell apoptosis in the acute stage of trauma. In the
case of secondary brain injury, the main form of cellular death
becomes apoptosis. This long-term phenomenon of cell apoptosis in
the thalamencephalon, as a result of ventricular liquid impact, may
be one of the important reasons for persistent coma in patients
with craniocerebral trauma subsequent to trauma injury.
In conclusion, nerve cell apoptosis was apparent in
the ventricular fluid impact model, and the reason for the
apoptosis was closely associated with the impact of the ventricular
liquid on the periventricular structures. The results of this study
may be beneficial in the further development of new understandings
of the pathological mechanism of clinical craniocerebral injury and
its pathological changes (20–23).
References
|
1
|
Adelson PD, Fellows-Mayle W, Kochanek PM
and Dixon CE: Morris water maze function and histologic
characterization of two age-at-injury experimental models of
controlled cortical impact in the immature rat. Childs Nerv Sys.
29:43–53. 2013. View Article : Google Scholar
|
|
2
|
Mouzon BC, Chaytow H, Crynen G, et al:
Repetitive mild traumatic brain injury in a mouse model produces
learning and memory deficits accompanied by histological changes. J
Neurotrauma. 29:2761–2773. 2012. View Article : Google Scholar
|
|
3
|
Li Y, Zhang L, Kallakuri S, Zhou R and
Cavanaugh JM: Injury predictors for traumatic axonal injury in a
rodent head impact acceleration model. Stapp Car Crash J. 55:25–47.
2011.PubMed/NCBI
|
|
4
|
Feeney DM, Boyeson MG, Linn RT, Murray HM
and Dail WG: Responses to cortical injury: I. Methodology and local
effects of contusions in the rat. Brain Res. 211:67–77. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dijkers MP, Harrison-Felix C and Marwitz
JH: The traumatic brain injury model systems: history and
contributions to clinical service and research. J Head Trauma
Rehabil. 25:81–91. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morganti-Kossmann MC, Yan E and Bye N:
Animal models of traumatic brain injury: is there an optimal model
to reproduce human brain injury in the laboratory? Injury. 41(Suppl
1): S10–S13. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schwetye KE, Cirrito JR, Esparza TJ, Mac
Donald CL, Holtzman DM and Brody DL: Traumatic brain injury reduces
soluble extracellular amyloid-β in mice: a methodologically novel
combined microdialysis-controlled cortical impact study. Neurobiol
Dis. 40:555–564. 2010.PubMed/NCBI
|
|
8
|
Tashlykov V, Katz Y, Gazit V, Zohar O,
Schreiber S and Pick CG: Apoptotic changes in the cortex and
hippocampus following minimal brain trauma in mice. Brain Res.
1130:197–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kockro RA, Giacomelli R, Scheihing M,
Aschoff A and Hampl JA: A stereotactic device for rabbits based on
mandibular and cranial landmarks: technical note. J Neurosurg.
108:601–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marbacher S, Andereggen L, Neuschmelting
V, et al: A new rabbit model for the study of early brain injury
after subarachonoid hemorrhage. J Neurosci Methods. 208:138–145.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Khan M, Im YB, Shunmugavel A, Gilg AG, et
al: Administration of S-nitrosoglutathione after tramatic brain
injury protects the neurovascular unit and reduces secondary injury
in a rat model of controlled cortical impact. J Neuroinflammation.
6:322009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kao C, Forbes JA, Jermakowicz WJ, et al:
Suppression of thalamocortical oscillations following traumatic
brain injury in rats. J Neurosurg. 117:316–323. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rostami E, Davidsson J, Ng KC, et al: A
model for mild traumatic brain injury that induces limited
transient memory impairment and increased levels of axon related
serum biomarkers. Front Neurol. 3:1152012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singleton RH, Yan HQ, Fellows-Mayle W and
Dixon CE: Resveratrol attenuates behavioral impairments and reduces
cortical and hippocampal loss in a rat controlled cortical impact
model of traumatic brain injury. J Neurotrauma. 27:1091–1099. 2010.
View Article : Google Scholar
|
|
15
|
Alessandri B, Nishioka T, Heimann A,
Bullock RM and Kempski O: Caspase-dependent cell death involved in
brain damage after acute subdural hematoma in rats. Brain Res.
1111:196–202. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei XE, Wang D, Li MH, Zhang YZ, Li YH and
Li WB: A useful tool for the initial assessment of blood-brain
barrier permeability after traumatic brain injury in rabbits:
dynamic contrast-enhanced magnetic resonance imaging. J Trauma.
71:1645–1650. 2011. View Article : Google Scholar
|
|
17
|
Xing G, Ren M, O’Neill JT, et al: Pyruvate
dehydrogenase phosphatase1 mRNA expression is divergently and
dynamically regulated between rat cerebral cortex, hippocampus and
thalamus after traumatic brain injury: a potential biomarker of
TBI-induced hyper- and hypo-glycaemia and neuronal vulnerability.
Neurosci Lett. 525:140–145. 2012.
|
|
18
|
Fearing MA, Bigler ED, Wilde EA, et al:
Morphometric MRI findings in the thalamus and brainstem in children
after moderate to severe traumatic brain injury. J Child Neurol.
23:729–737. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Scheibel RS, Newsome MR, Troyanskaya M, et
al: Effects of severity of traumatic brain injury and brain reserve
on cognitive-control related brain activation. J Neurotrauma.
26:1447–1461. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Monea AG, Verpoest I, Vander Sloten J, Van
der Perre G, Goffin J and Depreitere B: Assessment of relative
brain-skull motion in quasistatic circumstances by magnetic
resonance imaging. J Neurotrauma. 29:2305–2317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ragatsky GG, Kamenir Y and Mayevsky A:
Effects of hyperbaric oxygenation on intracranial pressure
elevation rate in rats during the early phase of severe traumatic
brain injury. Brain Res. 1047:131–136. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rønning PA, Pedersen T, Skaga NO, Helseth
E, Langmoen IA and Stavem K: External validation of a prognostic
model for early mortality after traumatic brain injury. J Trauma.
70:E56–E61. 2011.PubMed/NCBI
|
|
23
|
Schoeler M, Loetscher PD, Rossaint R,
Fahlenkamp AV, et al: Dexmedetomidine is neuroprotive in an vitro
model for traumatic brain injury. BMC Neurol. 12:202012. View Article : Google Scholar : PubMed/NCBI
|