Introduction
Lipopolysaccharide (LPS), the major component of the
outer membrane of Gram-negative bacteria, also plays a key role in
the recognition and signaling responses that lead to the
elimination of invading pathogens. The immune system is important
for fighting bacterial infections and mediating deleterious host
reactions in animals and humans (1–2).
LPS-induced inflammation develops by the secretion
of various pro-inflammatory mediators, including tumor necrosis
factor-α (TNF-α), interleukin-1β (IL-1β), IL-6, cyclooxygenase-2
(COX-2), inducible nitric oxide synthase (iNOS) and prostaglandin
E2 (PGE2) (3). During infections,
IL-1β and TNF-α, which are the classic pro-inflammatory cytokines,
act first in the inflammation process. Nuclear factor (NF)-κB is
downstream of the signaling pathway activating IL-1β and TNF-α.
Recent studies have shown that NF-κB is central to the regulation
of a number of genes responsible for the generation of inflammatory
mediators, for example iNOS and COX-2 (4). The increased activation of NF-κB has
been observed in heart, brain, spleen and lung injuries following
LPS exposure (5,6).
Forsythia suspensa Vahl. (F. suspensa)
is a well-known Chinese herbal medicine that has been used as an
important source of medicine for pyrexia, inflammation, ulcers and
gonorrhea (7–9), based on its antioxidant,
antibacterial, antiviral, choleretic and antiemetic activity
(10–12). Studies have shown that
forsythiaside and forsyth from F. suspensa constitute the
major bioactive components of this plant (13,14).
Forsythiaside, a phenylethanoside, has been shown to exhibit
antibacterial, antioxidant and antiviral activity in vivo
and in vitro(15). A study
by Jiang et al(16) showed
that forsythiaside reduced serum levels of TNF-α and IL-6,
decreased the infiltration of leukocytes and reduced the
histopathological damage in a rat myocardial ischemia-reperfusion
(I/R) model. In addition, Forsythiaside has been demonstrated to
attenuate lipid peroxidation, decrease lipoprotein-induced
endothelin-1 secretion by endothelial cells and inhibit COX-2
activity (17–19). However, the effect of forsythiaside
on the inflammatory cytokine production induced by LPS in broiler
chickens has not been investigated. As well as the liver, spleen
and thymus, the bursa of Fabricius (BF) is a primary immune organ
and it is also a unique avian humoral immune organ (20). The present study aimed to
investigate the anti-inflammatory effect of forsythiaside by
examining changes in body temperature and levels of
pro-inflammatory cytokines, including IL-1β, IL-6 and TNF-α,
induced by LPS in the BFs of broiler chickens. Furthermore, NF-κB,
iNOS and COX-2 mRNA expression was examined to further investigate
the potential mechanisms involved in the effects of
forsythiaside.
Material and methods
Chemicals and reagents
Forsythiaside, with a purity of 98.0%, was obtained
from Chengdu Herbpurify Co., Ltd. (Chengdu, China) and
Escherichia coli LPS (L2880; serotype, O55:B5) was obtained
from Sigma-Aldrich (St. Louis, MO, USA). ELISA kits for TNF-α,
IL-1β and IL-6 were purchased from R&D Systems (Minneapolis,
MN, USA), while NO assay kits were obtained from the Nanjing
Jiancheng Bioengineering Institute (Nanjing, China). A BCA protein
assay kit was purchased from Wuhan Boster Bio-engineering Limited
Co. (Wuhan, China) and TRIzol reagent was obtained from Invitrogen
Life Technologies, (Carlsbad, CA, USA). Moloney murine leukemia
virus (M-MLV), RNase inhibitor, oligo-dT, deoxyribonucleotide
triphosphate (dNTP) and 5X buffer were purchased from Takara
Biotechnology (Dalian) Co., Ltd. (Dalian, China). A FastStart
Universal SYBR Green Master (Rox) was obtained from Roche
Diagnostics (Indianapolis, IN, USA).
Animals and treatment
One-day-old male Arbor Acres broiler birds were
obtained from a local hatchery and housed in starter batteries with
access to water and commercial feed ad libitum, in
accordance with NRC recommendations. At 15 days of age, 40 chickens
were randomly divided into four treatment groups, control, LPS and
LPS plus forsythiaside (30 or 60 mg/kg), with 10 chickens in each
group. In the LPS plus forsythiaside (30 or 60 mg/kg) groups, the
chickens were orally administered with forsythiaside at doses of 30
and 60 mg/kg body weight (BW), respectively, for seven days. At 21
days of age, the chickens in the LPS and the LPS plus forsythiaside
(30 or 60 mg/kg) groups were intravenously injected with LPS at 200
μg/kg BW, while the control group received an equal volume of
saline. The study was approved by the Northeast Agricultural
University, Harbin, China.
Determination of cloacal temperature
The cloacal temperature of each bird was measured
prior to and 3 h after injection of LPS using a thermocouple rectal
probe thermometer. In addition, the general behavioral changes of
these birds, including agility and feeding patterns, were also
observed following the treatments, prior to sacrifice. The chickens
were humanely euthanized by cervical dislocation and the BF was
collected from each animal. Each BF was frozen immediately with
liquid nitrogen and stored at −80°C until further analysis.
Sample collection
The isolated BFs were divided into two parts and one
part was weighed. Following this, 0.9% saline, measuring nine-fold
the weight of the BF tissue, (W:V=1:9) was added to a beaker. The
BFs were then minced, ground and centrifuged at 3,000 × g for 10
min. The extracted supernatant, representing a 10% tissue
suspension, was stored at −80°C until processing. The remaining
part of each BF was isolate RNA.
Measurement of NO levels
The concentration of NO in the BF tissues was
determined using an NO assay kit, according to the manufacturer’s
instructions. Briefly, the method involved measuring the levels of
NO metabolites, including nitrite and nitrate. Nitrate was reduced
first to nitrite by the action of nitrate reductase and the
reaction was then initiated by the addition of Griess reagent,
prior to the absorbance of the mixture at 550 nm being measured
(4).
Measurement of IL-1β, TNF-α and IL-6
levels
The tissue samples were centrifuged at 3,000 ×
g(Sigma-Aldrich, St. Louis, MO, USA), for 10 min at 4°C. Following
this, the cytokine concentrations of IL-1β, TNF-α and IL-6 in the
BFs were assayed using chicken ELISA kits, according to the
manufacturer’s instructions.
Measurement of IL-1β, TNF-α, IL-6, COX-2,
NF-κB and iNOS mRNA expression
RNA isolation and reverse
transcription
Total RNA was isolated using TRIzol reagent, in
accordance with the manufacturer’s instructions. Total RNA was
subsequently converted to cDNA using 8 μl oligo-dT primers and 8 μl
dNTP in 104 μl ddH2O at 70°C for 5 min, followed by 32
μl 5X buffer, 4 μl RNase inhibitor and 4 μl M-MLV at 42°C for 1 h.
The reaction was terminated by heating at 70°C for 15 min.
Quantitative polymerase chain reaction
(qPCR)
qPCR was performed using a LightCycler®
480 System (Roche Diagnostics) and the reactions were performed in
96-well plates (Roche Diagnostics) in a volume of 20 μl containing
10 μl LightCycler FastStart DNA Master SYBR Green I, 1.2 μl cDNA,
0.6 μl of each primer and 7.6 μl ddH2O. Standard cycling
conditions were used, including a pre-amplification step of 95°C
for 10 min, followed by amplification for 40 cycles of 95°C for 15
sec, 60°C for 1 min and 72°C for 20 sec. All the samples were
analyzed in triplicate. The mean cycle threshold (Ct) was
calculated for the target and house-keeping (β-actin) genes. The
amount of the target gene was normalized relative to that of the
housekeeping gene (ΔCt=Cttarget -
Cthousekeeping). The ΔΔCt value was calculated by
subtracting the ΔCt of the non-stimulated sample from the ΔCt of
the stimulated sample. The relative amount of the target gene in
the stimulated sample to that in the non-stimulated sample was
calculated by the 2−ΔΔCt method. The primers used are
shown in Table I.
 | Table IPrimer sequences for the real-time
polymerase chain reaction used in this study. |
Table I
Primer sequences for the real-time
polymerase chain reaction used in this study.
| Gene name | Gene bank accession
number | Primer sequence
(5′-3′) | Production length,
bp |
|---|
| TNF-α | GU230788.1 | Forward: GCC CTT CCT
GTA ACC AGAT G | 71 |
| | Reverse: ACA CGA CAG
CCA AGT CAA CG | |
| iNOS | NM_204961 | Forward: CCT GGA GGT
CCT GGA AGA GT | 82 |
| | Reverse: CCT GGG TTT
CAG AAG TGG C | |
| NF-κB p50 | M86930 | Forward: TCA ACG CAG
GAC CTA AAG ACA T | 162 |
| | Reverse: GCA GAT AGC
CAA GTT CAG GAT G | |
| COX-2 | NM_001167718.1 | Forward: TGT CCT TTC
ACT GCT TTC CAT | 84 |
| | Reverse: TTC CAT TGC
TGT GTT TGA GGT | |
| IL-6 | NM-204628 | Forward: AAA TCC CTC
CTC GCC AAT CT | 106 |
| | Reverse: CCC TCA CGG
TCT TCT CCA TAA A | |
| IL-1β | Y15006.1 | Forward: ACT GGG CAT
CAA GGG CTA CA | 142 |
| | Reverse: GCT GTC CAG
GCG GTA GAA GA | |
| β-actin | L08165 | Forward: CAC CAC AGC
CGA GAG AGA AAT | 135 |
| | Reverse: TGA CCA
TCA GGG AGT TCA TAG C | |
Statistical analysis
Quantitative data from the experiments are expressed
as the mean ± standard deviation. All groups were compared using a
one-way analysis of variance with SPSS 11.5 statistical software
(SPSS, Inc., Chicago, IL, USA) and an independent samples t-test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Clinical changes
Following LPS treatment, the chickens in the LPS
group showed symptoms of drowsiness and lethargy and exhibited
ruffled feathers and slight diarrhea within 3 h of injection. These
effects were not present in the control group, while in the LPS
plus forsythiaside (30 or 60 mg/kg) groups the symptoms were milder
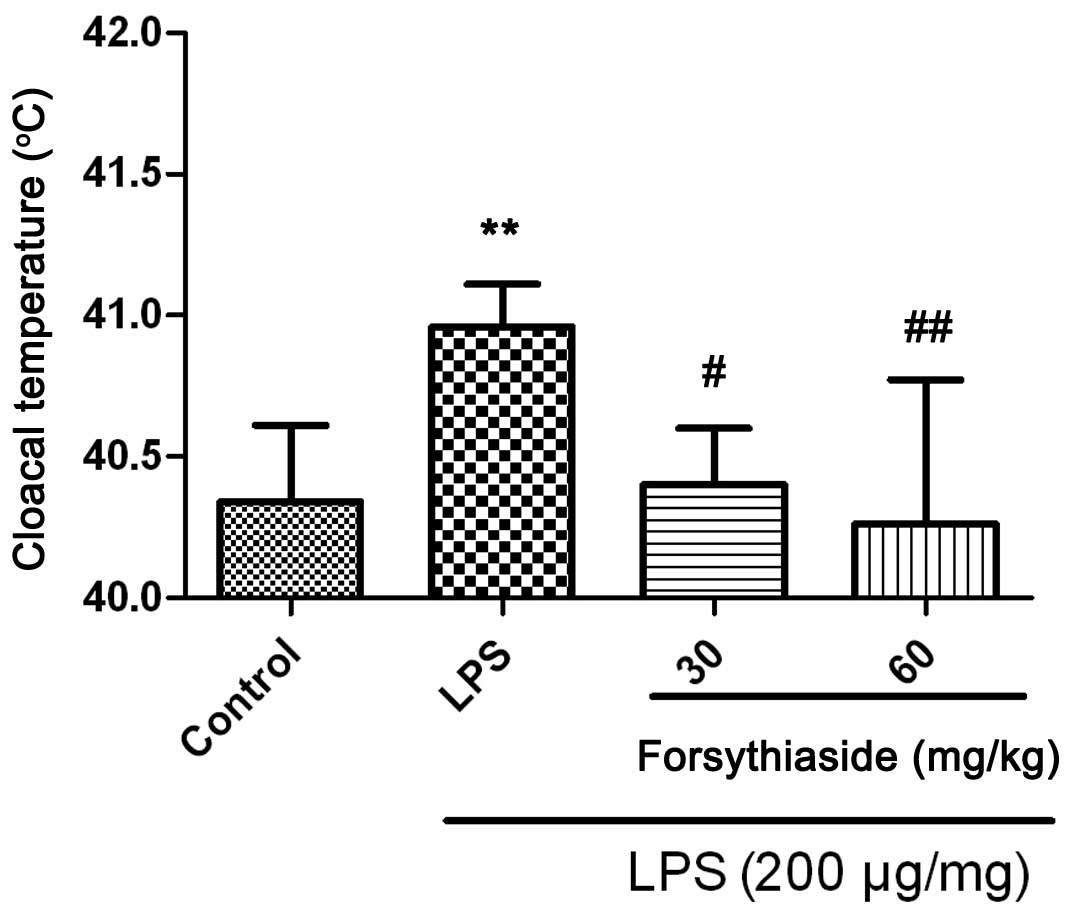
than those of the LPS group. In addition, the cloacal temperature
of the chickens in the LPS group was elevated at 3 h post
treatment, while the 30 or 60 mg/kg forsythiaside pretreatment for
seven days appeared to prevent the LPS-induced increase in cloacal
temperatures (Fig. 1).
Change in NO levels in the BFs of the
chickens
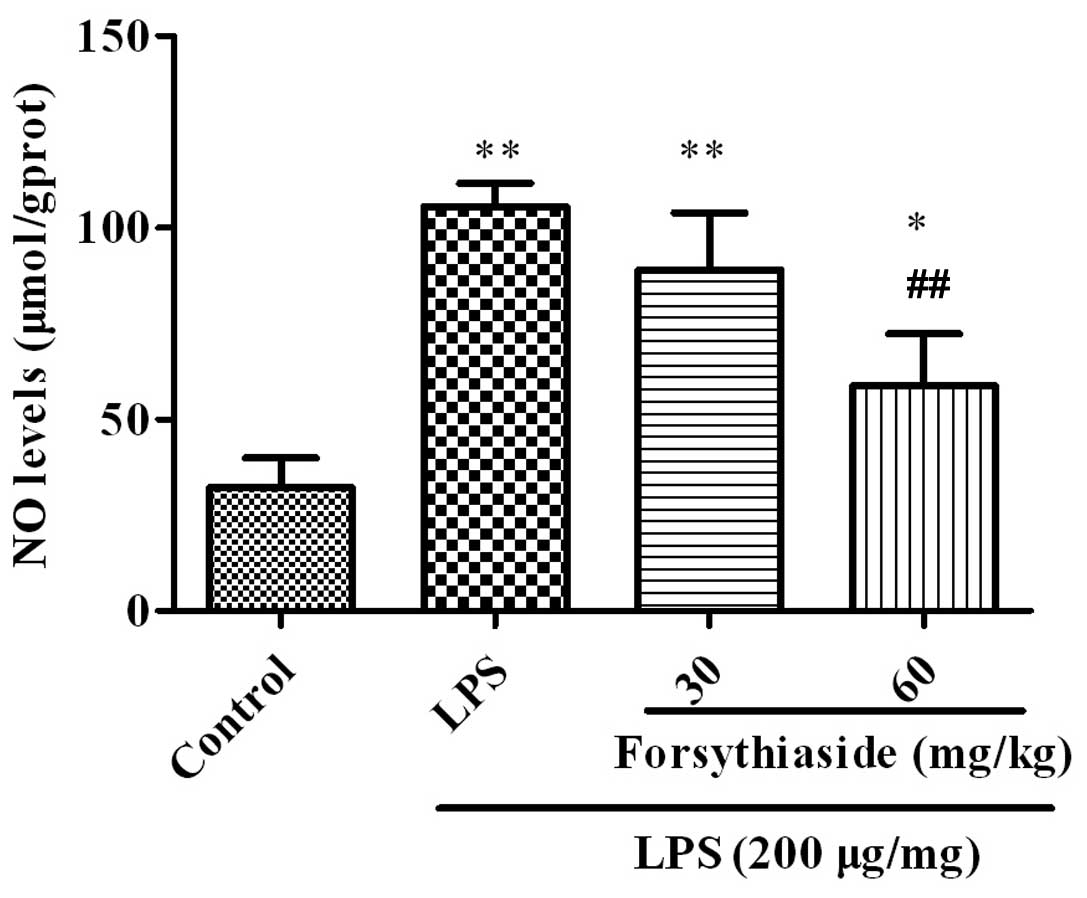
The NO levels were examined in the BFs of the
chickens and the results are shown in Fig. 2. In the LPS group, the NO level in
the BF was significantly increased to (105.5±6.2 μmol/g protein),
compared with the control group (P<0.01). When the chickens were
administered forsythiaside for seven days prior LPS injection,
i.e., in the LPS plus forsythiaside (30 or 60 mg/kg) groups, the NO
levels were significantly decreased to 89.2±14.9 and 58.7±136
μmol/g protein, respectively, compared with the LPS alone group
(P<0.01).
Changes in IL-1β, IL-6 and TNF-α levels
in the BFs of the chickens
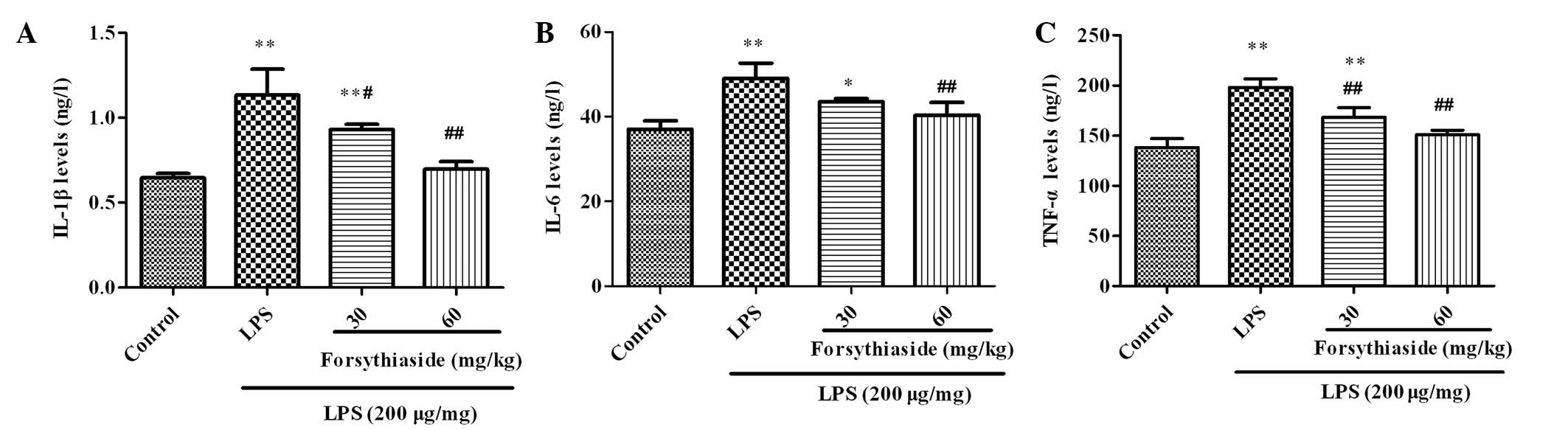
The concentrations of IL-1β, IL-6 and TNF-α in the
BF were examined using ELISA and the results are shown in Fig. 3. Three hours after LPS injection,
the levels of the cytokines, IL-1β, IL-6 and TNF-α, in the BF
homogenate were markedly increased compared with those in the
control group. As shown in Fig. 3,
pretreatment with forsythiaside (30 or 60 mg/kg) significantly
decreased the levels of IL-1β, IL-6 and TNF-α in a dose-dependent
manner.
Changes in IL-1β, TNF-α, IL-6, COX-2,
NF-κB and iNOS mRNA expression
The mRNA expression of IL-1β, TNF-α, IL-6, COX-2,
NF-κB and iNOS in the BF was examined and the results are shown in
Fig. 4. Three hours after LPS
injection, the mRNA expression of IL-1β, TNF-α, IL-6, COX-2, NF-κB
and iNOS in the BF homogenate of the LPS group were significantly
increased to 2.1±0.15-, 1.77±0.23-, 2.73±0.19-, 1.95±0.14-,
1.73±0.07- and 1.75±0.14-fold the expression levels of the control
group, respectively. However, pretreatment with forsythiaside (30
or 60 mg/kg) significantly decreased the levels of IL-1β, TNF-α,
IL-6, COX-2, NF-κB and iNOS mRNA expression compared with the LPS
alone group in a dose-dependent manner.
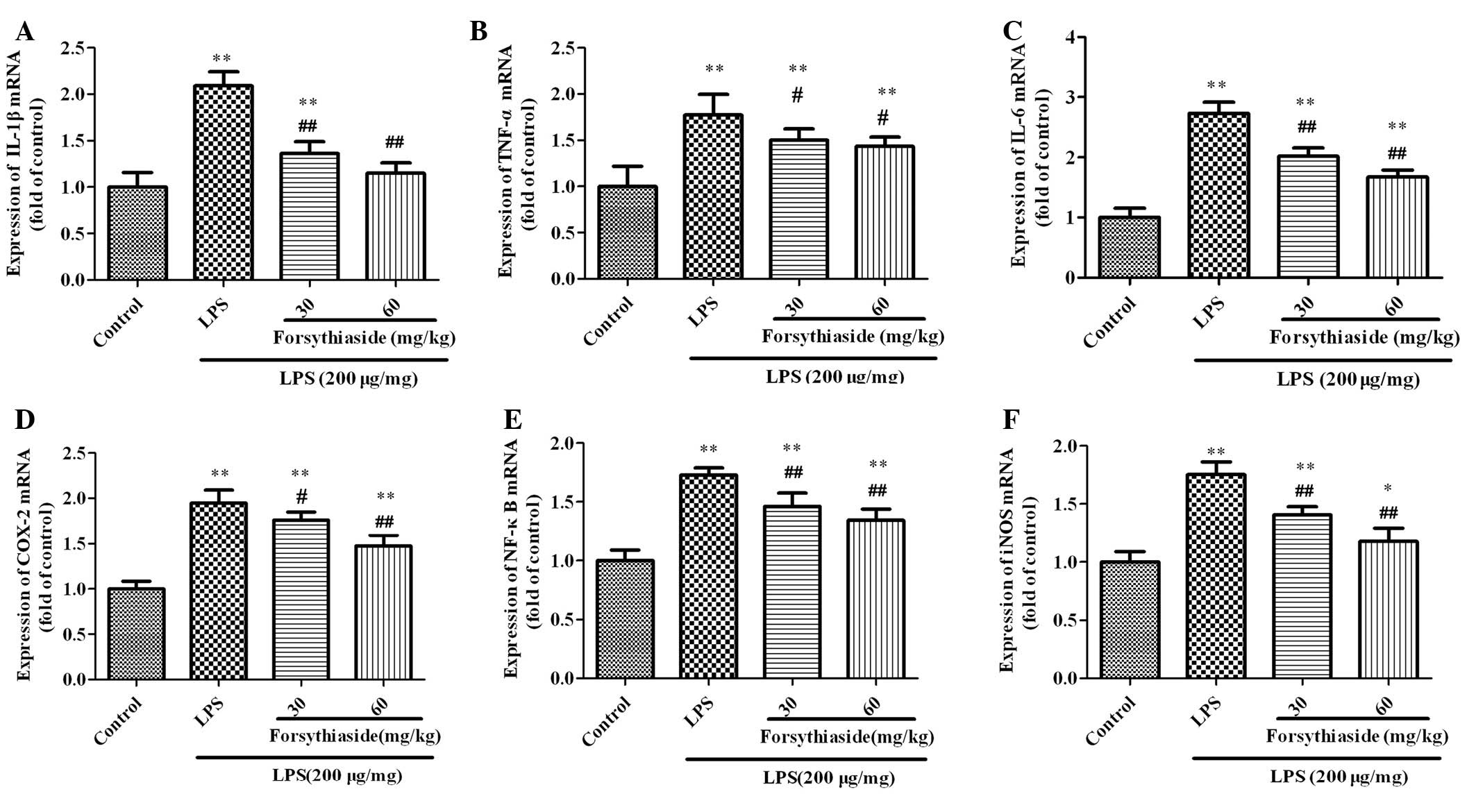 | Figure 4Effect of forsythiaside on the IL-1β,
TNF-α, IL-6, COX-2, NF-κB and iNOS mRNA expression in the bursa of
Fabricius of LPS-treated chickens: (A) IL-1β, (B) TNF-α, (C) IL-6,
(D) COX-2, (E) NF-κB and (F) iNOS. Data are expressed as the mean ±
standard deviation (n=10). *P<0.05 and
**P<0.01, vs. control group; #P<0.05
and ##P<0.01,vs. LPS-treated group. IL, interleukin;
TNF, tumor necrosis factor; LPS, lipopolysaccharide; COX-2,
cyclooxygenase-2; NF-κB, nuclear factor-κ; iNOS, inducible nitric
oxide; LPS, lipopolysaccharide. |
Discussion
In the present study, the effects of forsythiaside
on the acute-phase response to LPS-induced inflammation in the BFs
of broiler chickens were measured. Our results demonstrated that
forsythiaside exhibits a promising anti-inflammatory activity by
decreasing cloacal temperature and the manifestation of clinical
symptoms. In addition, these protective effects were found to
correlate with the attenuation of the inflammatory responses. The
in vitro anti-inflammatory effects of forsythiaside have
been reported in a previous study (13). However, to the best of our
knowledge, this study has demonstrated for the first time that
forsythiaside is able to protect against LPS-induced injury in the
BF of the chicken.
In the present study, when the chickens were
administered with LPS alone, the cloacal temperature of the
chickens was significantly increased, compared with the control
group, and specific abnormal symptoms were apparent. These
observations are consistent with previous studies (21–23).
However, in the chickens that were pretreated with forsythiaside,
these symptoms and increases in cloacal temperature were reduced.
IL-1β, IL-6 and TNF-α are the primary mediators of the acute-phase
response (21,24,25).
It is known that LPS stimulation leads to the production of the
pro-inflammatory cytokines, IL-1β, IL-6 and TNF-α, in chicken
organs, including the spleen, liver and BF (3,20,22,23).
Increases in the levels of these cytokines were observed 3 h after
intravenous injection of 200 μg/kg BW LPS and were reversed in
chickens pretreated with forsythiaside (30 or 60 mg/kg).
NO is a highly reactive free radical involved in a
number of physiological and pathological processes in the
inflammatory reaction (26). It is
produced by iNOS and reacts with superoxide to yield peroxynitrite,
particularly in immune cells. iNOS expression is associated with
the upregulation of NF-κB, and NF-κB sites identified in the iNOS
gene promoter region, which can be activated by LPS (1). In the present study, the levels of
NF-κB mRNA were significantly elevated at 3 h after injection of
LPS compared with the control group. The reduction in NO production
in the BFs treated with forsythiaside is likely to be relevant to
these observations and may be linked to alterations in the
signaling cascades triggered by iNOS expression. These results have
demonstrated that the anti-inflammatory effects of forsythiaside
may be mediated by the NF-κB-iNOS-NO signaling pathway. In
addition, the iNOS-NO signaling pathway may also have contributed
to the oxidative stress induced by LPS, which, in the LPS plus
forsythiaside group, was downregulated due to the antioxidative
effects of forsythiaside (13,27).
The NF-κB signaling pathway is regulated by a number
of different factors or signaling pathways, including IL-1β, TNF-α,
caspase-3, reactive oxygen species p38, c-Jun N-terminal kinases
and extracellular signal-regulated kinases/mitogen-activated
protein kinases (28,29). Inflammation and oxidative stress
are mutual influences in specific diseases and NF-κB may be pivotal
to this relationship (30). The
activation of NF-κB increases the expression of specific
inflammatory factors, including COX-2, IL-8 and TNF-α (29). In the present study, there was a
marked inhibition of IL-6, IL-1β, TNF-α and COX-2 secretion in the
BFs of chickens that were pretreated with forsythiaside, which may
be attributable to the effects of forsythiaside on NF-κB action
(31). Jiang et al(16) revealed that forsythiaside B
decreased inflammatory mediators, including NF-κB, TNF-α and IL-6,
in a rat myocardial I/R injury model.
In conclusion, results of the current study indicate
that forsythiaside reduces LPS-induced injury in the BFs of
chickens, due to its anti-inflammatory function. The mechanisms by
which forsythiaside exerts its anti-inflammatory effect correlate
with the inhibition of IL-6, IL-1β, TNF-α and COX-2 production, via
the inactivation of NF-κB. In addition, the NF-κB-iNOS-NO signaling
pathway may be important in this process. This study provide
further insight into the anti-inflammatory mechanisms of
forsythiaside.
Acknowledgements
This study was supported by a grants from the
National Science and Technology Supporting Projects (no.
2011BAD34B01-03) operated by the Ministry of Science and Technology
of China.
References
|
1
|
Shao DZ and Lin M: Platonin inhibits
LPS-induced NF-kappaB by preventing activation of Akt and IKKbeta
in human PBMC. Inflamm Res. 57:601–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shen YB, Piao XS, Kim SW, et al: The
effects of berberine on the magnitude of the acute inflammatory
response induced by Escherichia coli lipopolysaccharide in
broiler chickens. Poult Sci. 89:13–19. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bhatia M and Moochhala S: Role of
inflammatory mediators in the pathophysiology of acute respiratory
distress syndrome. J Pathol. 202:145–56. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen X, Yang X, Liu T, Guan M, et al:
Kaempferol regulates MAPKs and NF-κB signaling pathways to
attenuate LPS-induced acute lung injury in mice. Int
Immunopharmacol. 14:209–216. 2012.PubMed/NCBI
|
|
5
|
Mallard C: Innate immune regulation by
toll-like receptors in the brain. ISRN Neurol. 2012:7019502012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011.
|
|
7
|
Li HB and Chen F: Preparative isolation
and purification of phillyrin from the medicinal plant Forsythia
suspensa by high-speed counter-current chromatography. J
Chromatogr A. 1083:102–105. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee JY, Cho BJ, Park TW, et al:
Dibenzylbutyrolactone lignans from Forsythia koreana fruits
attenuate lipopolysaccharide-induced inducible nitric oxide
synthetase and cyclooxygenase-2 expressions through activation of
nuclear factor-κb and mitogen-activated protein kinase in RAW264.7
cells. Biol Pharm Bull. 33:1847–1853. 2010.
|
|
9
|
Sheng Z, Li JC and Li YH: Optimization of
forsythoside extraction from Forsythia suspensa by
Box-Behnken design. Afr J Biotechnol. 10:11728–11737. 2011.
|
|
10
|
Kinoshita K, Kawai T, Imaizumi T, et al:
Anti-emetic principles of Inula linariaefolia flowers and
Forsythia suspensa fruits. Phytomedicine. 3:51–58. 1996.
|
|
11
|
Wang L, Piao XL, Kim SW, et al: Effects of
Forsythia suspensa extract on growth performance, nutrient
digestibility, and antioxidant activities in broiler chickens under
high ambient temperature. Poult Sci. 87:1287–1294. 2008.
|
|
12
|
Li YH, Li MY, Cui L, et al: The effects of
ethanol extracts from Forsythia suspensa against
antibiotic-resistant Streptococcus suis isolates in vivo and
in vitro. In: Int Conf Bioinform Biomed Eng: 5th International
Conference; pp. 1–5. 2011
|
|
13
|
Qu H, Zhang Y, Wang Y, et al: Antioxidant
and antibacterial activity of two compounds (forsythiaside and
forsythin) isolated from Forsythia suspensa. J Pharm
Pharmacol. 60:261–266. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J and Zhang FX: Studies on the
antibiotic and antioxidant activities of weeping forsythia applied
in Chinese-style sausage. Chin Agric Sci Bull. 4:112–115. 2006.(In
Chinese).
|
|
15
|
Liu WB, Li DP, Zhang GL, et al: Study
progress of the pharmacological activity of Forsythoside A.
Zhongguo Xu Mu Shou Yi. 7:236–238. 2011.(In Chinese).
|
|
16
|
Jiang WL, Fu FH, Xu BM, et al:
Cardioprotection with forsythoside B in rat myocardial
ischemia-reperfusion injury: relation to inflammation response.
Phytomedicine. 17:635–639. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sahpaz S, Garbacki N, Tits M and Bailleil
F: Isolation and pharmacological activity of phenylpropanoid esters
from Marrubium vulgare. J Ethnopharmacol. 79:389–3892. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martin-Nizard F, Sahpaz S, Furman C, et
al: Natural phenylpropanoids protect endothelial cells against
oxidized LDL-induced cytotoxicity. Planta Med. 69:207–211. 2003.
View Article : Google Scholar
|
|
19
|
Martin-Nizard F, Sahpaz S, Kandoussi A, et
al: Natural phenylpropanoids inhibit lipoprotein-induced
endothelin-1 secretion by endothelial cells. J Pharm Pharmacol.
56:1607–1611. 2004. View Article : Google Scholar
|
|
20
|
Koutsos EA, García López JC and Klasing
KC: Carotenoids from in ovo or dietary sources blunt systemic
indices of the inflammatory response in growing chicks (Gallus
gallus domesticus). J Nutr. 136:1027–1231. 2006.PubMed/NCBI
|
|
21
|
Xie H, Rath NC, Huff GR, et al: Effects of
Salmonella typhimurium lipopolysaccharide on broiler
chickens. Poult Sci. 79:33–40. 2000.
|
|
22
|
Meriwether LS, Humphrey BD, Peterson DG,
et al: Lutein exposure, in ovo or in the diet, reduces parameters
of inflammation in the liver and spleen laying-type chicks
(Gallus gallus domesticus). J Anim Physiol Anim Nutr (Berl).
94:e115–e122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shanmugasundaram R and Selvaraj RK: Lutein
supplementation alters inflammatory cytokine production and
antioxidant status in F-line turkeys. Poult Sci. 90:971–976. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MacKay RJ and Lester GD: Induction of the
acute-phase cytokine, hepatocyte-stimulating factor/interleukin 6,
in the circulation of horses treated with endotoxin. Am J Vet Res.
53:1285–1289. 1992.PubMed/NCBI
|
|
25
|
Rath NC, Huff GR, Huff WE and Balog JM:
Factors regulating bone maturity and strength in poultry. Poult
Sci. 79:1024–1032. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoon HJ, Moon ME, Park HS, et al: Chitosan
oligosaccharide (COS) inhibits LPS-induced inflammatory effects in
RAW 264.7 macrophage cells. Biochem Biophys Res Commun.
358:954–959. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Korhonen R, Lahti A, Kankaanranta H and
Moilanen E: Nitric oxide production and signaling in inflammation.
Curr Drug Targets Inflamm Allergy. 4:471–479. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bonizzi G, Piette J, Schoonbroodt S, et
al: Reactive oxygen intermediate-dependent NF-kappaB activation by
interleukin-1beta requires 5-lipoxygenase or NADPH oxidase
activity. Mol Cell Biol. 19:1950–1960. 1999.PubMed/NCBI
|
|
29
|
Mendis E, Kim MM, Rajapakse N and Kim SK:
Suppression of cytokine production in lipopolysaccharide-stimulated
mouse macrophages by novel cationic glucosamine derivative involves
down-regulation of NF-kappaB and MAPK expressions. Bioorg Med Chem.
16:8390–8396. 2008. View Article : Google Scholar
|
|
30
|
McCabe C, Samali A and O’Brien T: Beta
cell cytoprotective strategies: establishing the relative roles for
iNOS and ROS. Biochem Biophys Res Commun. 342:1240–1248. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bengmark S: Curcumin, an atoxic
antioxidant and natural NFkappaB, cyclooxygenase-2, lipooxygenase,
and inducible nitric oxide synthase inhibitor: a shield against
acute and chronic diseases. JPEN J Parenter Enteral Nutr. 30:45–51.
2006. View Article : Google Scholar
|