Introduction
A number of studies have suggested that immune
factors are involved in the pathogenesis of epilepsy (1–3).
During seizures, the cellular immune function becomes abnormal,
which leads to changes in the intracorporeal cytokine levels. Among
these cytokines, interleukin-6 (IL-6), in particular, has been
implicated. IL-6 is generally synthesized by mononuclear
phagocytes, vascular endothelial cells and fibroblasts. It is also
produced by activated T cells. The main biological function of IL-6
is stimulating the maturation of activated B cells. In the nervous
system, neurons and glial cells (including microglia and
astrocytes) all secrete IL-6. The hippocampus, neocortex and
cerebella exhibit high expression levels of IL-6 and its receptor.
Similar to other cytokines, IL-6 binds to the α chain of the IL-6
receptor (IL-6R) on cell surfaces. Subsequently, the α chain and
the receptor subunit gp130 (IL-6Rβ chain) form a high-affinity
receptor that transmits a signal into cells through the
gp130/Jak/STAT route to regulate gene transcription (4). In general, IL-6 has a neuroprotective
effect on ischemic lesions, glutamate excitotoxicity and tumor
necrosis factor (TNF)-mediated injuries (5–7).
Furthermore, IL-6 has been shown to promote neuronal survival. In a
study by Thier et al(8), it
was revealed that the IL-6 secreted by sensory nerves and the IL-6
receptors jointly acted as neurotrophic factors for the maintenance
of neuronal survival. In addition, IL-6 has shown the ability to
induce PC12 cells to differentiate into nerve cells (9). IL-6 is able to protect striatal
neurons against N-methyl-D-aspartate (NMDA) toxicity
(8) and dopaminergic neurons
against 1-methyl-4-phenylpyridinium (MPP) toxicity (10). Moreover, IL-6 is able to promote
the regeneration of severed peripheral nerve cells (11). März et al(12) demonstrated that IL-6 was able to
induce cultured typical stellate cells to transform into
fibroblast-like cells, and to induce the expression of neurotrophic
factors. Moreover, in different brain regions, IL-6 induced
different neurotrophic factors, thus indicating that IL-6 was
involved in multiple central nervous system (CNS) functions
(11). It has been suggested that
IL-6 directly participates in the excitation of nerve cells.
Tancredi et al(13)
demonstrated that IL-6 inhibited the post-tetanic potentiation
(PTP) and the long-term potentiation (LTP) of nerve cells in the
hippocampal CA1 region, as well as participating in the plasticity
of the hippocampal synapse. Therefore, IL-6 is closely associated
with neurological excitability. Qiu et al(14) showed that increased IL-6 levels
were capable of notably changing the response to excitatory
neurotransmitter during CNS neuronal development, and that IL-6
pretreatment was able to change the level of influx of
extracellular calcium ions and the release of stored calcium, thus
selectively enhancing the response of intracellular calcium ions to
NMDA. In an experiment with mice, IL-6 was shown to be involved in
the response to cerebral injury (15). In addition, a number of studies
have demonstrated that IL-6 is able to influence the ethological
manifestations of animals (16–18).
IL-6 has been suggested to be associated with stress (19), as well as certain nervous and
mental illnesses, such as infantile autism (20) and migraine (21). Furthermore, IL-6 is closely
associated with neurological excitability; thus it has been
proposed that IL-6 may be involved in epilepsy. In 1999, Jankowsky
and Patterson (2) observed
increased levels of oncostatin M (OSM), leukemia inhibitory factor
(LIF), cardiotrophin-1 (CT-1), ciliary neurotrophic factor (cNTF)
and other cytokines in the hippocampi of pilocarpine-induced
epileptic rats, and proposed that cytokines were possibly
intercellular informational molecules in the mechanism underlying
the formation of epilepsy. D’Arcangle et al(22) later demonstrated that IL-6 was able
to reduce synapsin I phosphorylation, and transgenic mice with the
synapsin I deletion have been shown to be susceptible to
epileptiform seizures during growth (23). Therefore, IL-6 may be involved in
epileptic formation. Steffensen et al(24) observed that although IL-6 had a
certain protective effect on nerve cell damage under general
conditions, the continuously high expression of IL-6 in transgenic
mice increased neuron deficiency, gliosis and hippocampal seizure
discharge, inhibited the θ rhythm and participated in
epileptogenesis. Results from a number of studies (25–27)
have indicated that convulsions caused by viral infections may be
associated with IL-6. In a previous study, IL-6 was shown to
aggravate thermal shock in young mice (28). Moreover, in a study by Lehtimäki
et al(29), northern
blotting and in situ hybridization were used to assess the
effects of kainic acid on the levels of cytokines and their
corresponding receptors in the CNS of rats, and it was revealed
that IL-6 and IL-6R expression levels had increased. IL-6 has also
been suggested to be involved in the control of epilepsy, as well
as neuronal damage and protection. Kalueff et al(30) revealed that IL-6 was capable of
aggravating chemical-induced rat convulsions. Furthermore, in a
study by Peltola et al(31), it was observed in the clinic that,
following an epileptic attack, IL-6 levels in the cerebrospinal
fluid of patients appeared to increase. It was therefore proposed
that IL-1, IL-6 and TNF were regulators of colloid cells following
CNS damage. In addition, Lehtimäki et al observed that a
one-time epileptic attack was capable of increasing the IL-6
concentration in the blood of patients (32). A Japanese study showed that among
pediatric patients with influenza and neurological complications,
the IL-6 concentration in the blood of the convulsion group was
higher than that of the delirium group and the control group,
respectively (33). Therefore, the
interrelation between IL-6 and epilepsy requires further study. As
clinical research is limited in the actual condition of patients,
it is only possible to detect the expression of IL-6 in
cerebrospinal fluid or blood. Furthermore, the expression of IL-6
in living brain tissue is unclear. However, the majority of the
previous animal experiments have used a chemical-induced epileptic
rat model, with which it was difficult to avoid the impact of the
chemicals themselves on the toxicity and stimulatory effect of
IL-6. Therefore, the present study used the electrical kindling rat
model of epilepsy to explore the possible roles of IL-6 and
antiepileptic drugs in the incidence and treatment of epilepsy. By
observing the changes in the IL-6 mRNA expression level in the
hippocampi of electrically kindled epileptic rats, and the effect
of an antiepileptic drug on the rats, this methodology was able to
avoid the direct toxic action of chemical drugs and reduce the
interference of the chemicals, thus making any obtained conclusions
more reliable.
Topiramate (TPM) is a second-generation
antiepileptic drug that affects a variety of cellular mechanisms
(34), such as the inhibition of
voltage-dependent sodium channels and the negative modulation of
L-type calcium channels. TPM has been suggested to exhibit
neuroprotective and antiepileptic effects (35), and was therefore used in this
study.
Materials and methods
Animals and grouping
A total of 24 specific pathogen-free (SPF) male
Sprague-Dawley (SD) rats, with body weights ranging from 180 to 280
g, were obtained from the Experimental Animal Center of Soochow
University (License no. SYXK-Su-2002-0037; Suzhou, China). The rats
were randomly divided into four groups: blank control, surgical
control, epilepsy and TPM, with six rats in each group. The rats
were bred in separate cages away from intense light, under a 24-h
circadian cycle, and were allowed to freely eat and drink. This
study was performed in strict accordance with the recommendations
in the Guide for the Care and Use of Laboratory Animals of the
National Institutes of Health (8th edition, 2011). The animal use
protocol was reviewed and approved by the Institutional Animal Care
and Use Committee (IACUC) of Soochow University.
Preparation of electrically kindled
animal models
All rats, with the exception of those in the blank
control group, were anaesthetized with 4% (mass concentration)
chloral hydrate and fixed onto the brain stereotaxic apparatus. The
incisor was set to 2.4 mm below the ear rod. Based on the spectra,
the left basolateral amygdaloid nucleus of the rats was determined
(2.0 mm caudal to the bregma, 5.0 mm lateral to the midline and 8.0
mm ventral to the dura). A bipolar electrode with a diameter of
0.20 mm was inserted and fixed onto the surface of the skull. The
rats were left to recuperate for 1 week following the surgery, and
were then stimulated with 2-sec-long 50-Hz, 400-μA continuous
single-phase square waves with a width of 1 msec, once daily. For
the blank control and surgical control groups, no stimulation was
applied. Following Racines’s grading (36), the seizure intensities were
observed and recorded. If the rats presented a grade 5 reaction
five consecutive times, the kindling was deemed successful and
stimulation was immediately stopped.
Synchronous recording of a video
electroencephalogram
A Cadwell video electroencephalogram analysis system
(Cadwell Laboratories, Inc., Kennewick, WA, USA) was used to
synchronously record the background electroencephalogram of the
rats in the free state and the afterdischarge duration (ADD) during
electrical stimulation. Afterdischarge (AD) refers to a period of
epileptiform activity observed with at least five confirmable
high-amplitude, multiple-spiked waves that appear following
electrical stimulation. ADD is the duration from the end of
stimulation to the end of the AD.
Drug experiment and specimen
acquisition
Following successful kindling, 2 ml/day normal
saline was intragastrically administered to the rats in the blank
control, surgical control and epilepsy groups, whereas 85 mg/kg/day
TPM was intragastrically administered to the rats in the TPM group.
For all groups, the intragastric administration was conducted for
eight days. In the epilepsy and TPM groups, electrical stimulation
was conducted once prior to the intragastric administration,
according to the parameters required for kindling. The
characteristics of the seizures and the corresponding
electroencephalograms were recorded simultaneously. At the end of
the intragastric administration, the electrical stimulation was
repeated, and the resulting electroencephalograms were recorded.
Moreover, the characteristics of the seizure and the changes in the
electroencephalograms between the epilepsy and TPM groups were
compared. Twenty-four hours subsequent to the second electrical
stimulation, the rats were anaesthetized and the thoracic cavity
was opened. Following the puncturing of the aortic arch, the brain
of each rat was extracted and washed with phosphate-buffered saline
(PBS) and the right cerebral hippocampi were homogenized. The
homogenates were stored at −70°C for future use.
Reverse transcription-polymerase chain
reaction (RT-PCR)
The homogenized hippocampal tissues were treated
with TRIzol® reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) for cell lysis, and then the RNA was extracted
with chloroform, precipitated with isopropyl alcohol, eluted with
ethyl alcohol and dissolved in diethyl pyrocarbonate (DEPC)
solution. Following this, an ultraviolet spectrophotometer (752
Ultraviolet Spectrophotometer Model; Shanghai Precision Scientific
Instrument Corp., Shanghai, China) was used to measure the
absorbance at 260 nm (A260), and the RNA concentration
was calculated. Subsequently, reverse transcription was performed
and the cDNA product was stored at 4°C. The obtained cDNA was
amplified using PCR with the following primers: β-actin (expected
product size of 265 bp) forward primer (P1),
5′-AGGCCGGCTTCGCGGGCGAC-3′, and reverse primer (P2),
5′-CTCGGGAGCCACACGCAGCTC-3′; and IL-6 (expected product size of 442
bp) forward primer (P3), 5′-GAGAAAAGAGTTGTGCAATGGC-3′, and reverse
primer (P4), 5′-ACTAGGTTTGCCGAGTAGACC-3′. The PCR conditions were
as follows: initial denaturation at 94°C for 5 min, followed by 30
cycles of denaturation at 94°C for 30 sec, annealing at 56°C for 30
sec and extension at 72°C for 1 min, followed by a final extension
step at 72°C for 7 min. Subsequent to electrophoresis, the PCR
amplification products were analyzed using a gel imaging analysis
system. A BioCaptMW analysis system (BioCaptMW version 99.05 s
system; Vilber Courmat, Marine-La-Vallee, France) was used to
detect the gray value of each band, and β-actin was used as the
internal reference to semiquantitatively determine the IL-6 mRNA
expression levels.
Statistical analysis
The SPSS statistical software package (SPSS, Inc.,
Chicago, IL, USA) was used for all statistical analysis. The
experimental data are expressed as the mean ± standard deviation.
The means of the measurement data were compared among the multiple
groups using one-way analysis of variance (ANOVA), once the
homogeneity of the variance had been confirmed. The
Student-Newman-Keuls (SNK) method was used for comparisons between
groups. A Fisher’s test was used for the count data. P<0.05 was
considered to indicate a statistically significant difference.
Results
Kindling and electroencephalogram
records
With regard to the rat kindling process, the
shortest period required to achieve kindling was 7 days, while the
longest period was 19 days. The mean number of days was 13.50±3.99
days. In the epilepsy group, all the rats presented with grade 5
seizures. Following TPM treatment (the TPM group), the intensity,
amplitude and duration of the seizures were decreased, or the
seizures of the rats were completely controlled. Among the TPM
group, one rat exhibited grade 1 seizures, three rats exhibited
grade 3 seizures and two rats exhibited grade 4 seizures. None of
the rats exhibited grade 5 seizures in the TPM group. The two
groups were significantly different with regard to seizure grade
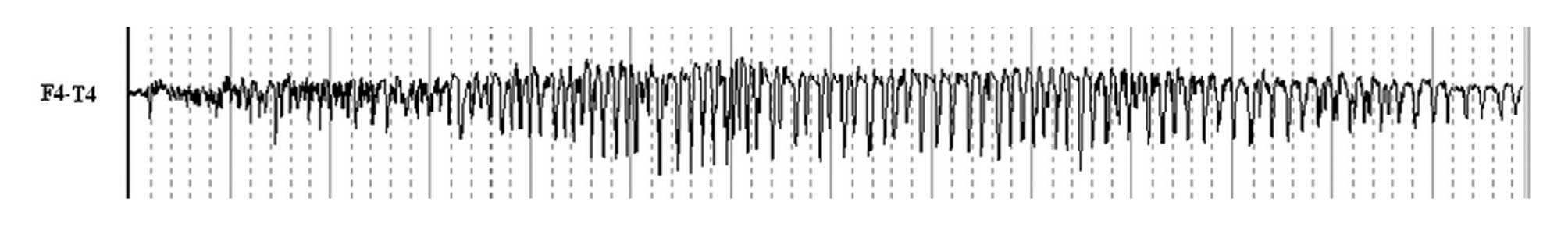
(P<0.05). At all recorded points, the ADD values of the
electrically kindled rats were between 21,450 and 119,720 msec
(Fig. 1). Moreover, the mean ADD
values of the epilepsy and TPM groups were 78,205.67±32,567.93 and
23,880.83±20,184.50 msec, respectively. The ADD of the TPM group
was significantly shorter than that of the epilepsy group
(P<0.05).
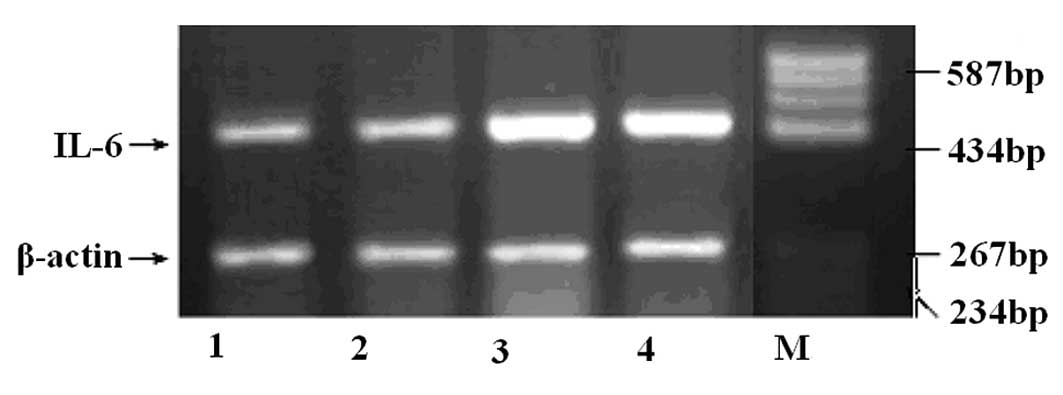
RT-PCR detection of IL-6
The variance analyses of the semiquantitative RT-PCR
results for IL-6 mRNA in the hippocampi of the rats in the four
treatment groups were significantly different (F=5.959, P<0.05).
As shown in Fig. 2, compared with
the levels in the two control groups (1.57±0.69 for the blank
control group and 1.80±0.54 for the surgical control group), the
IL-6 mRNA levels (mean 2.83±0.40) in the hippocampi of rats in the
epilepsy group were significantly increased (P<0.05). In
addition, the TPM group (2.18±0.56) had a different recovery rate;
however, the difference in IL-6 expression was not significant
between the TPM and epilepsy groups (P>0.05; Fig. 2).
Discussion
As a model for temporal lobe epilepsy, the kindling
model is widely used in studies of epilepsy, since the spontaneous
limbic system epilepsy caused by electrical kindling is similar to
temporal lobe epilepsy. In the CNS, a number of regions are able to
act as electrical stimulation sites for kindling rats. Among these
regions, the basolateral amygdaloid nucleus has the most extensive
conduction bundle link, and may be the focus of the epileptiform
manifestations in the limbic system (36). In the present study, a solid
electrode was inserted into the rats, and electroencephalograms
were recorded when the rats were in the free state. This surgical
method was convenient, and the detection results and the actual
values were reliable. Furthermore, compared with the
chemical-induced epilepsy model, the surgical method avoided the
interference of drug toxicity on the experimental data and improved
the reliability of the results.
The mechanisms that regulate IL-6 expression in the
hippocampus have yet to be elucidated. IL-1, interferon (IFN) and
TNF, as well as certain neurotransmitters, are capable of promoting
IL-6 secretion (37–39), and it was observed that following
LTP induction with an embedded electrode, the expression levels of
certain cytokines were notably increased. In particular, IL-6
levels were more significantly increased than those of the other
cytokines (39). In a study on the
changes in IL-6 expression following cerebral ischemia in rats, Ali
et al(40) demonstrated
that NMDA and ionomycin (a calcium ion carrier) upregulated IL-6
mRNA expression, which indicated that the NMDA receptor-mediated
calcium ion influx induced the neurons to generate IL-6. Therefore,
LTP and calcium ion influx induce neurons to generate IL-6. The
present study demonstrated that the level of IL-6 expression in the
hippocampi of electrically kindled rats was increased. One
potential mechanism for this is that the electrical kindling
increased the levels of excitatory neurotransmitters and decreased
the levels of γ-aminobutyric acid (GABA) in the neurons, thereby
increasing the influx of neuronal calcium ions, and, consequently,
neuronal IL-6 expression. Furthermore, the LTP induced by
electrical stimulation may also have been involved in promoting
IL-6 secretion.
The involvement of IL-6 with nervous system diseases
has been an area of increasing focus, and IL-6 is generally
recognized to exhibit neuroprotective effects (41). However, compared with wild-type
mice, the hippocampal seizure discharge rates of transgenic mice
with continuously high IL-6 expression have been shown to be
increased, whereas the θ rhythm of the transgenic mice was
inhibited; this continuously high IL-6 expression has been
indicated to be involved in seizure formation (24). In the present study, the IL-6
expression in the hippocampi of the electrically kindled rats was
increased. The preliminary increase in IL-6 expression may have
been a protective mechanism intended to preserve the neurons.
However, the continuously high expression may have been involved in
the kindling process by increasing gliosis and hippocampal seizure
discharge, among other mechanisms (25). Antiepileptic drugs decrease IL-6
expression by varying degrees, and may therefore be useful for
maintaining the appropriate IL-6 levels. TPM reduces the frequency
and intensity of seizures in epileptic rats, in addition to
reducing the cerebral injury caused by epilepsy or anoxia (42). In the present study, although TPM
was able to induce the recovery of the IL-6 levels, the difference
between the IL-6 levels in the TPM and epilepsy groups was not
significant. This phenomenon may have been due to the relatively
small sample size utilized, as well as insufficient control over
the experimental conditions in the study. Thus, this result remains
to be verified in future studies with larger sample sizes. Further
studies are also required to investigate the exact mechanisms
underlying the action of IL-6 in epilepsy.
Acknowledgements
The authors would like to thank Professor Yin
Qi-Zhang and Mrs. Gong Shan for their technical assistance in the
establishment of the model. This study was supported by grants from
the Project of Science and Technology Commission of Shanghai
Baoshan in China (no. 09-E-3) and from the ‘Medical-Engineering
(Science) Cross-Research Fund’ of Shanghai Jiaotong University (no.
YG2010MS16).
References
|
1
|
Jankowsky JL and Patterson PH: The role of
cytokines and growth factors in seizures and their sequelae. Prog
Neurobiol. 63:125–149. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jankowsky JL and Patterson TA:
Differential regulation of cytokine expression following
pilocarpine-induced seizure. Exp Neurol. 159:333–346. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maroso M, Balosso S, Ravizza T, et al:
Toll-like receptor 4 and high-mobility group box-1 are involved in
ictogenesis and can be targeted to reduce seizures. Nat Med.
16:413–419. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang Y and Fuller GM: Phosphorylation and
internalization of gp130 occur after IL-6 activation of Jak2 kinase
in hepatocytes. Mol Biol Cell. 5:819–828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heinrich PC, Behrmann I, Müller-Newen G,
Schaper F and Graeve L: Interleukin-6-type cytokine signalling
through the gp130/Jak/STAT pathway. Biochem J. 334:297–314.
1998.PubMed/NCBI
|
|
6
|
Funk JA, Gohlke J, Kraft AD, McPherson CA,
Collins JB and Jean HG: Voluntary exercise protects hippocampal
neurons from trimethyltin injury: possible role of interleukin-6 to
modulate tumor necrosis factor receptor-mediated neurotoxicity.
Brain Behav Immun. 25:1063–1077. 2011. View Article : Google Scholar
|
|
7
|
Liu Z, Qiu YH, Li B, Ma SH and Peng YP:
Neuroprotection of interleukin-6 against NMDA-induced apoptosis and
its signal-transduction mechanisms. Neurotox Res. 19:484–495. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Thier M, März P, Otten U, Weis J and
Rose-John S: Interleukin-6 (IL-6) and its soluble receptor support
survival of sensory neurons. J Neurosci Res. 55:411–422. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sterneck E, Kaplan DR and Johnson PF:
Interleukin-6 induces expression of peripherin and cooperates with
Trk receptor signaling to promote neuronal differentiation in PC12
cells. J Neurochem. 67:1365–1374. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Akaneya Y, Takahashi M and Hatanaka H:
Interleukin-1 beta enhances survival and interleukin-6 protects
against MPP+ neurotoxicity in cultures of fetal rat dopaminergic
neurons. Exp Neurol. 136:44–52. 1995.PubMed/NCBI
|
|
11
|
Toulmond S, Vige X, Fage D and Benavides
J: Local infusion of interleukin-6 attenuates the neurotoxic
effects of NMDA on rat striatal cholinergic neurons. Neurosci Lett.
144:49–52. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
März P, Heese K, Dimitriades-Schmutz B,
Rose-John S and Otten U: Role of interleukin-6 and soluble IL-6
receptor in region-specific induction of astrocytic differentiation
and neurotrophin expression. Glia. 26:191–200. 1999.PubMed/NCBI
|
|
13
|
Tancredi V, D’Antuono M, Cafè C, et al:
The inhibitory effects of interleukin-6 on synaptic plasticity in
the rat hippocampus are associated with an inhibition of
mitogen-activated protein kinase ERK. J Neurochem. 75:634–643.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu Z, Parsons KL and Gruol DL:
Interleukin-6 selectively enhances the intracellular calcium
response to NMDA in developing CNS neurons. J Neurosci.
15:6688–6699. 1995.PubMed/NCBI
|
|
15
|
Quarta S, Vogl C, Constantin CE, Üçeyler
N, Sommer C and Kress M: Genetic evidence for an essential role of
neuronally expressed IL-6 signal transducer gp130 in the induction
and maintenance of experimentally induced mechanical
hypersensitivity in vivo and in vitro. Mol Pain. 7:732011.
View Article : Google Scholar
|
|
16
|
Lee RH, Mills EA, Schwartz N, et al:
Neurodevelopmental effects of chronic exposure to elevated levels
of pro-inflammatory cytokines in a developing visual system. Neural
Dev. 5:22010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Burton MD, Sparkman NL and Johnson RW:
Inhibition of interleukin-6 trans-signaling in the brain
facilitates recovery from lipopolysaccharide-induced sickness
behavior. J Neuroinflammation. 8:542011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Munster BC, Aronica E, Zwinderman AH,
Eikelenboom P, Cunningham C and Rooij SE: Neuroinflammation in
delirium: a postmortem case-control study. Rejuvenation Res.
14:615–622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jankord R, Zhang R, Flak JN, Solomon MB,
Albertz J and Herman JP: Stress activation of IL-6 neurons in the
hypothalamus. Am J Physiol Regul Integr Comp Physiol.
299:R343–R351. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wei H, Zou H, Sheikh AM, et al: IL-6 is
increased in the cerebellum of autistic brain and alters neural
cell adhesion, migration and synaptic formation. J
Neuroinflammation. 8:522011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Burstein R and Levy D: Local
action of the proinflammatory cytokines IL-1β and IL-6 on
intracranial meningeal nociceptors. Cephalalgia. 32:66–72.
2012.
|
|
22
|
D’Arcangelo G, Tancredi V, Onofri F,
D’Antuono M, Giovedì S and Benfenati F: Interleukin-6 inhibits
neurotransmitter release and the spread of excitation in the rat
cerebral cortex. Eur J Neurosci. 12:1241–52. 2000.PubMed/NCBI
|
|
23
|
Terada S, Tsujimoto T, Takei Y, et al:
Inhibitory synaptic transmission in mice lacking synapsin I.
Neurosci Res. 28(Suppl 1): S571997. View Article : Google Scholar
|
|
24
|
Steffensen SC, Campbell IL and Henriksen
SJ: Site-specific hippocampal pathophysiology due to cerebral
overexpression of interleukin-6 in transgenic mice. Brain Res.
652:149–153. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kirkman NJ, Libbey JE, Wilcox KS, White HS
and Fujinami RS: Innate but not adaptive immune responses
contribute to behavioral seizures following viral infection.
Epilepsia. 51:454–464. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Libbey JE, Kennett NJ, Wilcox KS, White HS
and Fujinami RS: Interleukin-6, produced by resident cells of the
central nervous system and infiltrating cells, contributes to the
development of seizures following viral infection. J Virol.
85:6913–6922. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Libbey JE, Kennett NJ, Wilcox KS, White HS
and Fujinami RS: Once initiated, viral encephalitis-induced
seizures are consistent no matter the treatment or lack of
interleukin-6. J Neurovirol. 17:496–499. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fukuda M, Morimoto T, Suzuki Y, Shinonaga
C and Ishida Y: Interleukin-6 attenuates hyperthermia- induced
seizures in developing rats. Brain Dev. 29:644–648. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lehtimäki KA, Peltola J, Koskikallio E,
Keränen T and Honkaniemi J: Expression of cytokines and cytokine
receptors in the rat brain after kainic acid-induced seizures.
Brain Res Mol Brain Res. 110:253–260. 2003.PubMed/NCBI
|
|
30
|
Kalueff AV, Lehtimaki KA, Ylinen A,
Honkaniemi J and Peltola J: Intranasal administration of human IL-6
increases the severity of chemically induced seizures in rats.
Neurosci Lett. 365:106–110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peltola J, Hurme M, Miettinen A and
Keränen T: Elevated levels of interleukin-6 may occur in
cerebrospinal fluid from patients with recent epileptic seizures.
Epilepsy Res. 31:129–133. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lehtimäki KA, Keränen T, Palmio J, et al:
Increased plasma levels of cytokines after seizures in
localization-related epilepsy. Acta Neurol Scand. 116:226–230.
2007.PubMed/NCBI
|
|
33
|
Fukumoto Y, Okumura A, Hayakawa F, et al:
Serum levels of cytokines and EEG findings in children with
influenza associated with mild neurological complications. Brain
Dev. 29:425–430. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ben-Menachem E, Sander JW, Stefan H,
Schwalen S and Schauble B: Topiramate monotherapy in the treatment
of newly or recently diagnosed epilepsy. Clin Ther. 30:1180–1195.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Edmonds HL Jr, Jiang YD, Zhang PY and
Shank R: Topiramate as a neuroprotectant in a rat model of global
ischemia-induced neurodegeneration. Life Sci. 69:2265–2277. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Castillo CG, Mendoza S, Freed WJ and
Giordano M: Intranigral transplants of immortalized GABAergic cells
decrease the expression of kainic acid-induced seizures in the rat.
Behav Brain Res. 171:109–115. 2006. View Article : Google Scholar
|
|
37
|
Gadient RA and Otten UH: Interleukin-6
(IL-6) - a molecule with both beneficial and destuctive potentials.
Prog Neurobiol. 52:379–390. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pitkänen A, Savander V and LeDoux JE:
Organization of intra-amygdaloid circuitries in the rat: an
emerging framework for understanding functions of the amygdala.
Trends Neurosci. 20:517–523. 1997.PubMed/NCBI
|
|
39
|
Jankowsky JL, Derrick BE and Patterson PH:
Cytokine responses to LTP induction in the rat hippocampus: a
comparison of in vitro and in vivo techniques. Learn Mem.
7:400–412. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ali C, Nicole O, Docagne F, et al:
Ischemia-induced interleukin-6 as a potential endogenous
neuroprotective cytokine against NMDA receptor-mediated
excitotoxicity in the brain. J Cereb Blood Flow Metab. 20:956–966.
2000. View Article : Google Scholar
|
|
41
|
Calkavur S, Akisu M, Olukman O, et al:
Genetic factors that influence short-term neurodevelopmental
outcome in term hypoxic-ischaemic encephalopathic neonates. J Int
Med Res. 39:1744–1756. 2011. View Article : Google Scholar
|
|
42
|
Pitkänen A: Efficacy of current
antiepileptics to prevent neurodegeneration in epilepsy models.
Epilepsy Res. 50:141–160. 2002.PubMed/NCBI
|