Introduction
Systemic lupus erythematosus (SLE) is a chronic
relapsing autoimmune disease characterized by the production of
auto-antibodies directed against a range of cellular components and
its pathogenesis remains unclear. Genetic, hormonal, environmental
and immunological factors are involved in the pathogenesis of SLE
(1,2).
It is known that epigenetic modifications, including
DNA methylation and histone modifications, have a significant
impact on gene expression and abnormal decreases in T-cell DNA
methylation have been implicated in the development of drug-induced
and idiopathic lupus (3,4). The decreased T-cell DNA methylation
results in the overexpression of methylation-sensitive genes, such
as CD11a (LFA-1), perforin and CD70, in the T cells and this leads
to T-cell autoreactivity in vitro and autoimmunity in
vivo (5–9).
The methylation status of DNA is associated with
three types of enzymes, which have effects on maintenance
methylation, de novo methylation and demethylation (10,11).
DNA cytosine-5-methyl transferase 1 (DNMT1) is involved in
maintaining methylation, while methyl CpG binding domain protein 2
(MBD2) is associated with demethylation (12). The methylation-related molecules,
DNMT1 and MBD2, may also be associated with the development of
SLE.
Environmental effects on lupus may have a role in
mediating epigenetic changes in immunity. Exposure to ultraviolet
(UV) light has long been associated with the exacerbation of SLE
and photosensitivity remains a diagnostic criterion for this
disease (13), with up to 73% of
patients with SLE reporting photosensitivity (14). Most, but not all, cutaneous lupus
lesions occur in light-exposed areas and may be triggered by
sunlight exposure. Sunlight exposure itself is able to induce
systemic disease activity (15).
UV irradiation, particularly ultraviolet B light
(UVB, 290–320 nm), may induce systemic disease activity. However,
it remains unclear whether UVB affects SLE by altering DNA
methylation. Thus, the elucidation of the mechanisms of the effects
of UVB on DNA hypomethylation in SLE patients may provide clues to
the pathogenesis of the SLE. To investigate these mechanisms, we
analyzed the DNA methylation status and gene expression of DNMT1
and MBD2 in T cells from SLE patients and healthy controls
following treatment with different dosages of UVB irradiation. The
present study reports the findings in this field.
Materials and methods
Human subjects
Patients with SLE (n=35) were recruited from the
outpatient and inpatient services at the Huashan Hospital, Fudan
University (Shanghai, China). They included 31 females and 4 males
(mean age: 33.4 years; range, 18–51 years). A total of 21 gender-
and age-matched healthy volunteers served as the controls (18
females and 3 males; mean age 32.7 years; range, 19–51 years). This
study was approved by the institutional review board of the Huashan
Hospital (IRB: KY2009-054) and written informed consent was
obtained from all participants. All patients with SLE fulfilled at
least four of the criteria of the American Rheumatism Association
for the classification of SLE (13) and disease activity was assessed
using the SLE Disease Activity Index (SLEDAI) (16). Active disease was defined as an
SLEDAI score ≥5. In the 35 patients, 16 had the active disease,
while the other 19 were stable.
Peripheral blood mononuclear cells and
T-cell isolation
Peripheral blood mononuclear cells were isolated
using density gradient centrifugation. T cells were isolated by
CD3+ magnetic beads according to the manufacturer’s
instructions (T cell isolation kit; Miltenyi Biotec, Bergisch
Gladbach, Germany).
T-cell culture and UVB irradiation
CD3+ T cells were cultured in RPMI-1640
medium supplemented with 10% heat-inactivated fetal bovine serum
(FBS), 2 mM sodium pyruvate, 100 IU/ml penicillin and 100
μg/ml streptomycin.
UVB irradiation was performed using Waldman UV236B
lights (Waldmann Lighting Ltd., Wheeling, IL, USA), which emit the
majority of their energy within the UVB range (290–320 nm) with an
emission peak at 311 nm. CD3+ T cells were irradiated in
PBS with different doses of UVB (0, 50 and 100 mJ/cm2).
PBS was removed following irradiation and RPMI-1640 containing 10%
FBS was then added and the cells were cultured for 24 h. The doses
of UVB were selected based on the World Health Organisation
guidelines for sun exposure (17)
and on the standard erythemal dose (SED), a cumulative measure of
erythemal or sunburning solar UV irradiation (18). Following culturing, CD3+
T cells were harvested for the analyses of DNA methylation and mRNA
levels.
Flow cytometric analysis of
5-methylcytosine staining
Global DNA methylation was evaluated by staining the
cells with a specific monoclonal antibody against 5-methylcytidine
using previously published methods (19,20).
The T cells were washed with phosphate-buffered saline (PBS)
supplemented with 0.1% Tween-20 and 1% bovine serum albumin
(PBST-BSA), fixed with 0.25% paraformaldehyde at 37°C for 10 min
and 88% methanol at −20°C for at least 30 min. After two washes
with PBST-BSA, the cells were treated with 2 M HCl at 37°C for 30
min and then neutralized with 0.1 M sodium borate (pH 8.5). The
cells were blocked with 10% goat serum in PBST-BSA for 20 min at
37°C and incubated with anti-5-methylcytidine antibody (1
μg/ml) for 45 min at 37°C, followed by staining with goat
anti-mouse IgG conjugated with fluorescein isothiocyanate (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Finally, the cells
were washed three times with PBS and resuspended in PBS for
analysis by flow cytometry.
DNMT1 and MBD2 real-time reverse
transcription-polymerase chain reaction (RT-PCR)
Real-time quantitative RT-PCR was performed using an
iCycler IQ5 (Bio-Rad, Hercules, CA, USA) and previously published
methods (21). Total cellular RNA
was isolated from T cells using the TRIzol reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA). The RNA extracts were resuspended
in 15 μl RNase-free water and the RNA concentration was
adjusted to ∼1 μg/μl and used for cDNA synthesis. The
final reaction volume was 25 μl consisting of 12.5 μl
Maxima SYBR-Green master mix (Fermentas, Thermo Scientific,
Waltham, MA, USA), 0.5 μl each specific primer (10
μM), 1 μl cDNA template and 10.5 μl
nuclease-free H2O. The cycling conditions were 5 min at
94°C; then 40 cycles of 30 sec at 94°C, 30 sec at 55°C and 30 sec
at 72°C. The DNMT1 and MBD2 mRNA levels were quantified relative to
β-actin transcripts. The following primers were used: DNMT1
forward: 5′-GATTTGTCCTTGGAG AACGGTG-3′ and reverse:
5′-TGAGATGTGATGGTGGTT TGCC-3′; MBD2 forward: 5′-AGGTAGCAATGATGA
GACCCTTTTA-3′ and reverse: 5′-TAAGCCAAACAGCAG GGTTCTT-3′; β-actin
forward: 5′-GCACCACACCTT CTACAATGAGC-3′ and reverse:
5′-GGATAGCACAGCCTG GATAGC AAC-3′.
Statistical analysis
The data were analyzed using the Student’s t-test,
regression analysis or analysis of variance (ANOVA) as appropriate,
calculated with Stata 7.0 software. P<0.05 was condsidered to
indicate a statistically significant result.
Results
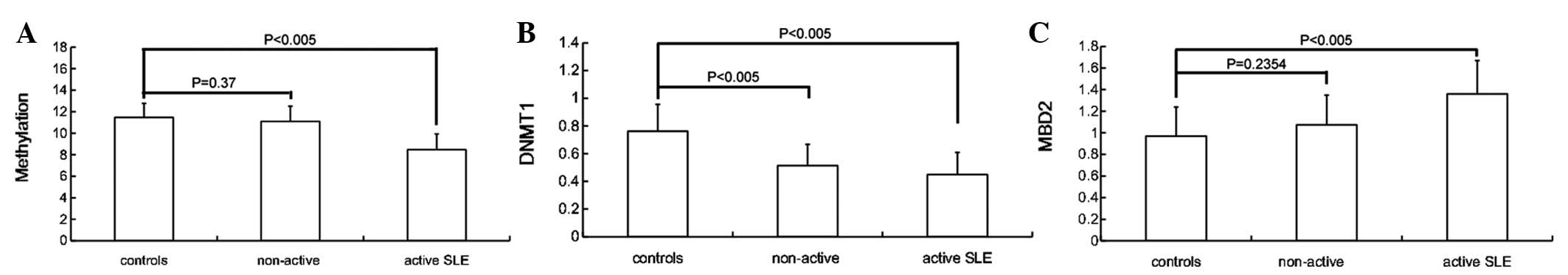
Patterns of DNA hypomethylation in SLE
patients
To determine whether global DNA methylation was
decreased in SLE patients, T cells were isolated from 35 patients
and 21 normal controls. DNA hypomethylation was observed in the
patients with SLE, whose DNA methylation level was 9.892±1.939
compared with 11.479±1.291 for the controls, and the difference was
statistically significant (P= 0.0016). Statistically significant
differences in DNA methylation were observed between the active
(SLEDAI ≥5) and non-active patients (SLEDAI <5; 8.469±1.458 vs.
11.090±1.418, respectively; P<0.005), while no apparent
differences were observed between the patients with non-active SLE
and controls (P=0.37; Fig. 1).
There were no statistically significant differences between males
and females in the patients or in the control group. In the present
study, no correlation was observed between the methylation value
and the patient’s age, although other studies (22) have reported age-related changes in
DNA methylation.
Comparison of DNA methylation following
various dosages of UVB radiation
The DNA methylation levels following irradiation
with various dosages of UVB are shown in Table I. The level of DNA methylation
decreased following UVB irradiation in the SLE patients and
controls. The reduction in DNA methylation was statistically
significant following irradiation with 50 mJ/cm2 UVB in
the patients with active SLE (P<0.005), while the decrease was
statistically significant following irradiation with 100
mJ/cm2 UVB in the non-active SLE patients (P<0.005)
and controls (P<0.005). It was observed that the patients with
active SLE had markedly lower levels of DNA methylation
(P<0.005) following irradiation with 100 mJ/cm2
UVB.
 | Table ILevel of DNA methylation following
various dosages of UVB radiation in SLE patients and controls. |
Table I
Level of DNA methylation following
various dosages of UVB radiation in SLE patients and controls.
| Group | Cases | 0
mJ/cm2 | 50
mJ/cm2 | 100
mJ/cm2 |
|---|
| Active SLE | 16 | 8.469±1.458 | 7.061±1.326a | 6.148±1.324a |
| Non-active SLE | 19 | 11.090±1.418 | 10.186±1.385 | 8.276±1.411a |
| Controls | 21 | 11.479±1.291 | 10.869±1.326 | 9.883±1.408a |
mRNA levels of DNMT1 and MBD2 in SLE
patients
Quantitative real-time PCR assays were performed to
evaluate the mRNA levels of DNMT1 and MBD2 in the T cells. β-actin
was selected as a control for normalizing the mRNA levels, as its
expression does not change in the T cells of patients with lupus
(23). Patients with SLE had
significantly lower levels of DNMT1 mRNA (0.484±0.158) compared
with the controls (0.762±0.194) and the difference was
statistically significant (P<0.005). The levels of DNMT1 mRNA
were lower in active patients (SLEDAI ≥5), but there was no
significant difference between the active and non-active patients
(SLEDAI <5; 0.449±0.160 vs. 0.514±0.154, respectively;
P=0.2352). The MBD2 mRNA levels were significantly higher in the
patients with active SLE compared with the non-active SLE patients
(1.360±0.310 vs. 1.074±0.274, respectively; P=0.0066), while there
was no significant difference between the controls (0.970±0.269)
and non-active patients (P=0.2354; Fig. 1). There was no correlation between
the levels of mRNA and the age of the patient.
Comparison of mRNA levels of DNMT1 and
MBD2 following various dosages of radiation
The mRNA levels of DNMT1 and MBD2 following various
dosages of UVB irradiation are shown in Tables II and III, respectively. The expression of DNMT1
decreased following 100 mJ/cm2 UVB irradiation in the
patients with active SLE and the difference was statistically
significant (P=0.0166). No significant differences were observed in
the expression of DNMT1 following various dosages of UVB
irradiation (50 mJ/cm2 and 100 mJ/cm2) in the
non-active SLE patients (P>0.05) and the controls (P>0.05).
There were no significant difference in the levels of MBD2 mRNA
following various dosages of UVB irradiation in the patients
(active SLE and non-active SLE) and controls (P>0.05).
| Table IILevel of DNMT1 mRNA following various
dosages of UVB radiation in SLE patients and controls. |
Table II
Level of DNMT1 mRNA following various
dosages of UVB radiation in SLE patients and controls.
| Group | Cases | 0
mJ/cm2 | 50
mJ/cm2 | 100
mJ/cm2 |
|---|
| Active SLE | 16 | 0.449±0.160 | 0.402±0.137 | 0.364±0.127a |
| Non-active SLE | 19 | 0.514±0.154 | 0.494±0.144 | 0.488±0.140 |
| Controls | 21 | 0.762±0.194 | 0.734±0.191 | 0.701±0.161 |
| Table IIILevel of MBD2 mRNA following various
dosages of UVB radiation in SLE patients and controls. |
Table III
Level of MBD2 mRNA following various
dosages of UVB radiation in SLE patients and controls.
| Group | Cases | 0
mJ/cm2 | 50
mJ/cm2 | 100
mJ/cm2 |
|---|
| Active-SLE | 16 | 1.360±0.310 | 1.334±0.303 | 1.386±0.312 |
| Non-active SLE | 19 | 1.074±0.274 | 1.104±0.280 | 1.124±0.296 |
| Controls | 21 | 0.970±0.269 | 0.966±0.283 | 0.991±0.284 |
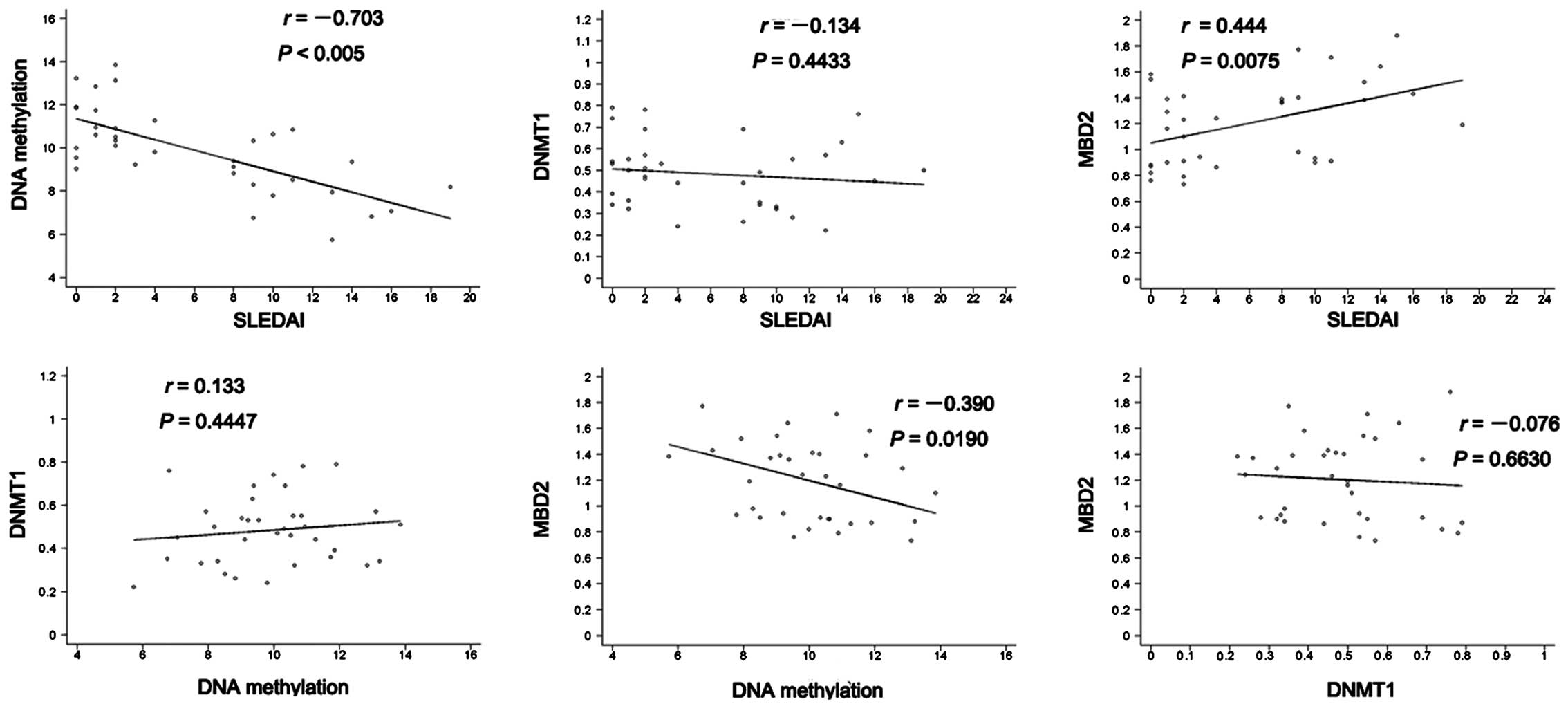
Correlations between DNA hypomethylation,
levels of DNMT1 and MBD2 mRNA and disease activity
The associations between the DNA hypomethylation and
the mRNA levels of DNMT1 and MBD2 in the SLE patients were
analyzed. DNA methylation showed an inverse correlation with SLEDAI
in the SLE patients and the correlation was statistically
significant (r=−0.703, P<0.005). However, no positive or
negative correlations with DNA methylation were observed for the
DNMT1 levels in the SLE patients. A negative correlation was
observed between MBD2 expression and DNA methylation in the
patients with SLE (r=−0.39, P=0.019). Furthermore, the level of
MBD2 expression was positively correlated with SLEDAI (r=−0.444,
P=0.0075). No correlation between the mRNA levels of DNMT1 and MBD2
was observed in the SLE patients (r=−0.076, P=0.663; Fig. 2).
In addition, whether medications (such as prednisone
and cyclophosphamide) are responsible for T-cell DNA
hypomethylation and the changes in the mRNA levels of DNMT1 and
MBD2 was investigated. No statistically significant differences
were observed between the treated and untreated groups with respect
to either DNA methylation or the mRNA levels of DNMT1 and MBD2.
Following UVB irradiation, the DNA methylation level
in the patients with active SLE (SLEDAI ≥5) was significantly lower
compared with that of the non-active SLE patients (SLEDAI <5).
The DNA methylation levels of all patients decreased following UVB
irradiation irrespective of whether the patients were treated.
Furthermore, the mRNA levels of DNMT1 decreased in the patients
with active SLE following UVB irradiation, irrespective the
treatment with medications such as prednisone and
cyclophosphamide.
Whether DNA methylation was correlated with
laboratory parameters, such as anti-dsDNA titers, and C3 and C4
concentrations, was also studied. No statistically significant
positive or negative correlations were observed between DNA
methylation and the laboratory parameters. Moreover, no differences
were observed between these groups with regard to the mRNA levels
of DNMT1 and MBD2.
Discussion
SLE is a chronic relapsing autoimmune disorder
characterized by multiple T-cell biochemical abnormalities and
autoantibody production (24).
However, the mechanisms underlying idiopathic SLE remain unclear.
Previous studies have indicated that genetic and environmental
factors contribute to the pathogenesis of SLE.
DNA methylation refers to the addition of a methyl
group to position 5 of the cytosine ring. This process, catalyzed
by a group of DNA methyltransferase, occurs in CG pairs, usually
clustered within or around promoter sequences to form CpG islands.
DNA methylation is an epigenetic process linked to the regulation
of several biological events, including embryonic development,
transcriptional regulation of gene expression, X chromosome
inactivation, genomic ‘imprinting’, chromatin modification and
silencing endogenous retroviruses (25,26).
Any changes in the pattern of DNA methylation in the promoters of
mature cells may have pathological consequences. Changes to the
methylation pattern have been reported to contribute to the
development of several diseases (10,11)
and certain hereditary immunodeficiency disorders are known to be
associated with abnormal methylation patterns (27,28).
It has been demonstrated that the adoptive transfer of T cells made
auto-reactive by treatment with DNA methylation inhibitors
(29,30) is sufficient to cause a lupus-like
disease in unirradiated syngeneic mice. A growing body of evidence
suggests a role for abnormal T-cell DNA methylation in causing
drug-induced and idiopathic lupus (31).
The underlying mechanism by which hypomethylated DNA
exists in SLE patients remains unclear. DNMT1 maintains the levels
and patterns of methylated DNA during mitosis, which has a five- to
30-fold preference for hemimethylated substrates (32), while DNA methyltransferase (DNMT)
3A and 3B (DNMT3B) are responsible for de novo DNA
methylation during development (33). Certain studies (32,34)
have observed that T cells from patients with active lupus have
diminished DNMT1 mRNA levels. The observation that inhibiting
ras-MAPK signaling decreases DNMT1 expression to the same extent as
is observed in lupus and that this decrease correlates with DNA
hypomethylation in vitro and in vivo, suggests that
the defect in DNMT1 is the primary reason for DNA hypomethylation
in SLE (35).
Several types of methyl-CpG binding proteins (MBDs)
have been identified, including MBD1, MBD2, MBD3, MBD4, MeCp2 and
Kaiso (36,37). MBD2 binds methylated cytosine and
attracts chromatin inactivation complexes containing histone
deacetylase. MBD2 is able to demethylate DNA (12,38)
and is also a transcriptional repressor of genes (39).
UV irradiation has been shown to lead to specific
demethylation events during subsequent rounds of replication
(40). As a consequence of
aberrant repair or repair methylation, UV irradiation has also been
used to activate the transcription of a quiescent metallothionine
gene, which, based on 5-azacytidine-reactivation experiments, is
considered to be under methylation control (41). In clinical practice, UV
irradiation, particularly with UVB light (290–320 nm), may induce a
flare-up of lupus disease activity (42).
To investigate the role of UVB in the pathogenesis
of SLE, the DNA methylation levels and mRNA levels of DNMT1 and
MBD2 were first analyzed in patients with SLE and in controls
without UVB irradiation. The results showed that, without UVB
irradiation, the patients with SLE had significantly lower DNA
methylation levels, lower expression levels of DNMT1 and higher
expression levels of MBD2 mRNA compared with the controls. Certain
studies (42) have observed that
the levels of DNMT1 decrease with aging and that this decrease is
correlated with the hypomethylation of sequences flanking the ITGAL
promoter, a gene which encodes CD11a, a subunit of LFA-1. This
hypomethylation may be a contributing mechanism to increased T-cell
gene expression and the development of autoimmunity with aging
(29). In the present study, no
correlation existed between DNMT1 mRNA levels and SLEDAI and no
correlation was observed between DNMT1 mRNA levels and age in the
control group and SLE patients without UVB irradiation, while the
levels of DNMT1 mRNA were similar in males and females. In the
present study, an inverse correlation was observed between
methylation and SLEDAI in the patients without UVB irradiation,
while no correlation was observed between methylation and DNMT1
mRNA levels in the group of patients without UVB irradiation.
Furthermore, the level of MBD2 expression was negatively correlated
with DNA methylation in patients with SLE without UVB irradiation
and positively correlated with SLEDAI. Due to the demethylation
effect of MBD2, the increased expression of MBD2 mRNA may be
associated with DNA hypomethylation in patients with SLE. This is
compatible with the study by Balada et al (43).
DNA methylation decreased following irradiation with
various dosages of UVB. Moreover, DNA methylation levels decreased
significantly in patients with active SLE following a low dose of
UVB (50 mJ/cm2) radiation, but decreased significantly
only following a high dose of UVB (100 mJ/cm2) radiation
in the non-active SLE patients and controls. The mRNA levels of
DNMT1 decreased significantly after the high dose of UVB (100
mJ/cm2) radiation in the patients with active SLE, while
no significant differences were observed in the non-active SLE
patients and controls following the two radiation dosages. No
significant differences were observed in the levels of MBD2 mRNA
following the different dosages of UVB irradiation in the patients
and controls. Patients with active SLE appear to have a higher
sensitivity to UVB exposure that results in significant decreases
in DNA methylation levels, suggesting that UVB exposure may have a
role in the pathogenesis of SLE by decreasing DNA methylation,
which may be caused by the decreased expression of DNMT1. This is
consistent with the clinical phenomenon whereby active patients are
more sensitive to sunlight.
In the present study, neither DNA hypomethylation
nor the mRNA levels of DNMT1 and MBD2 were accounted for by the
type of medication the patients were taking, with or without UVB
irradiation. Certain studies have reported that the drugs commonly
used to treat lupus do not inhibit DNA methylation (44,45);
changes in the methylation status of SLE patients are unlikely to
be the result of corticosteroid treatment (44,46)
and the mRNA expression levels of DNMT1, DNMT3a, DNMT3b, MBD2 and
MBD4 are not affected (43,44).
Similar results in which there were no differences between
non-treated patients and patients who had received medication (such
as prednisone and cyclophosphamide) irrespective of UVB irradiation
were observed in the present study.
It has been reported that SLE patients in remission
have normal DNA methylation levels and reduced amounts of
5-methylcytosine are observed in the T-cell DNA of active patients,
but not non-active patients (44).
A negative correlation was also observed between methylation and
SLEDAI iin the current study. No correlation were observed between
the laboratory parameters tested and DNA methylation or the mRNA
levels of DNMT1 and MBD2. In one study (46), it was noted that T cells from
patients with lupus exhibit diminished levels of DNMT activity.
Another study (47) suggested that
the decrease in DNMT enzyme activity is due to a decrease in the
level of the DNMT mRNA. It has been observed that the mRNA levels
of the MBDs (MBD2 and MeCP2) involved in the DNA methylation
process are significantly higher in lupus patients (48). We suggest that the laboratory
parameters may reflect the disease activity but do not completely
parallel the fundamental defects, such as the epigenetics of the
disease, and the global DNA methylation may depend on multiple
factors, such as the DNMT mRNA levels, DNMT activity, MBD mRNA
levels and the transcript levels of other enzymes involved on the
DNA methylation process. It is possible that a complicated
regulatory mechanism for these enzymes may be involved in human T
cells.
In conclusion, epigenetic changes appears to be
important in the regulation of SLE. Global DNA hypomethylation has
an important role in the pathogenesis of SLE. Lower expression
levels of DNMT1 mRNA and higher expression levels of MBD2 mRNA may
be involved in the pathogenesis of SLE, but are not the only
regulation factors for global DNA methylation in SLE. DNA
methylation and DNMT1 mRNA expression levels decrease in patients
with active SLE following UVB exposure, indicating that UVB may
have a role in the pathogenesis of SLE. The mechanism of the effect
of UVB on DNA methylation in SLE patients is complicated and should
be studied further. This may provide important evidence for
assisting in the development of new treatments for SLE through sun
protection, as well as providing a deeper understanding of the
etiology of this disease.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (30972656) and
National Natural Science Foundation of Shanghai (12ZR1404500). The
authors would like to thank Dr Ke-fei Kang for his critical review
of the manuscript.
References
|
1
|
Sestak AL, Nath SK, Sawalha AH and Harley
JB: Current status of lupus genetics. Arthritis Res Ther.
9:2102007. View
Article : Google Scholar
|
|
2
|
Jönsen A, Bengtsson AA, Nived O, Truedsson
L and Sturfelt G: Gene-environment interactions in the aetiology of
systemic lupus erythematosus. Autoimmunity. 40:613–617.
2007.PubMed/NCBI
|
|
3
|
Attwood JT, Yung RL and Richardson BC: DNA
methylation and the regulation of gene transcription. Cell Mol Life
Sci. 59:241–257. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Richardson BC: Role of DNA methylation in
the regulation of cell function: autoimmunity, aging and cancer. J
Nutr. 132(8 Suppl): 2401S–2405S. 2002.PubMed/NCBI
|
|
5
|
Richardson BC, Strahler JR, Pivirotto TS,
et al: Phenotypic and functional similarities between
5-azacytidine-treated T cells and a T cell subset in patients with
active systemic lupus erythematosus. Arthritis Rheum. 35:647–662.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yung R, Powers D, Johnson K, et al:
Mechanisms of drug-induced lupus. II. T cells overexpressing
lymphocyte function-associated antigen 1 become autoreactive and
cause a lupuslike disease in syngeneic mice. J Clin Invest.
97:2866–2871. 1996. View Article : Google Scholar
|
|
7
|
Quddus J, Johnson KJ, Gavalchin J, et al:
Treating activated CD4+ T cells with either of two
distinct DNA methyltransferase inhibitors, 5-azacytidine or
procainamide, is sufficient to cause a lupus-like disease in
syngeneic mice. J Clin Invest. 92:38–53. 1993.
|
|
8
|
Kaplan MJ, Lu Q, Wu A, Attwood J and
Richardson B: Demethylation of promoter regulatory elements
contributes to perforin overexpression in CD4+ lupus T
cells. J Immunol. 172:3652–3661. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu Q, Ray D, Gutsch D and Richardson B:
Effect of DNA methylation and chromatin structure on ITGAL
expression. Blood. 99:4503–4508. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakao M: Epigenetics: interaction of DNA
methylation and chromatin. Gene. 278:25–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Richardson B and Yung R: Role of DNA
methylation in the regulation of cell function. J Lab Clin Med.
134:333–340. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Detich N, Theberge J and Szyf M:
Promoter-specific activation and demethylation by MBD2/demethylase.
J Biol Chem. 277:35791–35794. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tan EM, Cohen AS, Fries JF, et al: The
1982 revised criteria for the classification of systemic lupus
erythematosus. Arthritis Rheum. 25:1271–1277. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wysenbeek AJ, Block DA and Fries JF:
Prevalence and expression of photosensitivity in systemic lupus
erythematosus. Ann Rheum Dis. 48:461–463. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lehmann P, Hölzle E, Kind P, Goerz G and
Plewig G: Experimental reproduction of skin lesions in lupus
erythematosus by UVA and UVB radiation. J Am Acad Dermatol.
22:181–187. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bombardier C, Gladman DD, Urowitz MB,
Caron D and Chang CH: Derivation of the SLEDAI. A disease activity
index for lupus patients The Committee on Prognosis Studies in SLE.
Arthritis Rheum. 35:630–640. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
WHO: Global Solar UV Index: A Practical
Guide. World Health Organization; Geneva, Switzerland: 2002
|
|
18
|
Gies P: Australia has more than enough
solar UV radiation. Clin Exp Optom. 86:71–73. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Habib M, Fares F, Bourgeois CA, et al: DNA
global hypomethylation in EBV-transformed interphase nuclei. Exp
Cell Res. 249:46–53. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Milutinovic S, Zhuang Q, Niveleau A and
Szyf M: Epigenomic stress response. Knockdown of DNA
methyltransferase 1 triggers an intra-S-phase arrest of DNA
replication and induction of stress response genes. J Biol Chem.
278:14985–14995. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X and Seed B: A PCR primer bank for
quantitative gene expression analysis. Nucleic Acids Res.
31:e1542003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fuke C, Shimabukuro M, Petronis A, et al:
Age related changes in 5-methylcytosine content in human peripheral
leukocytes and placentas: an HPLC-based study. Ann Hum Genet.
68:196–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Klinman DM, Mushinski JF, Honda M, et al:
Oncogene expression in autoimmune and normal peripheral blood
mononuclear cells. J Exp Med. 163:1292–1307. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tsokos GC and Kammer GM: Molecular
aberrations in human systemic lupus erythematosus. Mol Med Today.
6:418–424. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Surani MA: Imprinting and the initiation
of gene silencing in the germ line. Cell. 93:309–312. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ng HH and Bird A: DNA methylation and
chromatin modification. Curr Opin Genet Dev. 9:158–163. 1999.
View Article : Google Scholar
|
|
27
|
Knight SJ, Flannery AV, Hirst MC, et al:
Trinucleotide repeat amplification and hypermethylation of a CpG
island in FRAXE mental retardation. Cell. 74:127–134. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu GL, Bestor TH, Bourc’his D, et al:
Chromosome instability and immunodeficiency syndrome caused by
mutations in a DNA methyltransferase gene. Nature. 402:187–191.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yung R, Chang S, Hemati N, Johnson K and
Richardson B: Mechanisms of drug-induced lupus. IV. Comparison of
procainamide and hydralazine with analogs in vitro and in
vivo. Arthritis Rheum. 40:1436–1443. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yung RL, Quddus J, Chrisp CE, Johnson KJ
and Richardson BC: Mechanism of drug-induced lupus. I. Cloned Th2
cells modified with DNA methylation inhibitors in vitro
cause autoimmunity in vivo. J Immunol. 154:3025–3035.
1995.PubMed/NCBI
|
|
31
|
Sawalha AH and Richardson B: DNA
methylation in the pathogenesis of systemic lupus erythematosus.
Curr Pharmacogenomics. 3:73–78. 2005. View Article : Google Scholar
|
|
32
|
Deng C, Yang J, Scott J, Hanash S and
Richardson BC: Role of the ras-MAPK signaling pathway in the DNA
methyltransferase response to DNA hypomethylation. Biol Chem.
379:1113–1120. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Okano M, Xie S and Li E: Cloning and
characterization of a family of novel mammalian DNA (cytosine-5)
methyltransferases. Nat Genet. 19:219–220. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ogasawara H, Okada M, Kaneko H, Hishikawa
T, Sekigawa I and Hashimoto H: Possible role of DNA hypomethylation
in the induction of SLE: relationship to the transcription of human
endogenous retroviruses. Clin Exp Rheumatol. 21:733–738.
2003.PubMed/NCBI
|
|
35
|
Vertino PM, Yen RW, Gao J and Baylin SB:
De novo methylation of CpG island sequences in human
fibroblasts overexpressing DNA (cytosine-5-)-methyltransferase. Mol
Cell Biol. 16:4555–4565. 1996.PubMed/NCBI
|
|
36
|
Fan G and Hutnick L: Methyl-CpG binding
proteins in the nervous system. Cell Res. 15:255–261. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Klose RJ and Bird AP: Genomic DNA
methylation: the mark and its mediators. Trends Biochem Sci.
31:89–97. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bhattacharya SK, Ramchandani S, Cervoni N
and Szyf M: A mammalian protein with specific demethylase activity
for mCpG DNA. Nature. 397:579–583. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ng HH, Zhang Y, Hendrich B, et al: MBD2 is
a transcriptional repressor belonging to the MeCP1 histone
deacetylase complex. Nat Genet. 23:58–61. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kastan MB, Gowans BJ and Lieberman MW:
Methylation of deoxycytidine incorporated by excision-repair
synthesis of DNA. Cell. 30:509–516. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lieberman MW, Beach LR and Palmiter RD:
Ultraviolet radiation-induced metallothionein-I gene activation is
associated with extensive DNA demethylation. Cell. 35:207–214.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Z, Deng C, Lu Q and Richardson B:
Age-dependent DNA methylation changes in the ITGAL (CD11a)
promoter. Mech Ageing Dev. 123:1257–1268. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Balada E, Ordi-Ros J and Vilardell-Tarrés
M: DNA methylation and systemic lupus erythematosus. Ann NY Acad
Sci. 1108:127–136. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Richardson B, Scheinbart L, Strahler J,
Gross L, Hanash S and Johnson M: Evidence for impaired T cell DNA
methylation in systemic lupus erythematosus and rheumatoid
arthritis. Arthritis Rheum. 33:1665–1673. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nyce J, Liu L and Jones PA: Variable
effects of DNA-synthesis inhibitors upon DNA methylation in
mammalian cells. Nucleic Acids Res. 14:4353–4367. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Deng C, Kaplan MJ, Yang J, et al:
Decreased Ras-mitogen-activated protein kinase signaling may cause
DNA hypomethylation in T lymphocytes from lupus patients. Arthritis
Rheum. 44:397–407. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yoder JA, Soman NS, Verdine GL and Bestor
TH: DNA (cytosine-5)-methyltransferases in mouse cells and tissues.
Studies with a mechanism-based probe. J Mol Biol. 270:385–395.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lei W, Luo Y, Lei W, et al: Abnormal DNA
methylation in CD4+ T cells from patients with systemic
lupus erythematosus, systemic sclerosis, and dermatomyositis. Scand
J Rheumatol. 38:369–374. 2009.PubMed/NCBI
|