Introduction
It is well known that heme oxygenase (HO) functions
as the rate-limiting enzyme in heme degradation, which takes place
in the endoplasmic reticulum. There are three isoforms of HO in the
body, namely HO-1, 2 and 3 (1). HO
has been reported to be present in all tissues and is located in
microsomes (2). Recently HO-1 and
2 have been shown to be present in mitochondria (3,4).
HO-1 is inducible by inflammatory cytokines and oxidants, including
nitric oxide (NO), whereas HO-2 and 3 are expressed constitutively
(1,5). It was reported that HO-1 mRNA was
present in different regions of the brain, especially the
hippocampus and cerebellum (6).
Certain studies have reported impaired spatial navigation learning
ability in transgenic mice overexpressing HO-1 (7). Other studies have shown an
age-related decrease in HO-1 expression to be present in specific
brain regions, including the hippocampus. Further, neotrofin (AIT),
a cognitive-enhancing and neuroprotective drug, was found to cause
a robust increase in HO-1 immunoreactive protein in the same
regions (8). Alzheimer’s disease
(AD) is a common age-associated dementia featuring progressive loss
of neurons and synapses, gliosis and the accumulation of intra- and
extracellular protein deposits. In individuals with AD, the
increasing impairment of learning and memory eventually leads to a
definitive diagnosis. It appears that oxidative injury is central
in the pathogenesis, even prior to the appearance of amyloid
deposits (9,10). Intrahippocampal injection of a
lentiviral vector expressing nuclear factor (erythroid-derived
2)-like 2 (Nrf2) was found to improve spatial learning in a mouse
model of Alzheimer’s disease and Nrf2 gene transfer was associated
with a robust reduction in astrocytic but not microglial activation
as well as the induction of the Nrf2 target gene HO-1 (11). It appears that the role of HO-1 in
the modulation of learning ability is complex. The aim of the
present study was to elucidate the correlation between HO-1 and
learning.
Materials and methods
Subjects
Thirty-five nine-day-old male NIH mice were housed
in polyethylene cages (five mice per cage) and fed with standard
chow pellets and drinking water until they reached 55 days old.
This study was carried out in strict accordance with the
recommendations in the Guide for the Care and Use of Laboratory
Animals of the National Institutes of Health. The animal use
protocol was reviewed and approved by the Institutional Animal Care
and Use Committee (IACUC) of the Second Affiliated Hospital of
Chongqing Medical University, China. All experiments were conducted
in accordance with the guidelines of Chongqing Medical University
and the Animal Care Committee.
Surgery
Following anesthesia with 4% chloral hydrate, a
stainless steel cylindrical cannula (outer diameter, 0.6 mm; inner
diameter, 0.4 mm) with a stopper was implanted so that the tip of
the cannula was in the left lateral ventricle (1.3 mm lateral to
the midline, 0.3 mm posterior to the bregma, 2.0 mm ventral to the
dura). The cylindrical cannula was fixed with dental cement mixed
with fast condensation glue. During the surgery, body temperature
was monitored and maintained at 37±0.5°C.
Behavioral test/training
In the step-down test, mice were placed on the
platform. If the mice stepped down onto the floor they received a
36 V AC foot shock. Mice typically jumped quickly onto the platform
to avoid the electric stimulation. The error number (more than two
extremities touching the grid) and the electric shock time were
recorded for 10 min. A day later, the mice were placed on the
platform again but without electrifying the grid. The step-down
latency and the time remaining on the platform were recorded over a
5 min period.
In the step-through test, mice were first placed in
the illuminated compartment. Mice would typically enter the dark
compartment and encounter a 42 V AC foot shock. The time taken to
enter the dark compartment from the illuminated one was recorded.
Memory retention trials were performed 24 h later by placing the
mice into the illuminated compartment and measuring the dark
component entry latency.
In the Morris water maze test, the mice were placed
in the maze for 60 sec without a platform. A hidden platform was
then placed in the center of one quadrant. The mice were given four
trials per day for four consecutive days. In each trial, the mice
were placed in the water, facing the edge of the pool in one of the
quadrants. The start position varied in a quasi-random fashion, so
that the mice never started at the same place in two consecutive
trials. The mice were then allowed to seek the hidden platform. If
a mouse failed to find the platform in 60 sec, it was placed on the
platform for another 15 sec. There was an inter-trial interval of
30 min. On the fifth day, the mice were placed in a randomly
selected quadrant and the time to reach the hidden platform and
swim speed were recorded.
Measurement of heme oxygenase
activity
Hippocampi were dissected on ice and samples were
stored in liquid nitrogen until analysis. The samples were
homogenized for 30 sec in three volumes of 0.01 mol/l
trisaminomethane hydrochloride (Tris-HCl) containing 0.01 mol/l
sucrose and 0.0001 mol/l ethylenediaminetetra-acetic acid disodium
salt (EDTANa2; 4°C, pH 7.4) using a UP-50H ultra-sonic
homogenizer and centrifuged at 4°C at 18,800 ± g for 15 min.
Supernatant (0.2 ml) was added to the reaction mixture [2 mmol/l
glucose-6-phosphate (Sigma, St. Louis, MO, USA), 0.2 units
glucose-6-phosphate dehydrogenase (Sigma), 20 μmol/l hemin,
2 mg rat liver cytoplasm and 0.8 mmol/l nicotinamide adenine
dinucleotide phosphate-oxidase (NADPH; Sigma)]. The total volume
was 1.0 ml. The mixture was aerobically incubated for 30 min at
37°C in dark conditions and stopped with the addition of 1.0 ml
ice-cold chloroform. The production of bilirubin was measured with
a double-beam spectrophotometer at ΔOD at 530 nm (extinction
coefficient, 40 mM cm−1 for bilirubin). Protein
concentration was determined using the Coomassie blue method.
Western blot analysis
Hippocampal tissue was lysed in a solubilization
buffer containing 10 mM Tris-HCl (pH 7.4), 0.15 mM NaCl, 1% Nonidet
P-40 (Sigma), 0.1% sodium dodecyl sulfate (SDS), 0.001 mg/ml
leupeptin (Sigma), 0.001 mg/ml pepstatin (Sigma), 0.001 mg/ml
aprotonin (Sigma) and 10% phenylmethylsulfonyl fluoride (PMSF;
Sigma) and centrifuged at 4°C at 20,000 × g for 15 min. Protein
concentration was estimated using the Bradford method. Protein (40
μg) was subjected to 12% SDS polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred to a nitrocellulose
membrane. The membrane was blocked for 1 h in phosphate-buffered
saline (PBS) containing 5% fat-free milk and 0.2% Tween-20. The
blot was incubated for 2 h at 37°C separately with the primary
antibody for HO-1 (Sigma), HO -2 (Sigma) and β-actin (Santa Cruz
Biotechnology Inc., Santa Cruz, CA, USA), at a 1:400 concentration,
followed by incubation for 1 h at 37°C with the secondary antibody
(peroxidase-conjugated goat anti-rabbit IgG (Santa Cruz).
Immunoreactive bands of HO-1 and 2 were visualized with
chemiluminescence reagents. The chemiluminescent signal of the band
was detected using a lumino image analyzer (Bio-Rad, Hercules, CA,
USA).
Real-time PCR
Total RNA of hippocampus was isolated according to
the RNA isolation kit (Takara Bio, Inc., Shiga, Japan)
instructions. Complementary DNA (cDNA) was synthesized according to
the instructions of the reverse transcription kit (Toyobo, Osaka,
Japan). Amplified DNA was visualized using an ethidium bromide
stain with a 2% agarose gel. Results were quantified using the
Quantity One analysis software (Bio-Rad). Since
glyceraldehyde-3-phosphate dehydrogenase (G3PDH) was the
housekeeping gene, amplification signals for HO-1 and 2 mRNA were
normalized with the amplification signals of G3PDH mRNA. The HO-1
5′ primer was 5′-AGCACTATGTAAAGCGTCTC-3′ and the 3′ primer was
5′-CGGTCTTAGCCTCTTCTGT-3′ (282 bp). The HO-2 5′ primer was
5′-GACCCAATTCTACCTGTTT-3′ and the 3′ primer was
5′-CCATCCTCCAGGGTTTCT-3′ (207 bp). The G3PDH 5′ primer was
5′-ACCACAGTCCATGCCATCAC-3′ and the 3′ primer was
5′-TCCACCACCCTGTTGCTGTA-3′ (450 bp).
Study design
All experiments were performed when the mice were
fifty days old. Mice were divided into six groups [Hemin, ZnPPIX,
artificial cerebrospinal fluid (ACSF), behavior training,
dark-reared and control groups]. Each group had five mice, apart
from the control group which had ten. All groups were kept in a
long daylight cycle condition (16 h light/8 h dark), apart from the
dark-reared group which were kept in dark conditions. The hemin,
ZnPPIX, ACSF, behavior training and control group (only five mice)
underwent the behavior test/training once the
intracerebroventricular (icv) injection was completed. Biochemical
experiments were conducted after the behavior test.
Statistical analysis
Data were expressed as means ± standard deviation
and were assessed using ANOVA and t-test analyses. P<0.05 was
considered to indicate a statistically significant result.
Results
Effect of icv injection with various
drugs on behavioral test performance
As shown in Table
I, there was no significant difference among the various groups
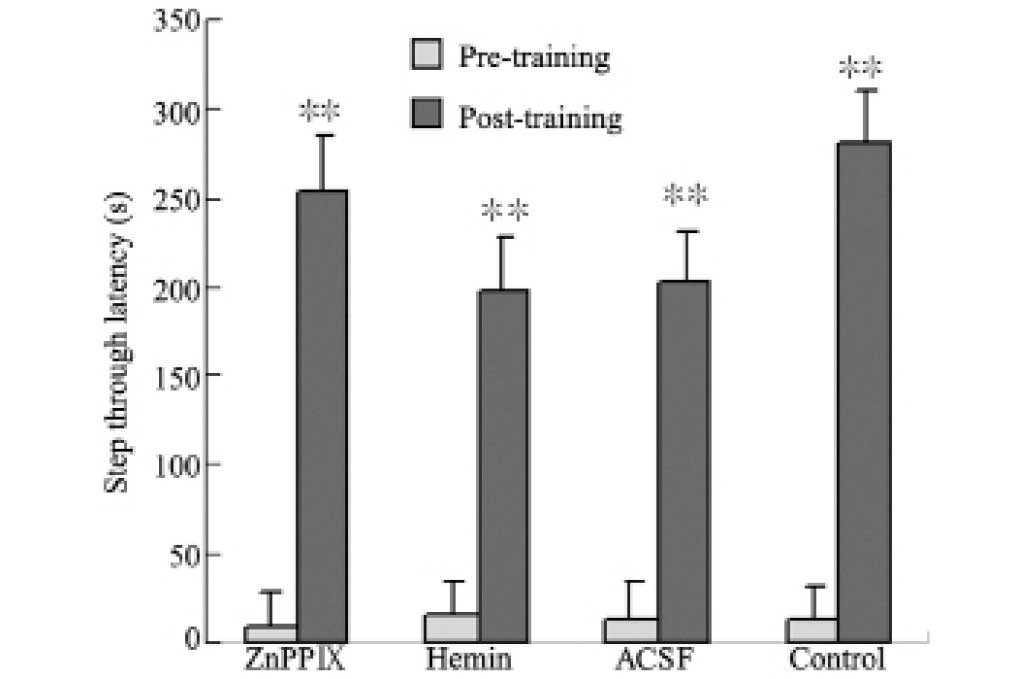
in the step-down test. In the step-through test, there was a
significant difference in the step-through latency between pre-and
post-training in each group. No significant difference was observed
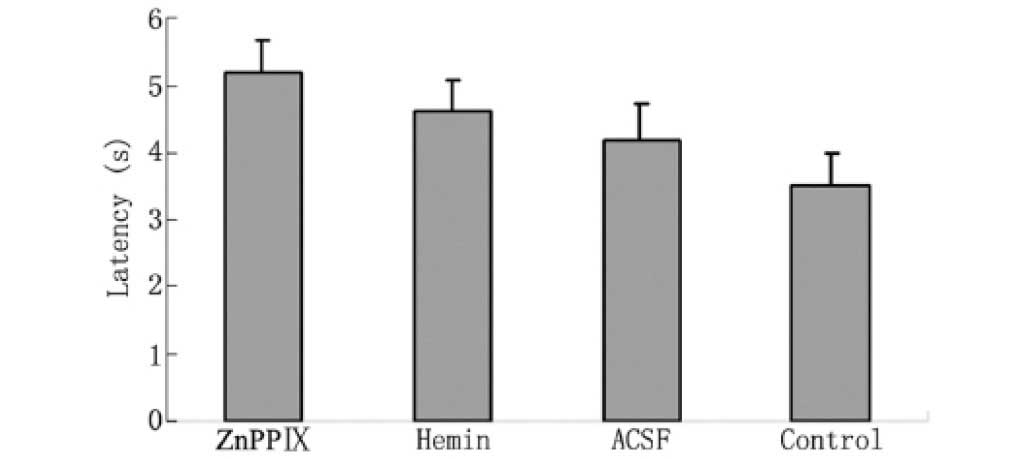
between the hemin, ACSF, ZnPPIX and control groups (Fig. 1). The data from the Morris water
maze test did not indicate any significant differences between the
pretreatment groups in swim speed or in the mean latency to reach
the platform (Fig. 2, Table II).
 | Table IEffect of intracerebroventricular
(icv) injection with various treatments on the step-down test
(n=10, means ± SEM); icv injection with various drugs had no effect
on the results of behavior tests. |
Table I
Effect of intracerebroventricular
(icv) injection with various treatments on the step-down test
(n=10, means ± SEM); icv injection with various drugs had no effect
on the results of behavior tests.
| Group | Error number | Electric shock time
(sec) | Step-down latency
(sec) | Time remaining on
platform (sec) |
|---|
| ZnPPIX | 1.8±0.41 | 30.9±15.50 | 71.9±30.17 | 158.2±30.10 |
| Hemin | 2.5±0.38 | 22.4±5.31 | 72.3±30.26 | 148.1±29.82 |
| ACSF | 1.9±0.28 | 44.0±22.42 | 83.6±31.05 | 120.5±34.12 |
| Control | 2.4±0.51 | 15.6±5.09 | 86.7±28.84 | 168.7±32.86 |
| Table IIEffect of intracerebroventricular
(icv) injection with various treatments on swim speed (n=10, means
± SEM). |
Table II
Effect of intracerebroventricular
(icv) injection with various treatments on swim speed (n=10, means
± SEM).
| Group | Swim speed
(cm/sec) |
|---|
| ZnPPIX | 22.51±4.08 |
| Hemin | 26.34±3.37 |
| ACSF | 23.78±3.56 |
| Control | 24.04±5.36 |
Effect of icv injection with various
drugs on HO protein expression
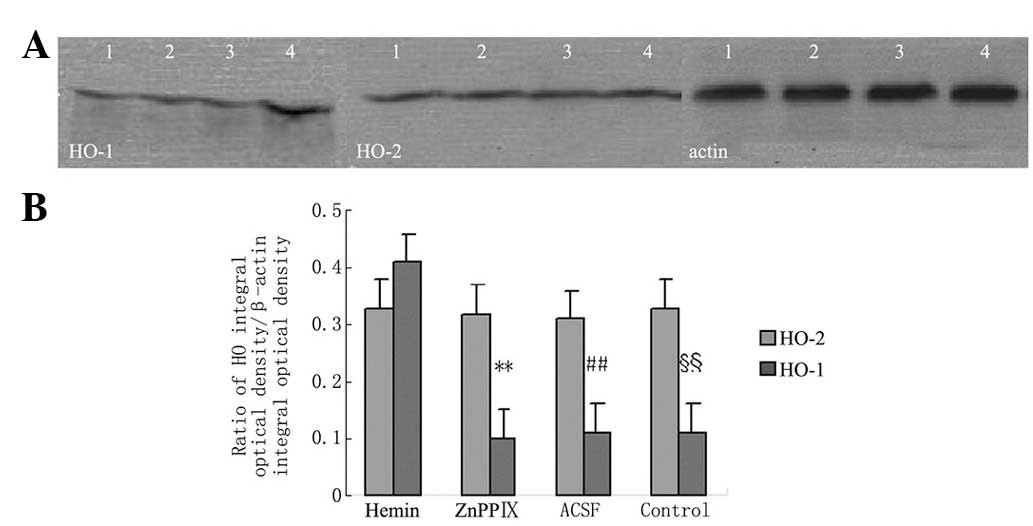
Fig. 3 shows that
hemin induced the expression of HO-1 protein. Hemin and ZnPPIX had
no effect on the expression of HO-1. None of the pretreatments
affected the expression of HO-2.
Effect of icv injection with various
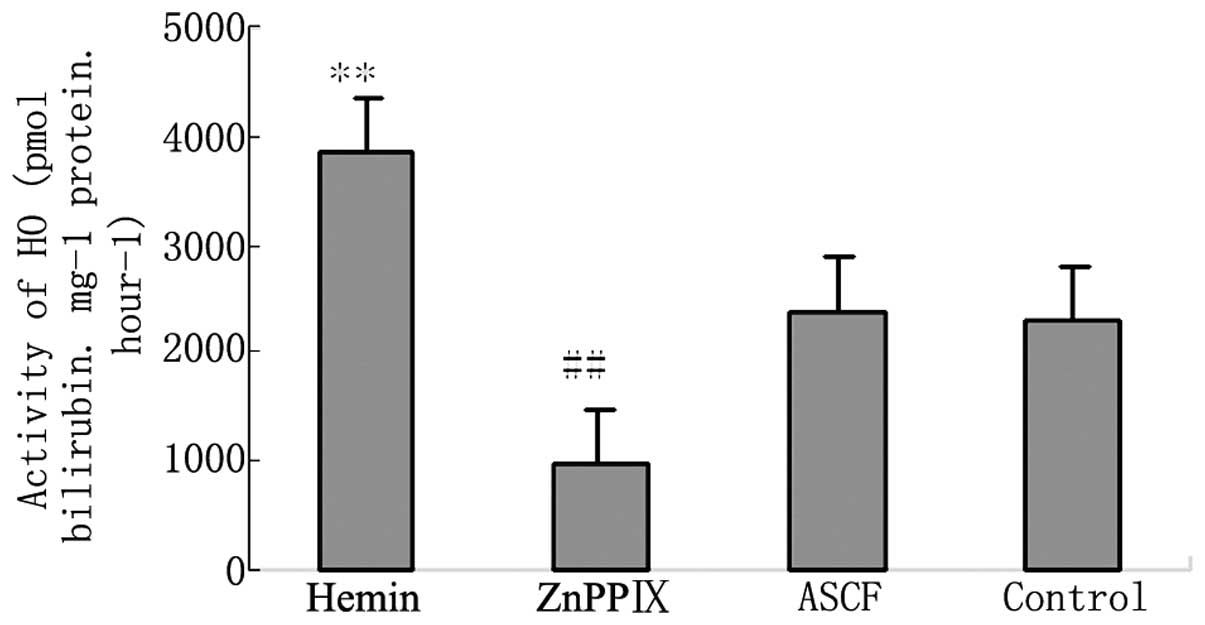
drugs on HO activity
An icv hemin injection caused the HO activity to
increase significantly. Injection of ZnPPIX icv significantly
suppressed the activity of HO. Injection of ACSF icv had no effect
on the activity of HO (Fig.
4).
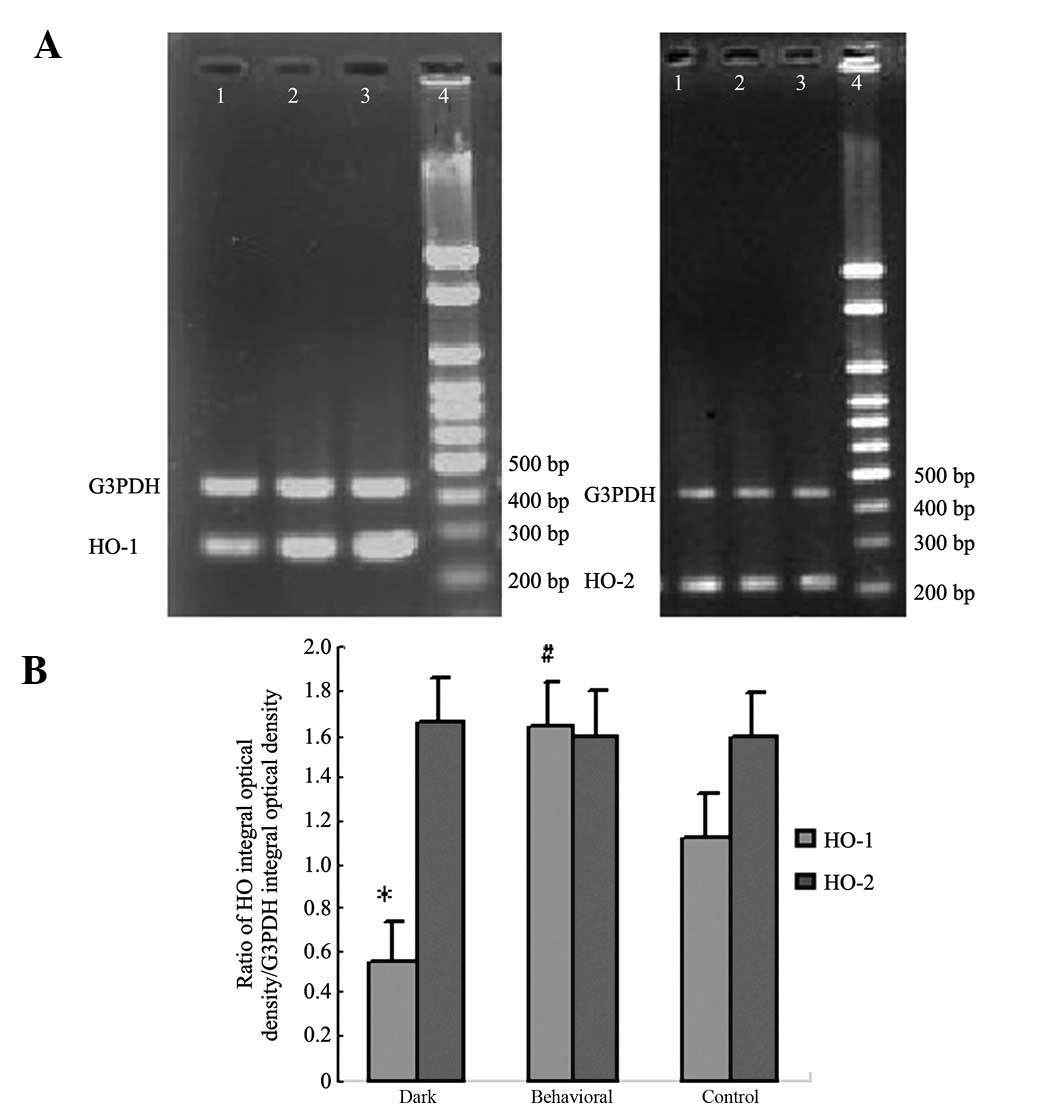
Effect of behavioral training on HO mRNA
expression
Mice reared in dark conditions showed a low
expression of HO-1 mRNA in hippocampal tissue. Behavioral training
was found to significantly upregulate the expression of HO-1
compared to the control condition. There was no significant
difference in HO-2 mRNA expression between the three groups
(Fig. 5).
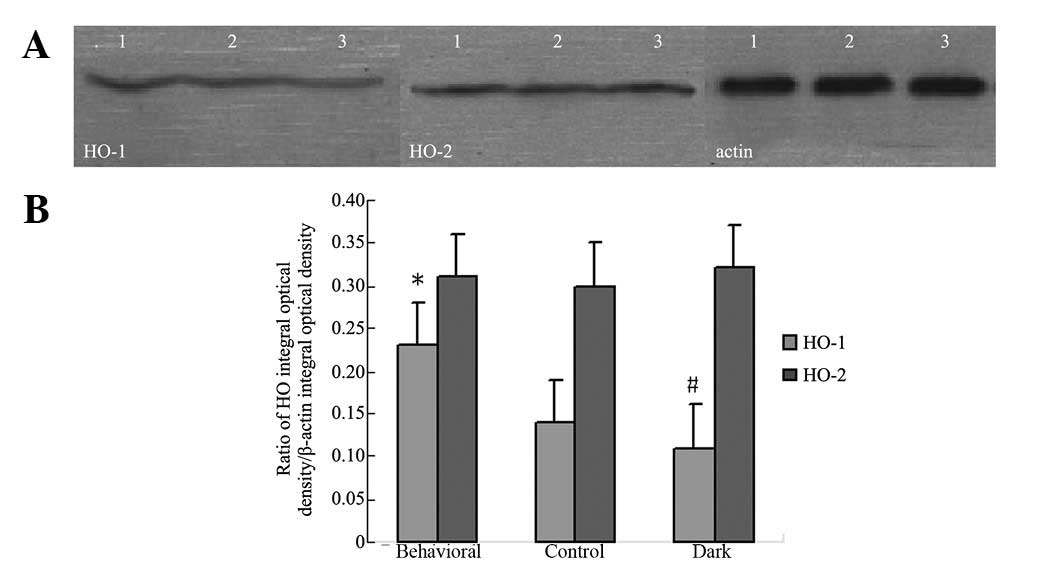
Effect of behavioral training on HO
protein expression
The HO-1 protein expression of mice reared in dark
conditions was lower than that of the control group. Behavioral
training was found to significantly upregulate HO-1 protein
expression. There was no difference in HO-2 protein expression
between the groups (Fig. 6).
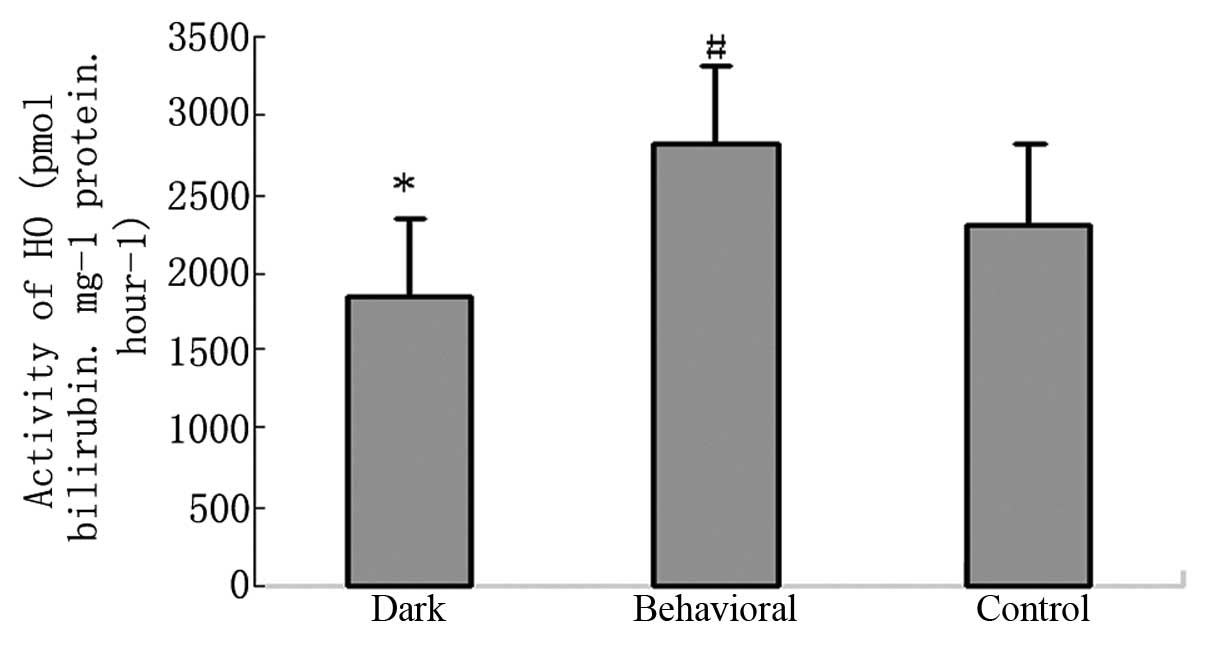
Effect of behavioral training on activity
of HO
Compared with the control group, the HO activity of
the dark-reared group was decreased. Behavioral training
significantly increased its activity (Fig. 7).
Discussion
HO-1 and 2 degrade heme to carbon monoxide (CO),
biliverdin and ferrous iron. Endogenous CO is involved in long-term
potentiation (LTP) and avoidance learning (12). A long-term activity-dependent
presynaptic enhancement during LTP has also been found to be
induced by CO (13). Biliverdin is
believed to be the most powerful endogenous antioxidant, which
efficiently scavenges peroxy radicals and protects cells from a
10,000-fold excess of hydrogen peroxide (14).
HO-2 is constitutively expressed in cells and
previous research has shown that HO-2 has no effect on memory and
learning ability (15). HO-1 is an
inducible form of HO. It may be induced by many factors, including
heme, heat shock, intensive light and ultraviolet exposure
(7). The activity of HO may be
blocked by metalloporphyrins, including chromium mesoporphyrin
(CrMP), manganese protoporphyrin, manganese mesoporphyrin (MnPP),
zinc protoporphyrin (ZnPP), tin mesoporphyrin and tin
protoporphyrin (SnPP) (16). The
expression of HO-1 protein and HO activity in the hemin icv
injection group was significantly increased when compared with the
control group. In the ZnPPIX injection group, the activity of HO
decreased without any change in HO-1 protein expression. ACSF
injected icv did not change the level of HO-1 protein or the
activity of HO. This indicates that the cannula implanting
operation itself had no effect on the expression of HO or its
activity. Consistent with previous studies, none of the
pretreatments affected the expression of HO-2 protein.
Behavioral tests were used to evaluate the effects
on learning ability of altering HO-1 in mature mice. The data show
that neither avoidance learning nor spatial navigation learning
changed with changes in HO-1. Overall, in the present study, there
was no correlation between learning ability and HO-1 in adult
mice.
In order to determine the function of HO-1 in the
immature mice, a dark-reared model was constructed. In this model,
the mice were deprived of the stress of sound and light since they
were young. It has been reported that stress affects multiple
memory systems (17). There is
some evidence to suggest that the intelligence quotient of children
with impaired vision or hearing is significantly lower than that of
children with normal vision (18,19).
According to this theory, it was possible that the intelligence
quotient of dark-reared mice was lower than that of control group,
which were raised in normal conditions and the intelligence
quotient of behavioral training mice was higher than that of
control mice. Based on these models, we detected the expression of
HO-1 and 2 by RT-PCR and western blot. Results showed that unlike
HO-2, the expression of HO-1 in the dark-reared mice was lower than
that of the control group. Moreover, the behavioral training group
was found to significantly upregulate HO-1 expression, when
compared with the control group. A similar change in HO activity
was also observed.
The results of the present study suggest that the
modification of HO-1 has no effect on established memory and
learning abilities in adult mice whose brains have developed
completely. On the other hand, the brain of a neonatal mouse is
immature and its learning ability and memory are poor. It has been
demonstrated that stimuli such as light and sound could promote the
development of memory systems, which stimulates the expression of
HO-1. HO-1 catalyzes heme to produce CO and biliverdin, which is
involved in LTP and in protecting neurons from oxidative stress. In
conclusion, the present findings suggest that the upregulation of
HO-1 expression may facilitate the memory and learning ability of
mice during development.
Acknowledgements
This study was supported by a grant
from Chongqing Medical University.
References
|
1.
|
McCoubrey WK Jr, Huang TJ and Maines MD:
Isolation and characterization of a cDNA from the rat brain that
encodes hemoprotein heme oxygenase-3. Eur J Biochem. 247:725–732.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Abraham NG and Kappas A: Pharmacological
and clinical aspects of heme oxygenase. Pharmacol Rev. 60:79–127.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Di Noia MA, Van Driesche S, Palmieri F, et
al: Heme oxygenase-1 enhances renal mitochondrial transport
carriers and cytochrome C oxidase activity in experimental
diabetes. J Biol Chem. 281:15687–15693. 2006.
|
|
4.
|
Turkseven S, Drummond G, Rezzani R, et al:
Impact of silencing HO-2 on EC-SOD and the mitochondrial signaling
pathway. J Cell Biochem. 100:815–823. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Donnelly LE and Barnes PJ: Expression of
heme oxygenase in human airway epithelial cells. Am J Respir Cell
Mol Biol. 24:295–303. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Scapagnini G, D’Agata V, Calabrese V, et
al: Gene expression profiles of heme oxygenase isoforms in the rat
brain. Brain Res. 954:51–59. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Huang Y, Wu L, Xu C, Yang B and Wang R:
Increased HO-1 expression and decreased iNOS expression in the
hippocampus from adult spontaneously hypertensive rats. Cell
Biochem Biophys. 46:35–42. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Ewing JF and Maines MD: Regulation and
expression of heme oxygenase enzymes in aged-rat brain: age related
depression in HO-1 and HO-2 expression and altered stress-response.
J Neural Transm. 113:439–454. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Resende R, Moreira PI, Proença T, et al:
Brain oxidative stress in a triple-transgenic mouse model of
Alzheimer disease. Free Radic Biol Med. 44:2051–2057. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Selkoe DJ: Alzheimer’s disease: genes,
proteins, and therapy. Physiol Rev. 81:741–766. 2001.
|
|
11.
|
Kanninen K, Heikkinen R, Malm T, et al:
Intrahippocampal injection of a lentiviral vector expressing Nrf2
improves spatial learning in a mouse model of Alzheimer’s disease.
Proc Natl Acad Sci USA. 106:16505–16510. 2009.PubMed/NCBI
|
|
12.
|
Cutajar MC and Edwards TM: Evidence for
the role of endogenous carbon monoxide in memory processing. J Cogn
Neurosci. 19:557–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Baranano DE, Rao M, Ferris CD and Snyder
SH: Biliverdin reductase: a major physiologic cytoprotectant. Proc
Natl Acad Sci USA. 99:16093–16098. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Burnett AL, Johns DG, Kriegsfeld LJ, et
al: Ejaculatory abnormalities in mice with targeted disruption of
the gene for heme oxygenase-2. Nat Med. 4:84–87. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Wu L: The pro-oxidant role of
methylglyoxal in mesenteric artery smooth muscle cells. Can J
Physiol Pharmacol. 83:63–68. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Vreman HJ, Ekstrand BC and Stevenson DK:
Selection of metalloporphyrin heme oxygenase inhibitors based on
potency and photoreactivity. Pediatr Res. 33:195–200. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Schwabe L, Oitzl MS, Philippsen C, et al:
Stress modulates the use of spatial versus stimulus-response
learning strategies in humans. Learn Mem. 14:109–116. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Niedzielski A, Humeniuk E, Błaziak P and
Gwizda G: Intellectual efficiency of children with unilateral
hearing loss. Int J Pediatr Otorhinolaryngol. 70:1529–1532. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Roizen N, Kasza K, Karrison T, et al:
Impact of visual impairment on measures of cognitive function for
children with congenital toxoplasmosis: implications for
compensatory intervention strategies. Pediatrics. 118:e379–e390.
2006. View Article : Google Scholar
|