Introduction
Insulin resistance (IR) refers to the reduced
biological efficacy of insulin on insulin effector organs and the
consequent decrease in glucose uptake and elimination in
surrounding tissues, including the liver, skeletal muscle and
adipose tissues. IR is a physiological and pathological state
wherein normal or above-normal concentrations of insulin are only
capable of exerting biological effects that are reduced compared
with those expected in the normal range. Under IR, higher levels of
insulin are required to induce a reaction comparable to that
induced by normal levels of insulin, and hyperinsulinaemia or
hyperproinsulinaemia, in which exogenous IR occurs, often accompany
the condition. Hyperinsulinaemia is one of the predominant
indicators of IR (1). Numerous
pathophysiological changes occur during IR, which increase risk
factors for various conditions, including diabetes, hypertension,
atherosclerosis, dyslipidaemia and central obesity. Considering the
high rates of morbidity and mortality associated with
cardiovascular diseases, the risks associated with IR have become a
widespread concern and the focus of medical research (2).
Obesity is a condition often accompanied by a
chronic and subclinical inflammation that is associated with IR,
insulin secretion and the development of atherosclerosis through
the secretion of inflammatory cytokines. Obesity increases the
incidence of type 2 diabetes mellitus (T2DM) and cardiovascular
events. Inhibition of the inflammatory process associated with
obesity may represent a potential pharmaceutical target for the
prevention and treatment of T2DM and coronary heart disease.
Numerous studies on fat cytokines initially identified obesity as a
low-grade inflammatory disease. Low-grade inflammation is a key
determining factor for IR, and low-grade inflammation molecules
have been associated with obesity and IR (3).
Adipose tissues are important sources of
inflammatory mediators (4), and
adiponectin is a fat cytokine closely associated with insulin
sensitivity. Numerous studies have demonstrated that, plasma
adiponectin concentrations of animals and humans are negatively
correlated with fasting blood glucose concentrations and insulin
concentrations, that are positively correlated with insulin
sensitivity. Furthermore, it has been shown that secretion of
plasma adiponectin is affected by peroxisome proliferator-activated
receptor γ (PPAR-γ) and insulin receptor substrate 1 (IRS-1), and
that adiponectin expression is negatively correlated with
interleukin (IL)-6 (5).
Astragalus polysaccharides (APS), which are capable
of improving IR and reducing blood glucose levels, are key
components of Astragalus mongholicus, which is widely used
in Traditional Chinese Medicine. Studies have demonstrated that APS
exert their functions through PPAR-γ and IRS-1 (6,7). The
aim of this study was to investigate the effect of APS on
adiponectin and IL-6 secretion by 3T3-L1 cells, and to evaluate the
potential of APS for clinical diabetes treatment.
Materials and methods
Cell culture
3T3-L1 adipocytes were inoculated in high-glucose
Dulbecco’s modified Eagle medium (DMEM; Hyclone Co., Thermo Fisher
Scientific, Waltham, MA, USA) containing 100 g/l foetal calf serum
at 37°C and in 5% CO2, in accordance with a modified
Delex method. Following contact inhibition for 2 days, high-glucose
DMEM (Hyclone Co.) containing 10 mg/l insulin, 200 pmol/l T3, 10
μg/ml transferrin and 0.5 mmol/l 3-isobutyl-1-methlyxanthine foetal
bovine serum was used for further culture, until mature adipocytes
were induced. Adipocytes were collected and assigned to
intervention groups upon reaching ~90% differentiation. The present
study was approved by the Ethics Committee of The Tianjin
Prevention and Treatment Center of Occupational Diseases.
Determination of cell viability
Five drops of mixed cultured cell suspension and
five drops of 0.4% trypan blue solution were mixed. Cells were then
counted using a haemocytometer (XB-K25, Shanghai Qiujing
biochemical reagent and Instrument Co., Ltd., Shanghai, China) for
2 min, and blue-stained dead cells were observed. The rate of
living cells was calculated as the ratio of living cells to the
total number of cells.
Cell growth curve
The cell suspension was seeded at a density of
1×105/ml in 24-well plates and randomly divided into
eight groups. The total cell number in each group was calculated,
and subsequently the mean was calculated from measurements obtained
from three wells. The cell counting method was based on previous
cell biology experiments (8).
Oil Red O staining
Following removal of the medium, 3T3-L1 cells were
washed three times with phosphate-buffered saline (PBS), fixed in
10% formalin solution for 30 min, then washed twice with PBS and
dip-stained with 5 ml Oil Red O working solution for 10 min. The
resultant orange fat droplets were observed using a microscope
(Olympus BX51WI-DPMC; Olympus Corporation, Tokyo, Japan). The Oil
Red O working solution was subsequently removed, and the 3T3-L1
cells were washed two times and re-dyed with haematoxylin for 10
min prior further images being obtained. This method simply and
rapidly reflects pre-adipocyte conversion rates cultured in
vitro. It has been demonstrated that the accuracy and
sensitivity of this method are similar to those observed for the
determination of marker enzyme (glycerol phosphate dehydrogenase)
during the differentiation of pre-adipocytes (9). Thus, the present method is frequently
used to identify the differentiation of pre-adipocytes cultured
in vitro.
Evaluation of adiponectin and IL-6
levels
Following identification, 3T3-L1 cells were seeded
in six-well plates at a density of 1×106 and assigned to
either the APS intervention groups or the control group. The final
concentrations of APS (American Generalisation Pharmaceutical
Company, Stanford University Science Park, Stanford, CA, USA) were
10, 0.1, 0.001 and 0 μg/μl for the high- moderate- and
low-concentration groups respectively. Following incubation at 37°C
and in 5% CO2 for 48 h, the supernatant was collected
for drug testing. Concentrations of adiponectin and IL-6 were
measured using enzyme-linked immunosorbent assay in accordance with
the kit instructions (Shanghai Chuanfu Biotech Co., Ltd., Shanghai,
China).
Statistical analysis
All experimental data are expressed as the mean ±
standard deviation. Statistical analyses were performed using SPSS
10.0 statistical software (SPSS, Inc., Chicago, IL, USA).
Heterogeneous variance data were corrected by t-test, and
homogeneous variance data were analyzed using one-way analysis of
variance, and the abnormal distribution data using the Spearman
correlation coefficient formula. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
Morphological observation
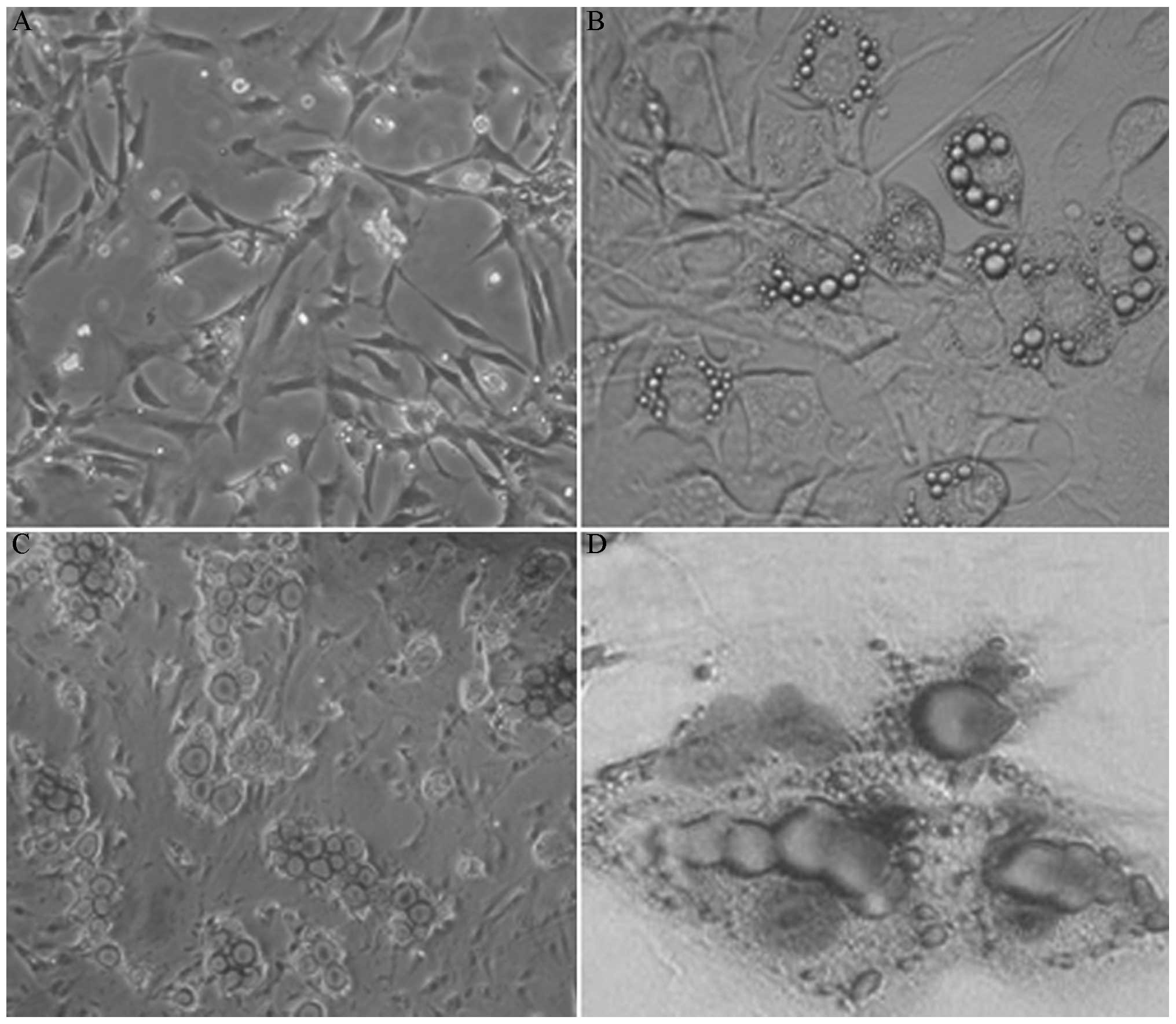
The majority of the inoculated 3T3-L1 cells
exhibited characteristic shuttle shapes with fibroblast-like
growth, round nuclei and sustained proliferation (Fig. 1A). Four days after induction, lipid
droplets appeared within the 3T3-L1 cells. Droplets were initially
focused at the cell periphery; however, the droplets gradually
increased in size and spread throughout the cell. The cells
subsequently became more round in morphology. Cells of various
sizes containing fat droplets were formed after ~10 days (Fig. 1B) or integrated into large lipid
droplets. The number of cells containing lipid droplets gradually
increased with time. Following Oil Red O and haematoxylin staining,
lipid droplets were stained red and nuclei were stained blue
(Fig. 1C and D).
Cell viability
The rate of living cells was ≥90%, indicating the
high viability of cells obtained from enzyme digestion. These cells
were thus used for subsequent culturing and monitoring.
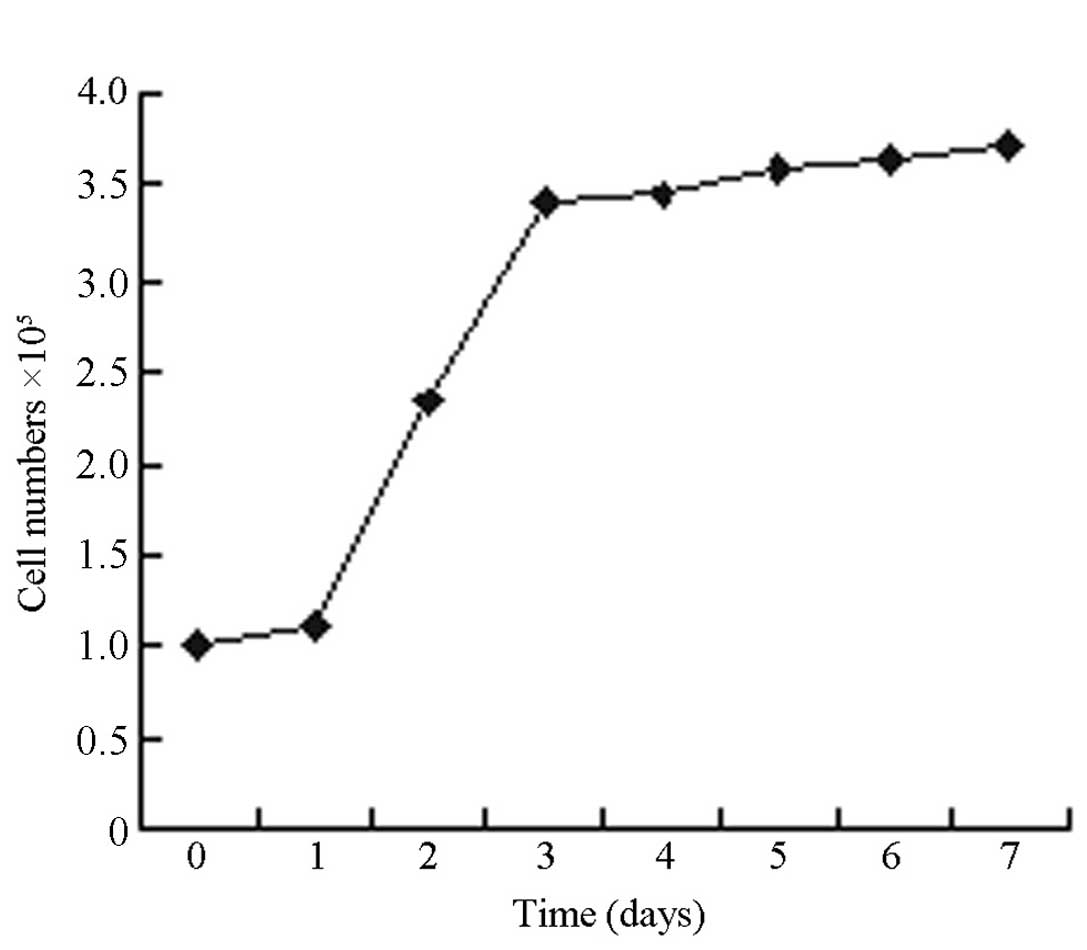
Cell growth curve
Fig. 2 shows that
the growth of 3T3-L1 cells formed an S-shaped curve. Proliferation
exhibited a pattern of latency, followed by a period of logarithmic
growth, then a plateau. The 3T3-L1 cells began to proliferate 24 h
after seeding, and the number of cells increased to ~3.5 fold after
72 h. Consequently, the initial 72 h was considered to be the
logarithmic growth period, during which the most rapid
proliferation occurred. The 3T3-L1 cells entered the plateau phase
on the fourth day after seeding. Using the Patterson formula,
Td=ΔT × lg2/(lgNt/lgN0), (where
Td, doubling time; ΔT, time interval; Nt, the
end point cell number; N0, the initial cell number) the
population doubling time of 3T3-L1 cells in the logarithmic growth
period was determined to be 41 h. Therefore, the second to fourth
day of 3T3-L1 cell proliferation was considered to be the critical
period of induction.
Adiponectin and IL-6 secretion following
APS intervention
Levels of adiponectin and IL-6 secretion by 3T3-L1
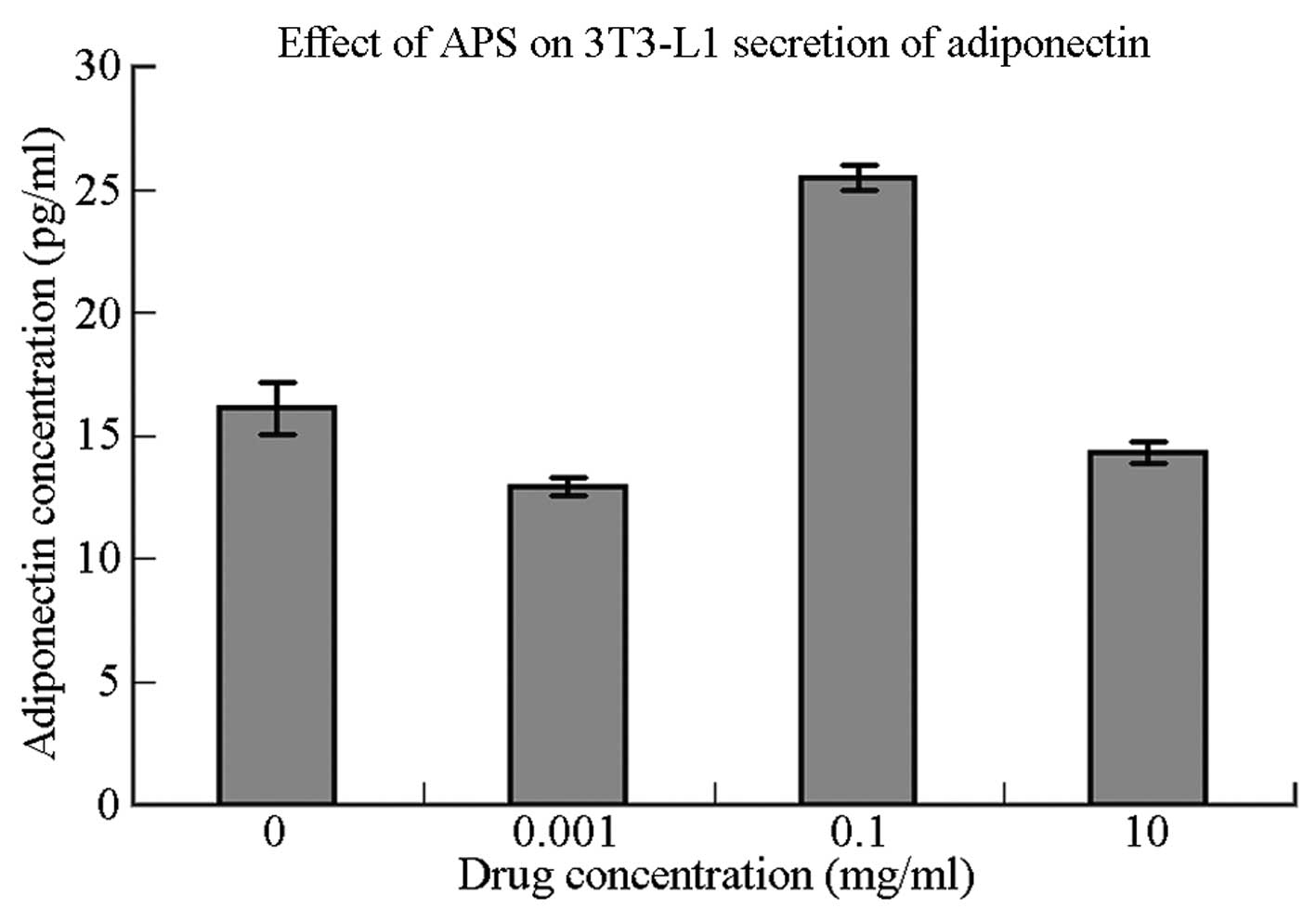
cells following stimulation with APS are shown in Table I. In comparison with the control
group, the 3T3-L1 cells in the moderate-concentration group (APS,
0.1 μg/μl) demonstrated significantly increased adiponectin
secretion (P=0.001). 3T3-L1 cells in the low-concentration group
(APS, 0.001 μg/μl) showed significantly reduced adiponectin
secretion (P<0.05), whereas the 3T3-L1 cells in the
high-concentration group (APS, 10 μg/μl) demonstrated no
significant reduction in adiponectin secretion (P>0.05) compared
with the control group.
 | Table IEffects of various concentrations of
APS on IL-6 and adiponectin secretion by 3T3-L1 cells. |
Table I
Effects of various concentrations of
APS on IL-6 and adiponectin secretion by 3T3-L1 cells.
| Concentration of APS
(μg/μl) | n | Concentration of IL-6
(pg/ml) | n | Concentration of
adiponectin (ng/ml) |
|---|
| 0 (Control
group) | 6 | 125.55±12.34 | 6 | 16.12±1.06 |
| 0.001 | 6 | 100.56±11.60 | 6 | 12.92±0.37a |
| 0.1 | 6 | 50.734±4.42a | 6 | 25.44±0.51a |
| 10 | 6 | 38.488±4.86a | 6 | 14.31±0.42 |
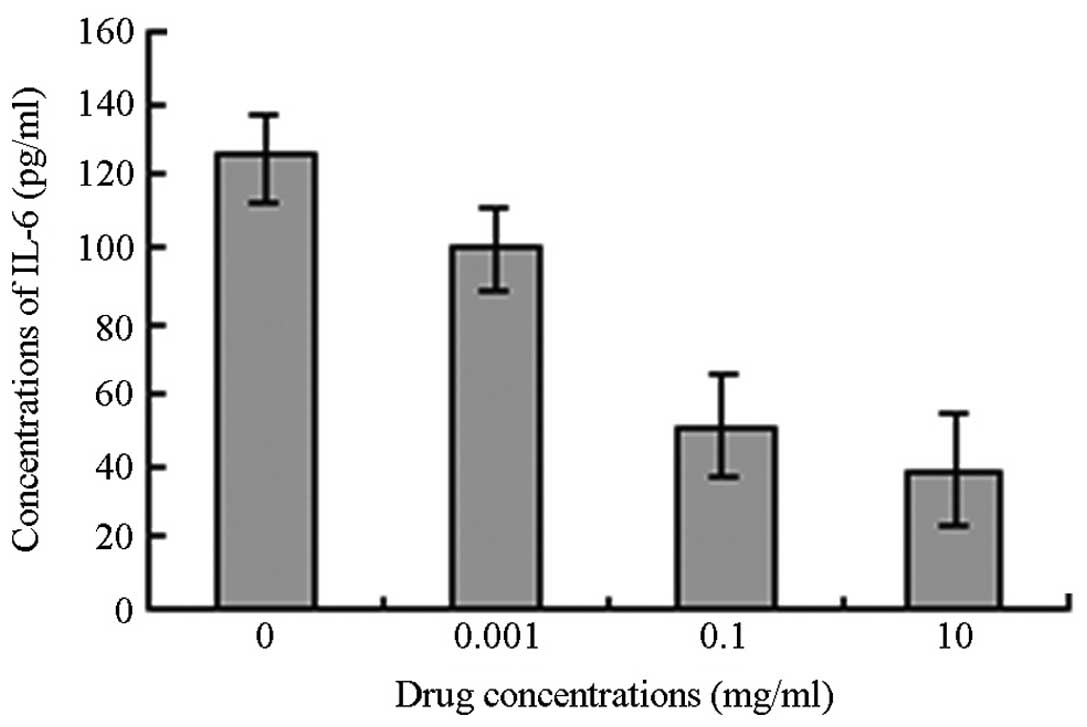
As the concentration of APS increased, the level of
IL-6 secretion by the 3T3-L1 cells was observed to decrease. While
the decrease in IL-6 secretion in the low-concentration group was
not considered statistically significant (P>0.05), the decreases
in the moderate- and high-concentration groups were considered
statistically significant compared with the IL-6 secretion by the
3T3-L1 cells in the control group (P<0.05) (Figs. 3 and 4).
Discussion
IR refers to the phenomenon wherein the body
exhibits a reduced biological response to insulin. Reductions in
the biological effect of each unit of insulin in insulin effector
organs result in a decrease in glucose uptake and elimination in
surrounding tissues, including the liver, skeletal muscle and
adipose tissues. As a pathological state, IR has a significant
association with metabolic syndrome and is closely associated with
obesity (10).
Found in Mongolia, A. mongholicus was first
recorded in ‘Shen Nong’s Herbal Classic’ as a top-grade herbal
plant that is sexual Gan, tepid. A. mongholicus is
beneficial for the spleen and lungs and has a number of functions,
inluding lifting yang, tonifying qi and strengthening exterior
(11). APS are water-soluble
macromolecular compounds with potent biological activities, and may
be extracted from A. mongholicus. APS exert various effects,
including oxygen free radical scavenging, lipid peroxidation
reduction and immune regulation, and exhibit antitumour and
anti-ageing properties (12).
In recent years, a number of studies have focused on
the endocrine function of adipose tissue for energy-storage organs.
Adipose tissue is capable of secreting numerous bioactive
substances involved in various metabolic processes, including
tumour necrosis factor α, plasminogen activator inhibitor-1, leptin
and IL-6. 3T3-L1 cells are frequently used as a model for studying
the endocrine function of adipocytes. Adiponectin, a lipid-derived
plasma protein specifically secreted by fat cells, has been
indicated to improve IR and exert anti-atherosclerotic and
anti-inflammatory effects (13).
However, the molecular mechanisms underlying the role of
adiponectin in the treatment of diabetes (14) remain unclear.
In vivo and in vitro experiments have
shown that the expression of adiponectin and PPAR-γ is closely
associated with fat-specific nuclear receptors. Decreases in PPAR-γ
transcriptional activity increase inflammatory cytokine secretion
and decrease levels of adiponectin, resulting in a decrease in
insulin sensitivity (15). mRNA
expression and adiponectin secretion in humans, obese (ob/ob) mice
and cultured 3T3-L1 cells increase following treatment with PPAR-γ
agonists (16–19). PPAR belongs to the hormone nuclear
receptor superfamily and is composed of three subtypes encoded by
three different genes: PPAR-α, -δ and -γ. PPAR-γ is one of the most
adipose tissue-specific nuclear transcription factors with
important functions in the proliferation and differentiation of fat
cells. PPAR-γ has also been found to increase the number of insulin
receptors in fat cell membranes, thereby upregulating adiponectin
expression (17).
Thiazolidinedione (TZD) is a synthetic ligand of
PPAR-γ, and is traditionally used for the treatment of IR. TZD
promotes glucose transport, increases adipose tissue insulin
sensitivity and is used to alleviate bone loss, fluid retention and
certain liver toxicities. Diabetes treatment guidelines worldwide
recommend that patients decrease their sugar intake, and avoid
weight gain and the problems associated with low blood sugar.
However, the potential of TZD may be limited, due to its
association with obesity and IR.
Given recent developments in Traditional Chinese
Medicine and molecular biology, numerous Chinese medicinal
ingredients have been identified to demonstrate functional
similarity to PPAR agonists without significant side-effects in
humans. Previous studies have demonstrated that APS increase PPAR-γ
mRNA expression and promote cell differentiation, similar to TZDs.
Furthermore, adiponectin secretion has been found to increase
following PPAR-γ activation. In the present study, following
stimulation with 0.1 μg/μl APS, 3T3-L1 adipocytes demonstrated
significant increases in adiponectin secretion compared with the
control group (P<0.01). These results are consistent with the
effective concentrations of APS necessary for stimulation of PPAR-γ
mRNA expression in fat cells reported in a previous study (2). Following stimulation with high and
low concentrations of APS, adiponectin secretion decreased compared
with that in the control group (P>0.05 and P<0.05,
respectively), indicating that optimal APS concentrations are
required to obtain the desired effect, or regulatory mechanisms
that lead to a reduction in adiponectin secretion occur. A previous
study suggested that APS are capable of enhancing the activity of
the insulin receptor and downstream molecules of
phosphatidylinositol 3-kinase (PI3K)-protein kinase B (PKB)
(20), and that the sustained
activation of P13K-PKB may reduce the expression of adiponectin
proteins in fat cells (21).
In addition to APS, other inflammatory factors,
including IL-6, have been shown to affect adiponectin secretion.
IL-6 is an inflammatory cytokine with endocrine characteristics
closely associated with adiponectin. Approximately one-third of
IL-6 in the body is secreted by fat cells. Plasma IL-6 levels
increase during IR associated with obesity and diabetes; therefore,
IL-6 plasma concentration may represent an independent predictor of
risk for T2DM (22). Endogenous
IL-6 may interact with adiponectin through paracrine and autocrine
transport mechanisms, and it has been demonstrated that IL-6
produced by local adipose tissue directly inhibits local
adiponectin production (23,24).
In this study, rat 3T3-L1 adipocytes exhibited a
significant concentration-dependent decrease in IL-6 secretion when
treated with APS compared with the control group (P<0.01).
However, levels of IL-6 secretion were not significantly correlated
with adiponectin secretion (R=−0.13). This was inconsistent with
the findings of Conroy (25), who
identified a negative correlation between IL-6 and adiponectin. Two
reasons may explain such contrasting findings: Firstly, differences
between in vivo and in vitro environments may
influence such contrasting results. For example, in an in
vitro environment, differences in co-stimulatory factors, cell
sources and different external stimuli are eliminated. Secondly,
adiponectin secretion may be affected by multiple factors, among
which IL-6 is only one. The reduced IL-6 concentrations induced by
APS may represent a mechanism by which IR may be improved. Rotter
et al (26) found that IL-6
is capable of regulating insulin transduction signals and the
development of IR, by inhibiting IRS-1, glucose transporter type 4
and PI3K in cultured 3T3-L1 adipocytes (27,28).
APS are considered to intervene in such pathways indicating that
APS may improve IR through multiple mechanisms.
In conclusion, in vitro experiments
demonstrated that optimal concentrations of APS are capable of
reducing secretion of the inflammatory cytokine IL-6 and increasing
secretion of the protective factor adiponectin, suggesting a
potential mechanism by which APS may ameliorate IR in the human
body. These findings may indicate a novel therapeutic approach for
the clinical treatment of diabetes, using APS. However, no
significant correlation was observed between APS and adiponectin
levels. Additional factors that may affect adiponectin secretion,
including the function and status of IL-6 and regulation by other
protein factors, require further study. Previous studies have shown
that APS affect the adenosine monophosphate-activated protein
kinase signalling (29) and
upstream inflammatory factor (30)
systems. Further investigations into the physiological mechanisms
of APS are required for the development of novel drugs for diabetes
treatment.
Acknowledgements
This study was supported by a grant from the
Department of Science, Henan, P.R China (224630170).
Abbreviations:
|
IR
|
insulin resistance
|
|
APS
|
Astragalus polysaccharides
|
|
IL-6
|
interleukin-6
|
|
IRS-1
|
insulin receptor substrate 1
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
PKB
|
protein kinase B
|
|
NF-κB
|
nuclear factor-κ-light-chain-enhancer
of activated B cells
|
|
PPAR
|
peroxisome proliferator-activated
receptor
|
References
|
1
|
Ferrannini E, Balkau B, Coppack SW, et al;
RISC Investigators. Insulin resistance, insulin response, and
obesity as indicators of metabolic risk. J Clin Endocrinol Metab.
92:2885–2892. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Haffner SM, D’Agostino R Jr, Mykkänen L,
et al: Insulin sensitivity in subjects with type 2 diabetes.
Relationship to cardiovascular risk factors: the Insulin Resistance
Atherosclerosis Study. Diabetes Care. 22:562–568. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tanti JF, Ceppo F, Jager J and Berthou F:
Implication of inflammatory signaling pathways in obesity-induced
insulin resistance. Front Endocrinol (Lausanne).
3:1812013.PubMed/NCBI
|
|
4
|
Fresno M, Alvarez R and Cuesta N:
Toll-like receptors, inflammation, metabolism and obesity. Arch
Physiol Biochem. 117:151–164. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zoico E, Di Francesco V, Olioso D, et al:
In vitro aging of 3T3-L1 mouse adipocytes leads to altered
metabolism and response to inflammation. Biogerontology.
11:111–122. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weng XG, Bai LW, Liu HZ, Wang T and Chen
LL: Effects of Astragalus polysaccharides on expression of
adiponectin in 3T3-L1 adipocytes. J Appl Clin Pediatr.
24:1598–1600. 2009.(In Chinese).
|
|
7
|
Zhao M, Zhang ZF, Ding Y, Wang JB and Li
Y: Astragalus polysaccharide improves palmitate-induced insulin
resistance by inhibiting PTP1B and NF-κB in C2C12 myotubes.
Molecules. 17:7083–7092. 2012.PubMed/NCBI
|
|
8
|
Zhang JB: Medical Cell Biology experiments
guidance and Exercises. Beijing: People’s Medical Publishing House;
2005, pp. 107
|
|
9
|
Ramírez-Zacarías JL1, Castro-Muñozledo F
and Kuri-Harcuch W: Quantitation of adipose conversion and
triglycerides by staining intracytoplasmic lipids with Oil red O.
Histochemistry. 97:493–497. 1992.PubMed/NCBI
|
|
10
|
Cheal KL, Abbasi F, Lamendola C,
McLaughlin T, Reaven GM and Ford ES: Relationship to insulin
resistance of the adult treatment panel III diagnostic criteria for
identification of the metabolic syndrome. Diabetes. 53:1195–1200.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao XM: Traditional Chinese Pharmacology.
Beijing: Traditional Chinese Medicine Publishing House; 2011, pp.
428
|
|
12
|
Wu M and Tan R: Advance in study on
astragalus polysaccharide. Journal of North Sichuan Medical
College. 28:17–22. 2013.(In Chinese).
|
|
13
|
Cui J, Panse S and Falkner B: The role of
adiponectin in metabolic and vascular disease: a review. Clin
Nephrol. 75:26–33. 2011.PubMed/NCBI
|
|
14
|
Phillips SA and Kung JT: Mechanisms of
adiponectin regulation and use as a pharmacological target. Curr
Opin Pharmacol. 10:676–683. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lavrenko AV, Shlykova OA, Kutsenko LA,
Mamontova TV and Kaĭdashev IP: Pharmacogenetic features of the
effect of metformin in patients with coronary heart disease in the
presence of metabolic syndrome and type 2 diabetes mellitus in
terms of PPAR-gamma2 gene polymorphism. Ter Arkh. 84:35–40.
2012.(In Russian).
|
|
16
|
Lefils-Lacourtablaise J, Socorro M, Géloën
A, et al: The eicosapentaenoic acid metabolite
15-deoxy-δ(12,14)-prostaglandin J3 increases adiponectin secretion
by adipocytes partly via a PPARγ-dependent mechanism. PLoS One.
8:e639972013.PubMed/NCBI
|
|
17
|
Kubota N, Terauchi Y, Kubota T, et al:
Pioglitazone ameliorates insulin resistance and diabetes by both
adiponectin-dependent and -independent pathways. J Biol Chem.
281:8748–8755. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tishinsky JM, Ma DW and Robinson LE:
Eicosapentaenoic acid and rosiglitazone increase adiponectin in an
additive and PPARγ-dependent manner in human adipocytes. Obesity
(Silver Spring). 19:262–268. 2011.PubMed/NCBI
|
|
19
|
Bak EJ, Park HG, Kim JM, Kim JM, Yoo YJ
and Cha JH: Inhibitory effect of evodiamine alone and in
combination with rosiglitazone on in vitro adipocyte
differentiation and in vivo obesity related to diabetes. Int J Obes
(Lond). 34:250–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu M, Ouyang JP, Wu K, et al: Effect of
Astragalus polysaccharide on Ser phosphorylation of protein kinase
B in skeletal muscle of KKAy mice. Medical Journal of Wuhan
University. 27:135–139. 2006.(In Chinese).
|
|
21
|
Gao P, Li Q, Sun YZ and Zhang JC:
Continuously activated Akt inhibits adiponectin secretion in 3T3-L1
adipocytes. Chinese Journal of Diabetes. 14:142–143. 1452006.(In
Chinese).
|
|
22
|
Goldberg RB: Cytokine and cytokine-like
inflammation markers, endothelial dysfunction, and imbalanced
coagulation in development of diabetes and its complications. J
Clin Endocrinol Metab. 94:3171–3182. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Phillips CM and Perry IJ: Does
inflammation determine metabolic health status in obese and
nonobese adults? J Clin Endocrinol Metab. 98:E1610–E1619. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei SS and Liang DD: The effect of
exercises on TNF-alpha, IL-6 and adiponectin in different fat diet
rats. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 29:280–282. 2013.(In
Chinese).
|
|
25
|
Conroy SM, Chai W, Lim U, Franke AA,
Cooney RV and Maskarinec G: Leptin, adiponectin, and obesity among
Caucasian and Asian women. Mediators Inflamm. 2011:2535802011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rotter V, Nagaev I and Smith U:
Interleukin-6 (IL-6) induces insulin resistance in 3T3-L1
adipocytes and is, like IL-8 and tumor necrosis factor-alpha,
overexpressed in human fat cells from insulin-resistant subjects. J
Biol Chem. 278:45777–45784. 2003. View Article : Google Scholar
|
|
27
|
Liu HF, Ren YH, Han ZX, et al: Effect of
astragalus polysaccharides on insulin resistance and gene
expression of GLUT4 in type 2 diabetes mellitus rats. Chinese
Journal of Gerontology. 31:3988–3989. 2011.(In Chinese).
|
|
28
|
Xu Y, Wang BH, LI K, et al:
Insulin-sensitization of Astragalus polysaccharide and its effect
on protein tyrosine phosphatase 1B. Medical Journal of Wuhan
University. 3:288–291. 2010.(In Chinese).
|
|
29
|
Hu YQ, Li J, Liu J, Ou-Yang JP and Song J:
APS improves free fatty acid metabolism by activating AMPK and
promoting translocation of FAT/CD36 in C2C12 myoblasts. Chin J
Pathophysiol. 29:637–640. 2013.(In Chinese).
|
|
30
|
Liang LJ, Wang HX, Sun XF, et al:
Astragalus polysaccharides ameliorate hypertrophy of
cardiomyocytes induced by tumor necrosis factor-α via TLR4/NF-κB
signaling pathway. Chin J Pharmacol Toxicol. 27:168–173. 2013.(In
Chinese).
|