Introduction
Myelodysplastic syndrome (MDS) is a clinically and
cytogenetically heterogeneous group of clonal diseases
characterized by ineffective hematopoiesis-associated cytopenias,
consequent bleeding, infections and a high risk of acute myeloid
leukemia (AML) transformation (1,2). The
International Prognostic Scoring System (IPSS) for MDS is based
upon weighted data on the bone marrow blast percentage, cytopenia
and cytogenetics and it separates patients into four prognostic
groups: Low, intermediate-1, intermediate-2 and high. The median
overall survival time of patients with high-risk MDS (commonly
defined as the patients with an IPSS risk score >1.0) is
approximately 14 months (3). These
patients have low remission rates and short periods of disease-free
survival despite chemotherapy (4–6). MDS
occurs more frequently in older individuals. Currently, there is no
curative treatment for MDS, with the exception of allogenic stem
cell transplantation which is unsuitable for the majority of
elderly patients due to comorbid illness or poor performance
status. Therefore, elderly patients generally receive low-dose
chemotherapy, supportive care or investigational treatment
(2,7). The general view is that complete or
partial remission is a prerequisite for the prolonged survival of
patients with high-risk MDS and it is achieved using chemotherapy
regimens similar to those used for AML (8). However, traditional chemotherapies
for MDS have limited success rates, so it is necessary to explore
novel therapeutic targets and agents that have higher selectivity
for tumor cells and less toxicity toward normal tissues.
Heat shock protein 90 (Hsp90) is attractive
molecular target as it acts as a chaperone that prevents the
degradation of a number of important cellular oncoproteins,
including receptor and nonreceptor kinases (9). The overexpression of Hsp90 in acute
leukemia cells has been confirmed by several studies (10,11).
Furthermore, the expression levels of Hsp90 are higher in blastic
MDS, which is associated with poor prognosis (12,13).
Hsp90 inhibitory agents, including the ansamycin antibiotic
geldanamycin, bind to the ATP-binding pocket of Hsp90, thereby
disrupting Hsp90 function, and thus present as promising drugs for
the treatment of cancer (14).
Phase I/II clinical trials of Hsp90 inhibitors have been conducted,
including a Phase I trial of the Hsp90 inhibitor tanespimycin
(17-AAG) in relapsed and refractory acute leukemia (15,16).
BIIB021 was the first ‘fully synthetic’ Hsp90
inhibitor to be used for the clinical treatment of solid tumors and
hematological malignancies (17,18).
BIIB021 induces the degradation of Hsp90 client proteins, including
human epidermal growth factor receptor-2 (HER-2), Akt and RAF
proto-oncogene serine/threonine-protein kinase (Raf-1), and results
in tumor growth inhibition (18).
A phase II clinical trial (19)
showed that BIIB021 improves the outcome of patients with
gastrointestinal stromal tumors refractory to imatinib and
sunitinib. Despite a broad prospect for further clinical
application of this agent, no studies have been conducted using MDS
cells. The present study focused on the therapeutic effects and
mechanisms of one molecularly targeted agent, the Hsp90 inhibitor
BIIB021, on high-risk MDS in vitro.
Methods and materials
Cell culture and reagents
SKM-1 cells (JCRB0118; Japanese Collection of
Research Bioresources Cell Bank, Osaka, Japan) were cultured in
RPMI-1640 medium (Gibco, Grand Island, NY, USA) with 10% fetal
bovine serum (Gibco) at 37°C in a humidified atmosphere of 5%
CO2. BIIB021 was purchased from Selleck Chemicals
(Houston, TX, USA). Methylcellulose and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). All
antibodies used in the western blot analysis were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA), with the
exception of human anti-β-actin. Insulin-like growth factor-1
(IGF-1) was purchased from Peprotech (Rocky Hill, NJ, USA). The
caspase inhibitors z-IETD-fmk and z-LEHD-fmk were purchased from
Biovision (Palo Alto, CA, USA).
Cell viability assay
To evaluate the effects of BIIB021 on the MDS cells,
MTT assays were performed. The SKM-1 cells were cultured at a
density of 5×104 cells/well in a 96-well plate and
treated with BIIB021 at concentrations of 50, 100, 200 and 400 nM,
respectively. Following 24 or 48 h incubation, MTT was added to
each well and the plates were incubated for an additional 4 h at
37°C. The supernatant was removed, followed by the addition of 200
μl dimethyl sulfoxide (Amresco, Solon, OH, USA). The absorbance at
a wavelength of 570 nm was detected with an enzyme-linked
immunosorbent assay plate reader (Bio-Rad, Hercules, CA, USA). Each
assay was performed three times in triplicate.
Annexin V binding assay
The cells were cultured at a density of
5×104 cells/well in a six-well plate and treated for 24
h with BIIB021 at concentrations of 0, 100, 200 and 400 nM. After
24 h of treatment at 37°C, the cells were collected and washed.
Aliquots of the cells were resuspended in 500 μl binding buffer and
stained with 5 μl Annexin V-fluorescein isothiocyanate (FITC) and 5
μl propidium iodide (PI; Biouniquer Technology Co., Ltd., Suzhou,
China) for 15 min in the dark, and examined by flow cytometry. Data
acquisition and analysis were performed on a FACSCalibur flow
cytometer (Becton-Dickinson, Franklin Lakes, USA) using CellQuest
software (Becton-Dickinson).
Cell cycle analysis
The cells were treated with BIIB021 at
concentrations of 0, 100 and 200 nM and incubated for 24 h at 37°C.
The cells were harvested, washed twice with cold phosphate-buffered
saline (PBS), and suspended and fixed in 75% ice-cold ethanol
overnight at 4°C. Subsequently, the sample was washed with PBS and
incubated with 250 μg/ml RNase A and 10 μg/ml PI for 30 min. The
cells were analyzed using a FACSCalibur flow cytometer.
Hoechst 33258 DNA staining
To detect the morphological changes following
treatment with BIIB021 (0, 100, 200 and 400 nM), SKM-1 cells were
plated at an initial density of 1×105 cells/well in a
24-well plate and treated with BIIB021 for 24 h. The treated cells
were fixed with 3.7% paraformaldehyde for 30 min and then stained
with Hoechst 33258 (0.5 μg/ml) for 20 min at room temperature. The
cells were counted under an Axiovert fluorescence microscope (Carl
Zeiss, Göttingen, Germany) with an excitation wavelength of 350 nm
and an emission wavelength of 460 nm.
Western blot analysis
The cells were harvested 24 h after treatment at the
indicated doses and times, and the cell lysates were subjected to
western blotting, performed as described previously (20). Briefly, the cells were collected
and lysed using 10 mM Tris, 1 mM ethylenediaminetetraacetic acid,
10 mM KCl and 0.3% Triton X-100 (pH 7.9). The concentration of the
protein samples was measured by the Bradford method. The protein
samples were separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and then electroblotted onto Hybond-P
polyvinylidene fluoride membranes (Amersham, Piscataway, NJ, USA).
The membranes were subjected to western blot analysis with primary
antibodies to caspase-8, -9 and -3, poly(ADP-ribose) polymerase
(PARP), p110δ, Akt, phospho-Akt, p65, phospho-p65, cyclin-dependent
kinase (CDK)4, CDK6, cyclin D1 and β-actin (Sigma-Aldrich). The
secondary antibodies used in this study were provided by
Multisciences Co., Ltd. (Hangzhou, China).
Statistical analysis
Experimental results are statistically presented as
the mean ± standard deviation. Data were analyzed by one-way
analysis of variance. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of BIIB021 on the viability of
the human MDS cell line
The SKM-1 cells were tested for sensitivity to
BIIB021 in a cell proliferation assay. The 50% inhibitory
concentration (IC50) values were 275.2 nM for 24 h and
163.9 nM for 48 h. As shown in Fig.
1A, BIIB021 effectively inhibited SKM-1 cell growth in a
concentration- and time-dependent manner. The Annexin V binding
assay confirmed that BIIB021 induced SKM-1 cell apoptosis in a
concentration-dependent manner (Fig.
1B). Morphological observation by Hoechst 33258 DNA staining
showed an increased number of cells with nuclear condensation and
fragmentation following treatment with BIIB021 for 24 h (Fig. 1C).
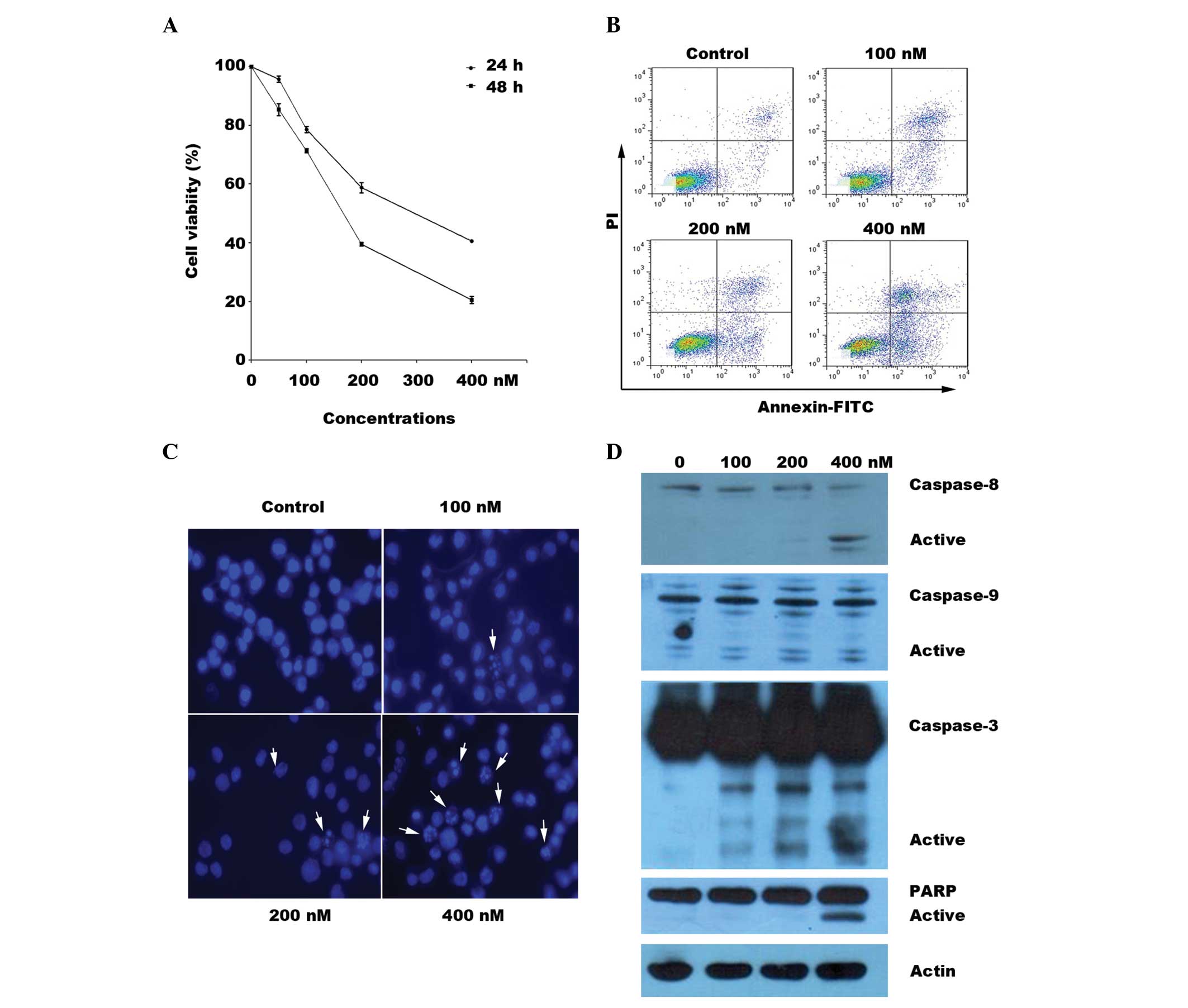 | Figure 1BIIB021 inhibits cellular viability
and induces apoptosis in SKM-1 cells. (A) After 24 and 48 h
treatment with BIIB021 (50, 100, 200 and 400 nM), the cells were
incubated with MTT to determine the levels of cell proliferation.
The data shown are the mean ± SD of three independent experiments.
(B) The SKM-1 cells were treated with BIIB021 at the indicated
concentrations for 24 h and processed for Annexin V-FITC and PI
double staining. The apoptotic cells were then quantitatively
monitored. (C) Followng a treatment similar to that in (B), the
nuclear morphology of the SKM-1 cells as analyzed by Hoechst 33258
staining. The arrows indicate apoptotic nuclei. (D) Cell lysates
were prepared from SKM-1 cells following incubation with or without
BIIB021 at the indicated concentrations for 24 h. Equal amounts of
proteins per sample were resolved by SDS-PAGE and then transferred
to a PVDF membrane, which was probed for the expression levels of
caspase-3, -8 and -9, PARP and β-actin. PI, propidium iodide; FITC,
fluorescein isothiocyanate; PARP, poly(ADP-ribose) polymerase; MTT,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide;
SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis; PVDF, polyvinylidene fluoride. |
BIIB021 induces SKM-1 cell apoptosis via
activation of the caspase family
To detect the mechanisms of BIIB021-induced cell
apoptosis in the MDS cells, western blotting was used to measure
the levels of activation of the caspase family. BIIB021 triggered
concentration-dependent cleavage of caspase-3, -8 and -9 and PARP
(Fig. 1D). Furthermore, the effect
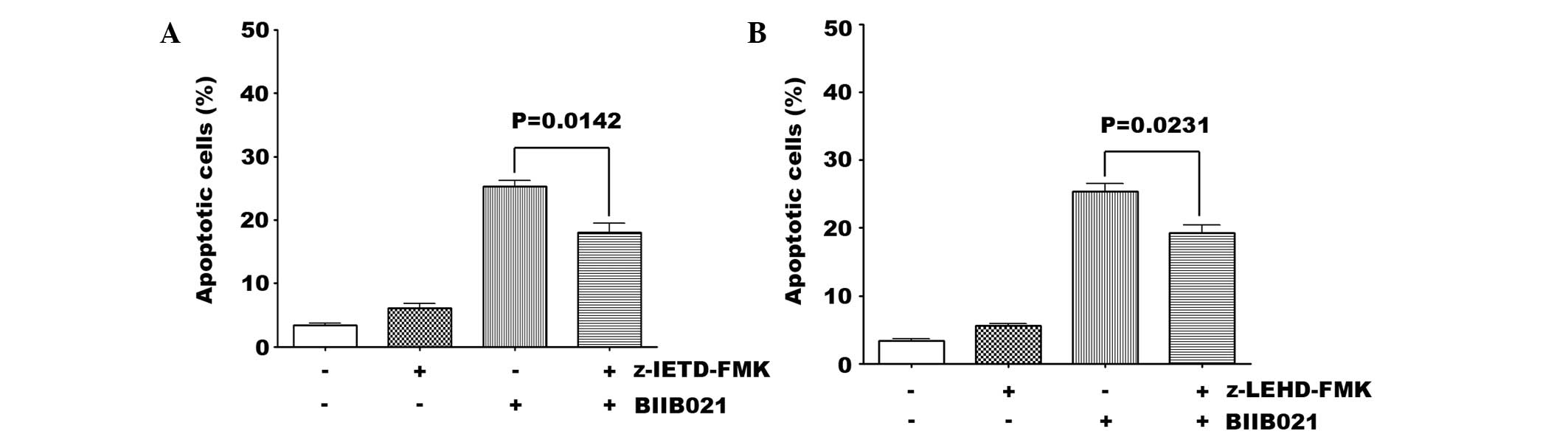
of caspase inhibitors on BIIB021-induced apoptosis was observed.
The caspase-8 (20 μM) and caspase-9 (20 μM) inhibitors partially
inhibited BIIB021-induced apoptosis (Fig. 2). These results suggest that
caspase-8 and -9 inhibitors are able to attenuate BIIB021-induced
apoptosis, which indicates that BIIB021 caused apoptosis through
activating the cascade to the caspase-8 and -9 pathways in SKM-1
cells.
BIIB021 inhibits the phosphatidylinositide 3-kinase
(PI3K)/Akt and nuclear factor (NF)-κB signaling pathways. It has
been demonstrated that the PI3K/Akt and NF-κB signaling pathways
are activated in high-risk MDS patients and activation of the
signaling pathways is responsible for the suppression of apoptosis
of MDS cells, contributing to AML transformation (21–24).
Furthermore, lasting activation of NF-κB results in drug resistance
(25). Therefore, the present
study tested whether the mechanisms of BIIB021-induced cell
apoptosis are involved in the PI3K/AKT and NF-κB signaling
pathways. Fig. 3A shows that the
protein expression levels of the PI3K isoforms p110δ, p65 and Akt
were only slightly reduced in the SKM-1 cells treated with BIIB021,
whereas BIIB021 markedly reduced the levels of phospho-Akt and
phospho-p65 in a concentration-dependent manner. Subsequently,
whether the downregulation of the levels of these proteins was
reversed by IGF-1 was examined. As shown in Fig. 3B, pretreatment of the SKM-1 cells
with 100 ng/ml IGF-1 did not attenuate the BIIB021-mediated
suppression of the protein expression, despite the fact that IGF-1
alone upregulated the expression levels of p110, p65 and Akt and of
phospho-Akt and phospho-p65.
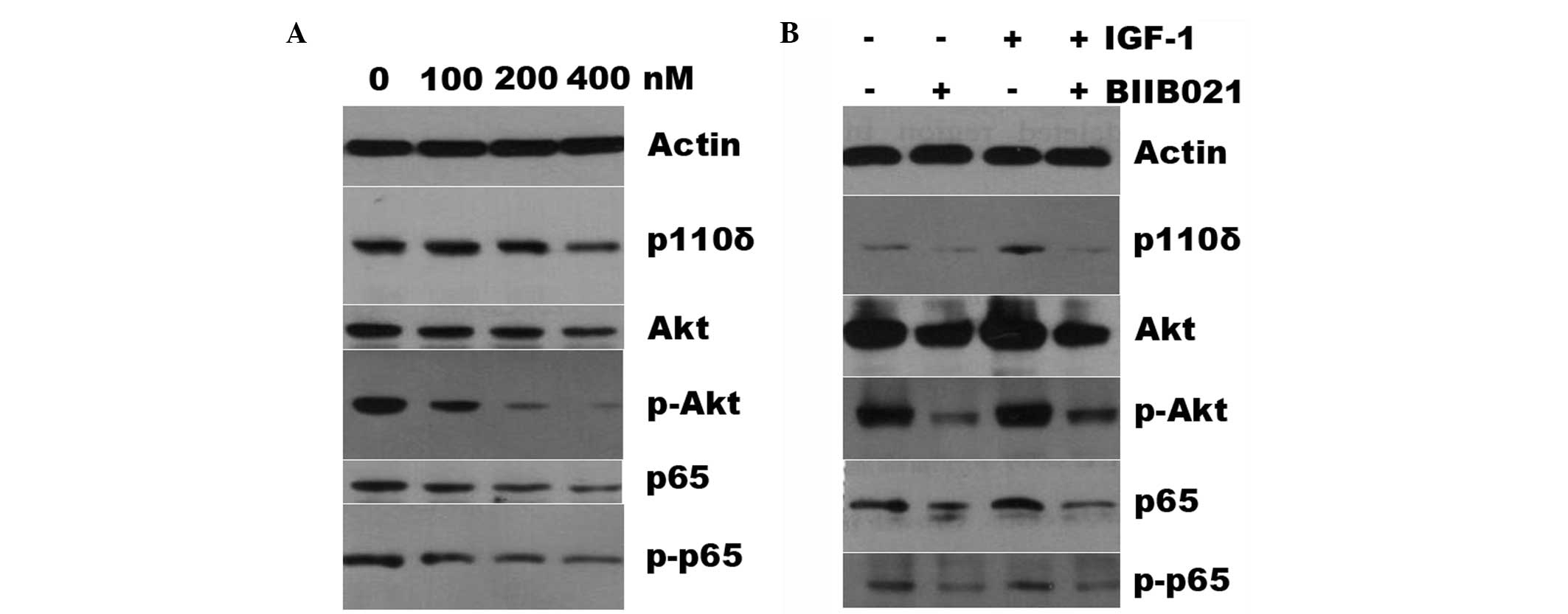 | Figure 3BIIB021 reduces the expression levels
of the PI3K/Akt and NF-κB pathway proteins, and IGF-1 does not
attenuate BIIB021-mediated inhibition of these proteins. (A) The
SKM-1 cells were treated with different concentrations (100, 200
and 400 nM) of BIIB021 for 24 h, and the total proteins were
extracted and subjected to western blot analysis using primary
antibodies for p110δ, Akt, phospho-Akt, p65 and phospho-p65. Actin
was used as a protein loading control. (B) The SKM-1 cells were
treated with or without BIIB02 (400 nM), IGF-1 (100 ng/ml), or
BIIB021 combined with IGF-1. Whole cell lysates were then subjected
to SDS-PAGE followed by immunoblotting with the antibody that
recognizes the corresponding antigens. PI3K, phosphatidylinositide
3-kinase; NF-κB, nuclear factor-κB; IGF-1, insulin-like growth
factor-1; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. |
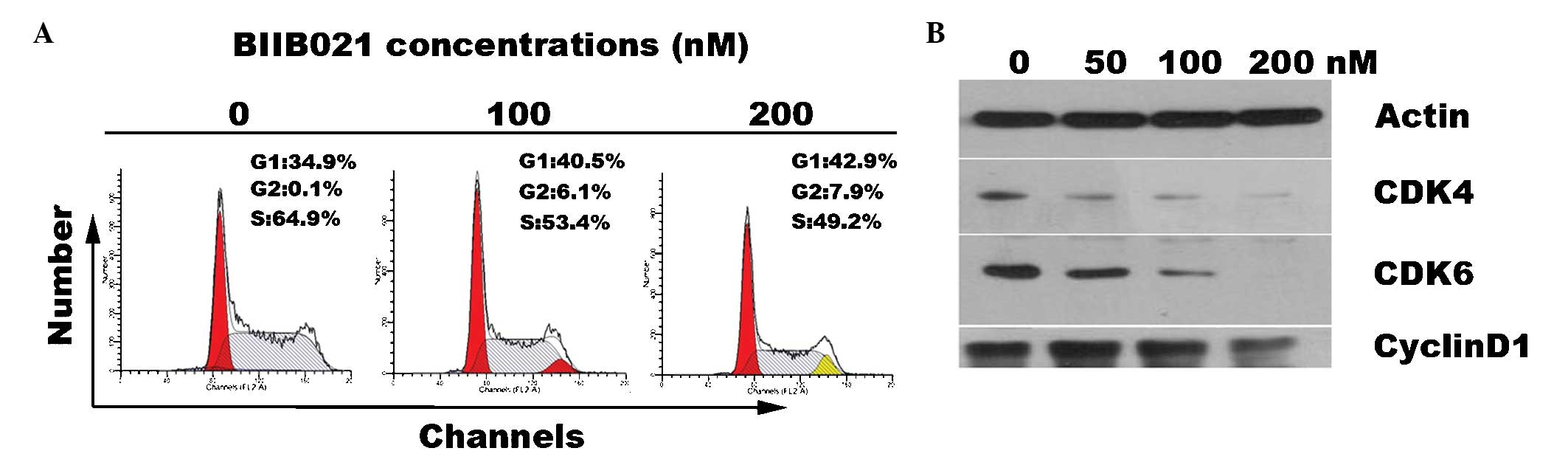
Inhibition of SKM-1 cell proliferation is
due to G0/G1 arrest via the regulation of cell cycle-related
proteins
A previous study showed that a Hsp90 inhibitor
induced cell cycle arrest at G2/M (26). On the basis of this, the effects of
BIIB021 treatment on the cell cycle were examined by DNA flow
cytometry in the present study. The results showed that, compared
with that of the untreated cells, the number of cells in the G1
phase was markedly increased in the BIIB021-treated group (34.9
versus 40.5 and 42.9% at 100 and 200 nM, respectively). In
addition, treatment with BIIB021 resulted in a reduction in the
percentage of cells in the S phase (Fig. 4A). These data suggest that BIIB021
inhibits S phase entry. It has been reported that the active
complex of CDK4/cyclin D1 allows the release of E2F transcription
factors that activate G1/S phase gene expression. In the present
study, the results indicated that BIIB021 markedly reduced the
levels of CDK4 and CDK6 in the MDS cells, but the inhibitory effect
on cyclin D1 was modest (Fig.
4B).
Discussion
Overexpression of Hsp90 has been observed in a
variety of types of cancer (27).
Previous studies have indicated the possible role of Hsp90 in MDS
pathogenesis and evolution (12,28).
For example, it has been reported that Hsp90 is overexpressed in
high-risk MDS, as compared with the levels in low-risk MDS and
normal bone marrow, and is associated with a poor prognosis for
patients (12). Certain Hsp90
inhibitors, including 17-AAG, are used as targeted therapies for
cancer as they showed anticancer activity in early clinical trials
(17,29,30).
BIIB021 has an improved pharmacological profile compared with that
of 17-AAG and other Hsp90 inhibitors, particularly with regard to
availability through chemical synthesis, metabolic stability, water
solubility and ease of administration via oral and intravenous
routes (18,31). The present study of BIIB021 shows
that the MDS cell line SKM-1 was blocked in the G1 phase of the
cell cycle and underwent apoptosis when treated with BIIB021. The
IC50 values of BIIB021 required to inhibit the growth of
the SKM-1 cells were 355.69 and 168.6 nM at 24 h and 48 h,
respectively, which is consistent with a previous study indicating
that the IC50 values of BIIB021 in various types of
solid tumor are in the range of 60 to 310 nM (32). The activation of caspase-8 and -9,
followed by the downstream activation of caspase-3 and PARP was
observed in the BIIB021-treated MDS cells in the present study.
Furthermore, caspase-8 and -9 inhibitors partially attenuated the
BIIB021-induced apoptosis. These results suggest that two main
pathways of procaspase activation (extrinsic death receptor pathway
and intrinsic mitochondrial pathway) are involved in the
BIIB021-induced apoptosis of MDS cells.
The PI3K/Akt signaling cascade represents one of the
major survival pathways that is deregulated in numerous types of
cancer and contributes to tumor pathogenesis and therapy
resistance. Constitutive activation of the PI3K/Akt signaling
pathway and NF-κB is a feature of patients with high-risk MDS
(21,33,34).
In the present study, it was identified that BIIB021 slightly
downregulated the expression levels of p110δ and Akt. Notably, the
drug markedly inhibited the phosphorylation of Akt. Marked
downregulation of the levels of p65 and phospho-p65 expression were
also observed, suggesting that BIIB021 inhibited the activation of
NF-κB, which is a downstream regulator of the PI3K/Akt signaling
pathway. As constitutive IGF-1/IGF-1 receptor (IGF-1R) signaling
contributes to deregulated PI3K activity and overexpression of
IGF-1R was observed in malignant clonal cells in bone marrow of MDS
in previous studies (35,36), the effects of exogenous IGF-1
stimulation on BIIB021-mediated inhibition of the PI3K/Akt
signaling pathways were tested in the present study. It was
observed that IGF-1 at 100 ng/ml increased the expression levels of
p110δ, phospho-Akt and phospho-p65, suggesting that IGF-1 activates
the PI3K/Akt signaling pathways in MDS cells. This result is
consistent with the evidence that IGF-1 is a strong PI3K activator
(37). However, exogenous IGF-1
stimulation did not abrogate the capacity of BIIB021 to inhibit Akt
and NF-κB. Collectively, inhibition of PI3K/Akt and NF-κB
contributes to BIIB021-induced apoptosis, implying that BIIB021 may
be useful for overcoming drug resistance.
Deregulation of the cell cycle pathway is a
contributor to the pathogenesis of MDS (38,39).
For example, cyclin D1 levels are increased in high-risk MDS,
thereby increasing the proliferation of leukemia (40,41).
BIIB021 has a high binding affinity for Hsp90 and consequently
inhibits the chaperone activity of Hsp90 and results in degradation
of the client proteins (18,41).
In the present study, in addition to the effect of G1 cell cycle
arrest, BIIB021 was demonstrated to inhibit the expression of CDK4,
CDK6 and cyclin D1, which are also Hsp90 client proteins, in MDS
cells. This effect of BIIB021 on the cell cycle pathway is similar
to that observed in other types of cancer (18).
In summary, the present study demonstrates for the
first time, to the best of our knowledge, that BIIB021 has marked
activity against MDS cells. Furthermore, BIIB021 causes the
degradation of several client proteins, including Akt, CDK4 and
CDK6 in MDS cells. Also, NF-κB activity was inhibited in the SKM-1
cells upon treatment with BIIB021 at low nanomolar concentrations.
Therefore, BIIB021 is potentially useful for clinical therapy in
the treatment of high-risk MDS.
Acknowledgements
This study was supported by a National Natural
Science Foundation of China grant (No. 81370645), the Doctoral Fund
of the Ministry of Education of China (No. 20120101110010) and
funds of the Department of Science and Technology of Zhejiang
Province (No. 2012C13021-2).
References
|
1
|
Cogle CR, Craig BM, Rollison DE and List
AF: Incidence of the myelodysplastic syndromes using a novel
claims-based algorithm: high number of uncaptured cases by cancer
registries. Blood. 117:7121–7125. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tefferi A and Vardiman JW: Myelodysplastic
syndromes. N Engl J Med. 361:1872–1885. 2009. View Article : Google Scholar
|
|
3
|
Komrokji RS, Corrales-Yepez M, Al Ali N,
et al: Validation of the MD Anderson Prognostic Risk Model for
patients with myelodysplastic syndrome. Cancer. 118:2659–2664.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sekeres MA, Schoonen WM, Kantarjian H,
List A, Fryzek J, Paquette R and Maciejewski JP: Characteristics of
US patients with myelodysplastic syndromes: results of six
cross-sectional physician surveys. J Natl Cancer Inst.
100:1542–1551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fukumoto JS and Greenberg PL: Management
of patients with higher risk myelodysplastic syndromes. Crit Rev
Oncol Hematol. 56:179–192. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gore SD and Hermes-DeSantis ER: Enhancing
survival outcomes in the management of patients with higher-risk
myelodysplastic syndromes. Cancer Control. 16(Suppl): 2–10.
2009.PubMed/NCBI
|
|
7
|
Szmigielska-Kapłon A and Robak T:
Hypomethylating agents in the treatment of myelodysplastic
syndromes and myeloid leukemia. Curr Cancer Drug Targets.
11:837–848. 2011.PubMed/NCBI
|
|
8
|
Garcia-Manero G: Treatment of higher-risk
myelodysplastic syndrome. Semin Oncol. 38:673–681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kamal A, Thao L, Sensintaffar J, Zhang L,
Boehm MF, Fritz LC and Burrows FJ: A high-affinity conformation of
Hsp90 confers tumour selectivity on Hsp90 inhibitors. Nature.
425:407–410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yufu Y, Nishimura J and Nawata H: High
constitutive expression of heat shock protein 90 alpha in human
acute leukemia cells. Leuk Res. 16:597–605. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Flandrin P, Guyotat D, Duval A, Cornillon
J, Tavernier E, Nadal N and Campos L: Significance of heat-shock
protein (HSP) 90 expression in acute myeloid leukemia cells. Cell
Stress Chaperones. 13:357–364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Duval A, Olaru D, Campos L, Flandrin P,
Nadal N and Guyotat D: Expression and prognostic significance of
heat-shock proteins in myelodysplastic syndromes. Haematologica.
91:713–714. 2006.PubMed/NCBI
|
|
13
|
Flandrin-Gresta P, Solly F, Aanei CM, et
al: Heat Shock Protein 90 is overexpressed in high-risk
myelodysplastic syndromes and associated with higher expression and
activation of Focal Adhesion Kinase. Oncotarget. 3:1158–1168.
2012.PubMed/NCBI
|
|
14
|
Didelot C, Lanneau D, Brunet M, Joly AL,
De Thonel A, Chiosis G and Garrido C: Anti-cancer therapeutic
approaches based on intracellular and extracellular heat shock
proteins. Curr Med Chem. 14:2839–2847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaufmann SH, Karp JE, Litzow MR, et al:
Phase I and pharmacological study of cytarabine and tanespimycin in
relapsed and refractory acute leukemia. Haematologica.
96:1619–1626. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jhaveri K, Taldone T, Modi S and Chiosis
G: Advances in the clinical development of heat shock protein 90
(Hsp90) inhibitors in cancers. Biochim Biophys Acta. 1823:742–755.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taldone T, Gozman A, Maharaj R and Chiosis
G: Targeting Hsp90: small-molecule inhibitors and their clinical
development. Curr Opin Pharmacol. 8:370–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lundgren K, Zhang H, Brekken J, et al:
BIIB021, an orally available, fully synthetic small-molecule
inhibitor of the heat shock protein Hsp90. Mol Cancer Ther.
8:921–929. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dickson MA, Okuno SH, Keohan ML, et al:
Phase II study of the HSP90-inhibitor BIIB021 in gastrointestinal
stromal tumors. Ann Oncol. 24:252–257. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang M, Zhang H, Liu T, Tian D, Gu L and
Zhou M: Triptolide inhibits MDM2 and induces apoptosis in acute
lymphoblastic leukemia cells through a P53-independent pathway. Mol
Cancer Ther. 12:184–194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nyåkern M, Tazzari PL, Finelli C, et al:
Frequent elevation of Akt kinase phosphorylation in blood marrow
and peripheral blood mononuclear cells from high-risk
myelodysplastic syndrome patients. Leukemia. 20:230–238.
2006.PubMed/NCBI
|
|
22
|
Yilmaz OH, Valdez R, Theisen BK, Guo W,
Ferquson DO, Wu H and Morrison SJ: Pten dependence distinguishes
haematopoietic stem cells from leukaemia-initiating cells. Nature.
441:475–482. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo W, Lasky JL, Chang CJ, et al:
Multi-genetic events collaboratively contribute to Pten-null
leukaemia stem-cell formation. Nature. 453:529–533. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Breccia M and Alimena G: NF-κB as a
potential therapeutic target in myelodysplastic syndromes and acute
myeloid leukemia. Expert Opin Ther Targets. 14:1157–1176. 2010.
|
|
25
|
Cilloni D, Martinelli G, Messa F,
Baccarani M and Saglio G: Nuclear factor κB as a target for new
drug development in myeloid malignancies. Haematologica.
92:1224–1229. 2007.
|
|
26
|
Liu KS, Zhang Y, Ding WC, et al: The
selective Hsp90 inhibitor BJ-B11 exhibits potent antitumor activity
via induction of cell cycle arrest, apoptosis and autophagy in
Eca-109 human esophageal squamous carcinoma cells. Int J Oncol.
41:2276–2284. 2012.PubMed/NCBI
|
|
27
|
Calderwood SK, Khaleque MA, Sawyer DB and
Ciocca DR: Heat shock proteins in cancer: chaperones of
tumorigenesis. Trends Biochem Sci. 31:164–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mjahed H, Girodon F, Fontenay M and
Garrido C: Heat shock proteins in hematopoietic malignancies. Exp
Cell Res. 318:1946–1958. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jego G, Hazoumé A, Seigneuric R and
Garrido C: Targeting heat shock proteins in cancer. Cancer Lett.
332:275–285. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Neckers L and Workman P: Hsp90 molecular
chaperone inhibitors: are we there yet? Clin Cancer Res. 18:64–76.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chiosis G, Lucas B, Huezo H, Solit D,
Basso A and Rosen N: Development of purine-scaffold small molecule
inhibitors of Hsp90. Curr Cancer Drug Targets. 3:371–376. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Braun T, Carvalho G, Fabre C, Grosjean J,
Fenaux P and Kroemer G: Targeting NF-kappaB in hematologic
malignacies. Cell Death Differ. 13:748–758. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kerbauy DM, Lesnikov V, Abbasi N, Seal S,
Scott B and Deeq HJ: NF-kappaB and FLIP in arsenic trioxide
(ATO)-induced apoptosis in myelodysplastic syndromes (MDSs). Blood.
106:3917–3925. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chapuis N, Tamburini J, Cornillet-Lefebvre
P, et al: Autocrine IGF-1/IGF-1R signaling is responsible for
constitutive PI3K/Akt activation in acute myeloid leukemia:
therapeutic value of neutralizing anti-IGF-1R antibody.
Haematologica. 95:415–423. 2010. View Article : Google Scholar
|
|
35
|
He Q, Li X, Zhang Z, et al: Overexpression
of IGF-IR in malignant clonal cells in bone marrow of
myelodysplastic syndromes. Cancer Invest. 28:983–988. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mendoza MC, Er EE and Blenis J: The
Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation. Trends
Biochem Sci. 36:320–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Economopoulou C, Pappa V, Papageorgiou S,
et al: Cell cycle and apoptosis regulatory gene expression in the
bone marrow of patients with de novo myelodysplastic syndromes
(MDS). Ann Hematol. 89:349–358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Quesnel B, Guillerm G, Vereecque R, et al:
Methylation of the p15(INK4b) gene in myelodysplastic syndromes is
frequent and acquired during disease progression. Blood.
91:2985–2990. 1998.PubMed/NCBI
|
|
39
|
Chen G, Zeng W, Miyazato A, et al:
Distinctive gene expression profiles of CD34 cells from patients
with myelodysplastic syndrome characterized by specific chromosomal
abnormalities. Blood. 104:4210–4218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Olnes MJ, Shenoy A, Weinstein B, et al:
Directed therapy for patients with myelodysplastic syndromes (MDS)
by suppression of cyclin D1 with ON 01910. Na Leuk Res. 36:982–989.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Böll B, Eltaib F, Reiners KS, et al: Heat
shock protein 90 inhibitor BIIB021 (CNF2024) depletes NF-kappaB and
sensitizes Hodgkin’s lymphoma cells for natural killer
cell-mediated cytotoxicity. Clin Cancer Res. 15:5108–5116.
2009.PubMed/NCBI
|