Introduction
Hypoxia-induced pulmonary hypertension (HPH) is a
complication of numerous pulmonary conditions, which is associated
with hypoxia. HPH is usually a life-threatening condition in
neonatal populations. In conditions of severe hypoxemia (1), pulmonary vascular contraction and
persistent cramp leads to persistent pulmonary hypertension with a
poor prognosis. Unrelieved pulmonary hypertension can lead to
right-sided heart failure and even mortality. However, the
pathogenesis of neonatal HPH is not completely clear.
A number of adult studies (2–4)
investigating the pathogenesis of HPH have been conducted in adult
populations and animals. Hypoxia may lead to the injury of
endothelial cells in the pulmonary arterioles, triggering the onset
of pulmonary hypertension. Injury-induced dysfunction of the
endothelium leads to the abnormal secretion of vasoactive
substances and cytokines, thus, causes a functional imbalance
between vasoconstrictors and vasodilators. This functional
imbalance exerts effects on vascular smooth muscle cells, resulting
in pulmonary vasoconstriction in the early stages, which is
followed by pathological changes in the pulmonary vascular wall,
vascular remodeling and pulmonary hypertension during the late
stages. As a key nuclear transcription factor in hypoxic
conditions, hypoxia-inducible factor (HIF)-1α is considered to be
closely associated with the pathogenesis of HPH, functioning as the
central link (5) in mediating
pulmonary vasoconstriction and the remodeling of pulmonary vascular
and hypoxia-induced gene expression. Endothelin (ET)-1 functions as
an endogenous vasoconstrictor; with increasing mean pulmonary
artery pressure (mPAP), the level of ET-1 (6–7) in
the serum and lung tissue also increases. Adrenomedullin (ADM)
functions as a vasodilator, thus, with increased levels of ADM
(8), pulmonary vasodilation is
gradually enhanced and the mPAP is reduced. ET-1 and ADM are key
target genes of HIF-1α (9), which
are involved in vasoconstriction, vascular remodeling and the
development of HPH. Study into the pathogenesis of HPH is limited
to adult and associated animal experiments, thus, research into
neonatal HPH requires further investigation. It remains uncertain
whether HIF-1α plays a similar role in the pathogenesis of HPH in
newborns as it does in adults. The effects of HIF-1α, ET-1 and ADM
in newborns with HPH are also unknown. Thus, the aim of the present
study was to establish a model of HPH in newborn rats in order to
evaluate the changes in the expression levels of HIF-1α, ET-1 and
ADM and the effect of HIF-1α and its regulatory factors, ET-1 and
ADM, in the blood serum and lung tissue. Providing information on
the pathogenesis of HPH in newborns may aid the identification of
novel treatments for HPH in neonates.
Materials and methods
Animal model
All the procedures involving animals were approved
by the Animal Care Council and Animal Ethics Committee of the First
Affiliated Hospital of Xinjiang Medical University (Ürümqi,
China).
Healthy newborn Wistar rats (weight, 15–25 g; age,
3–5 days) were used in the study. In total, data were collected
from 96 rats. Each animal was randomly assigned to one of the six
subgroups in the hypoxic or control groups (n=8 per subgroup). In
the hypoxic group, the subgroups were based on the duration of
hypoxia (3, 5, 7, 10, 14 or 21 days), while the subgroups of the
control group were subjected to normoxia for 3, 5, 7, 10, 14 or 21
days.
Rats in the hypoxic group were treated in order to
create a model of HPH (10). All
the rats in the hypoxic group were placed in a normobaric hypoxic
chamber and a gas mixture of 8% O2 mixed with
N2 was pumped into the chamber at a rate of 1.5 l/min.
The fraction of inspired oxygen (FiO2) in the chamber
was monitored using an oxygen analyzer (CY-100B; Lihua Science
& Technology Co., Ltd., Hangzhou, China) and maintained in the
range of 10±0.5%. Hypoxic conditions were sustained for 8 h per day
(day/night ratio, 12/12 h). For each subgroup in the hypoxic group,
the end point was at day 3, 5, 7, 10, 14 or 21 following exposure
to hypoxia. The rats in the control group were maintained under
similar conditions, but without being subjected to hypoxia.
mPAP monitoring
At the predesignated time points (day 3, 5, 7, 10,
14 or 21), the rats from the appropriate control and hypoxic
subgroups were anesthetized, placed on their backs, the trachea was
exposed and punctured with a venous trocar and the trocar was then
pushed into the trachea. An animal respirator (HX-200 small animal
ventilator, Taimeng Technology Co., Ltd., Chengdu, China) was
connected to provide mechanical ventilation with the following
respirator parameters: Respiratory rate, 100–120 breaths/min and
tidal volume, 2–3 ml/min). A longitudinal skin incision was made on
the left sternal border, and blunt layer-by-layer separation of the
tissues was performed until the heart was fully exposed. The tip of
a scalp needle was punched through the base of the pulmonary artery
and then orientated against the direction of the blood flow. The
other end of the scalp needle was connected to a pressure sensor
(BP-6; Thaimeng Technology Co., Chengdu, China) that measured the
mPAP (11).
Measurement of HIF-1α, ET-1 and ADM mRNA
expression levels in the lung tissue
Lung tissue samples were collected from the rats
within 5 min of sacrifice and stored in liquid nitrogen until
required for analysis. Total RNA was extracted from 60–100-mg
samples of the left upper lobe lung tissue. Quantitative polymerase
chain reaction (PCR) was performed with each group. The RNA
concentration and purity were determined using an ultraviolet
spectrophotometer (SmartSpec Plus; Bio-Rad, Hercules, CA, USA), and
cDNA was obtained using a reverse-transcription kit, according to
the manufacturer’s instructions. The samples were stored at −20°C.
The primers used for quantitative PCR analysis are shown in
Table I and the β-actin gene was
used as an internal reference to normalize the mRNA expression
levels of HIF-1α, ET-1 and ADM. The conditions for quantitative PCR
were as follows: 95°C for 30 sec; 40 cycles of 95°C for 10 sec,
53°C for 30 sec and 65°C for 10 sec; followed by a melting curve
program.
 | Table IPCR primers used in the study. |
Table I
PCR primers used in the study.
| Gene | Primers | Amplified fragment
length (bp) |
|---|
| β-actin | F:
GGAGATTACTGCCCTGGCTCCTA
R: GACTCATCGTACTCCTGCTTGCTG | 150 |
| HIF-1α | F:
CAACTGCCACCACTGATGAAT
R: CCACTGTATGCTGATGCCTTAG | 133 |
| ET-1 | F:
TGTTCAGACTGGCAGAGGAC
R: CAAGAAGAGGCAAGAGAATCACT | 120 |
| ADM | F:
GAGCGGACTGAGACAATC
R: GTAAGTAATGAGGCGTATGC | 192 |
Measurement of vascular remodeling
Tissue samples were collected at sacrifice from the
upper lobe of the right lung. The samples were fixed in 10% neutral
formalin for one week, subjected to conventional paraffin embedding
and 4-μm thick sections were prepared and stained with hematoxylin
and eosin. Three sections were randomly selected from each rat,
from which five pulmonary arterioles, next to the respiratory
bronchioles and alveoli with relatively round cross sections and a
diameter of 50–100 μm, were selected for analysis. The intima-media
thickness/external diameter ratio (MT%) and medial wall
cross-sectional area/vessel total cross-sectional area ratio (MA%)
of the selected arterioles were analyzed using pathological image
analysis software (PIPS-2020; Chongqing Tianhai Medical Equipment
Co., Ltd., Chongqing, China) to evaluate the thickening of the
pulmonary arteriole walls.
Statistical analysis
Results are expressed as the mean ± standard error.
One-way analysis of variance was used to evaluate the differences
between two groups, where P<0.05 was considered to indicate a
statistically significant difference. The analysis was performed
using the SPSS version 18.0 software (SPSS, Inc., Chicago, IL,
USA).
Results
Hypoxia increases the mPAP
Significant increases in the mPAP were observed in
each subgroup of the hypoxic group when compared with the
respective control subgroups of neonatal rats (P<0.05). The mPAP
elevated as the duration of hypoxia increased in the hypoxic group
(Table II).
| Table IIAnalysis of the mPAP in the various
hypoxic and control subgroups (n=8 per subgroup). |
Table II
Analysis of the mPAP in the various
hypoxic and control subgroups (n=8 per subgroup).
| Subgroup | mPAP (mmHg) |
|---|
| Day 3 hypoxia | 8.59±1.57b |
| Day 3 control | 6.14±1.02 |
| Day 5 hypoxia | 10.02±1.81a |
| Day 5 control | 8.24±1.06 |
| Day 7 hypoxia | 11.63±2.56b |
| Day 7 control | 8.33±0.76 |
| Day 10 hypoxia | 14.84±2.06b |
| Day 10 control | 10.16±2.15 |
| Day 14 hypoxia | 15.29±2.88b |
| Day 14 control | 10.92±2.74 |
| Day 21 hypoxia | 18.04±2.69b |
| Day 21 control | 12.17±1.64 |
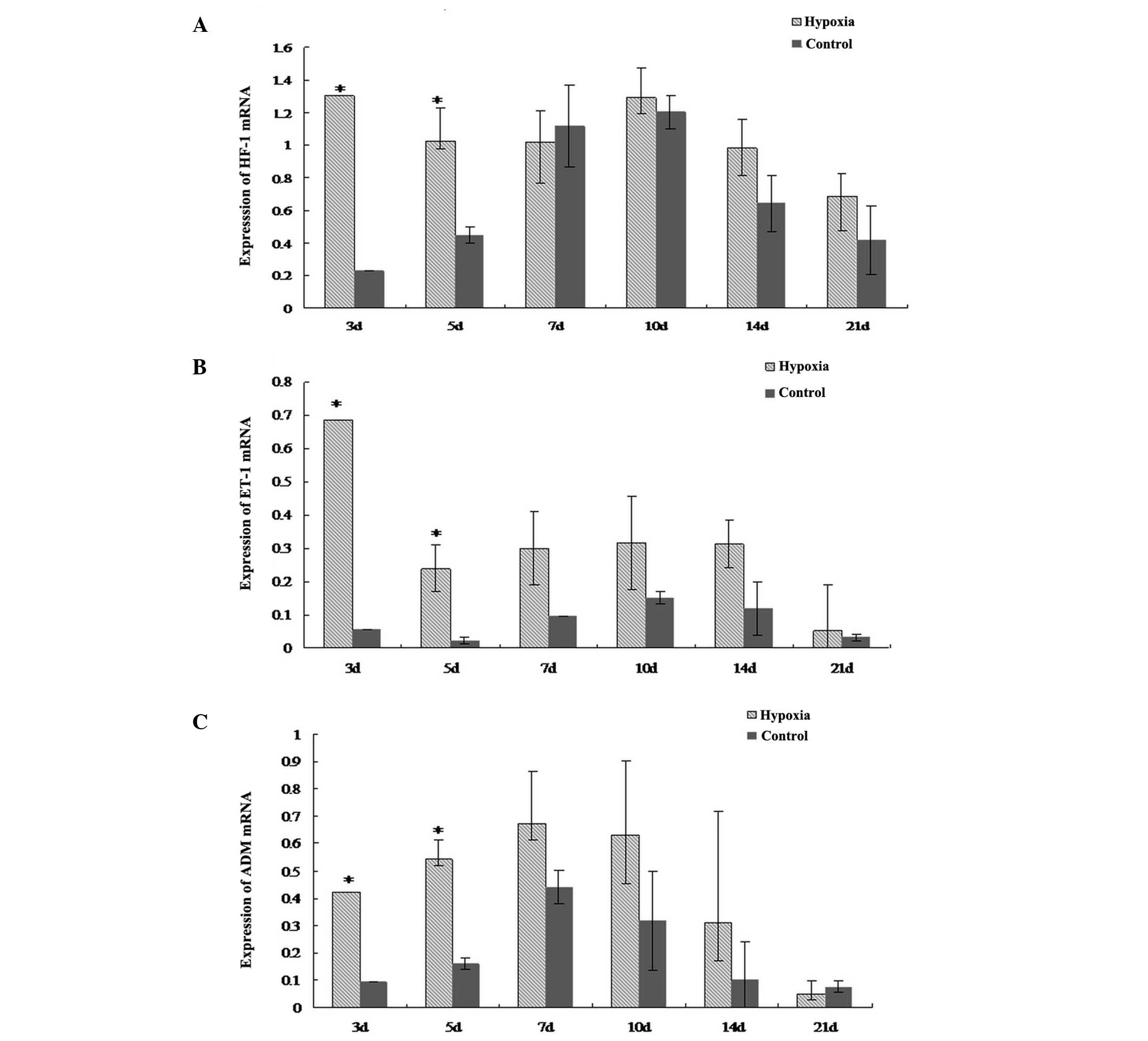
Hypoxia increases the mRNA expression
levels of HIF-1α, ET-1 and ADM in the lungs
The mRNA expression levels of HIF-1α, ET-1 and ADM
in the lungs of the neonatal rats in the hypoxic group were higher
at day three and five when compared with the respective control
subgroups (P<0.05; Fig. 1).
However, at the later time points, the mRNA expression levels of
HIF-1α, ET-1 and ADM were not significantly different between the
hypoxic and control groups.
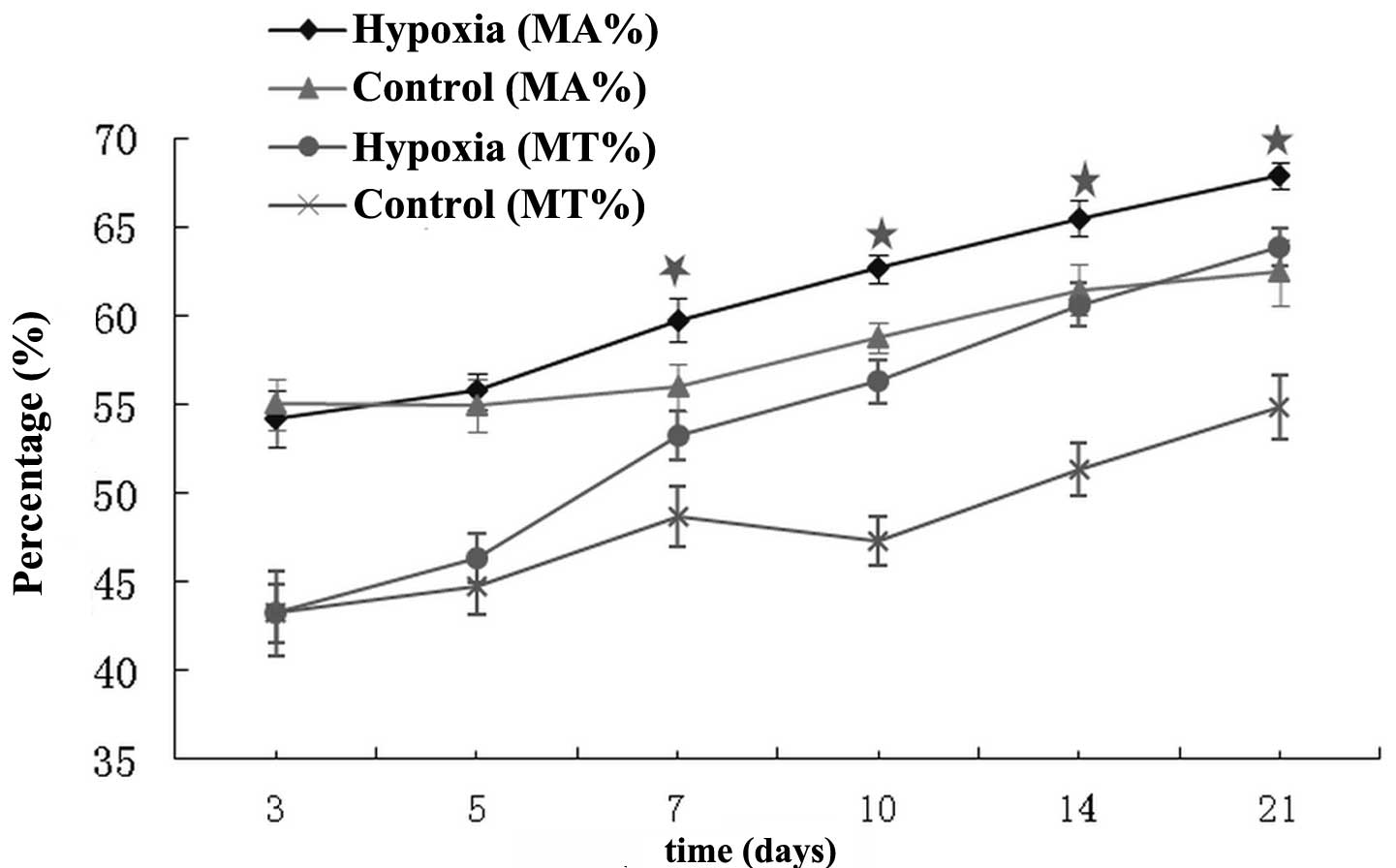
Pulmonary vascular remodeling
MT% and MA% ratios of the pulmonary arterioles
increased in the hypoxic group from day seven onwards when compared
with the respective control subgroups (P<0.05; Fig. 2).
Discussion
Research into the pathogenesis of HPH has revealed
that HIF-1α plays an important role in pulmonary vascular
constriction, vascular remodeling (12–14)
and the development of pulmonary hypertension, possibly by
regulating the expression levels of ET-1 and ADM at a
transcriptional level. HIF-1α expression is very sensitive to the
intracellular oxygen concentration. Under normoxic conditions,
HIF-1α is continuously synthesized and expressed in the cytoplasm,
but is rapidly degraded by the ubiquitin-mediated pathway. However,
under hypoxic conditions, HIF-1α expression exponentially increases
and HIF-1α accumulates in the nucleus due to the inhibition of
hydroxylation and protease degradation (15). The present study demonstrated that
the mRNA expression levels of HIF-1α increased during the early
stages of hypoxia, however, after seven days of hypoxia, HIF-1α
mRNA was expressed at a similar level to the control group. This
observation may be due to the development of tolerance to hypoxia.
Under moderate hypoxic conditions, HIF-1α expression is
predominantly regulated by changes in the oxygen concentration
(2). In the present study,
neonatal rats were continuously exposed to low concentrations of
oxygen at a FiO2 of 10±0.5%, with no changes in the
oxygen concentration. Therefore, at the late stages of hypoxia (day
seven onwards), HIF-1α mRNA expression was not regulated by hypoxia
and was similar to that of the control group. The results of the
present study indicate that hypoxia induces high levels of HIF-1α
mRNA expression in the lung tissues during the early stages of HPH
development. However, the modulatory effects of hypoxia on HIF-1α
expression become weaker during the late stages of hypoxia, with
the HIF-1α expression levels reverting to normal levels, instead of
increasing with the mPAP.
ET-1, a vasoconstrictive substance, is secreted by
endothelial cells in response to injury or excessive activation
(16). ET-1 exerts marked effects
on vasoconstriction and vascular remodeling. The lung is one of the
most important target organs for ET-1 metabolism (17). During the early stages of hypoxia,
arterial endothelial cells are stimulated to secrete large amounts
of HIF-1α, which further induces high expression levels of the
HIF-1α target gene, ET-1. However, as there is a lack of dense
granules in pulmonary endothelial cells for the storage of ET-1
mRNA, and ET-1 mRNA is unstable, ET-1 activity gradually decreases
over time during hypoxia. In the present study, the mRNA expression
of ET-1 increased during the first five days of hypoxia; however,
no marked changes were observed after five days. During the late
stages of hypoxia, severe damage to the endothelial cells leads to
continually low levels of ET-1 secretion, and the degradation of
ET-1 in the lung tissues also decreases. Therefore, ET-1 mRNA
expression was not significantly different in the lung tissues of
the hypoxic and control groups during the late stages of hypoxia.
The results indicate that ET-1 may play an important role in the
pathogenesis of HPH in newborn rats.
Imbalances between vasoconstrictors and vasodilators
play a key role in the development of HPH. There are two
HIF-1α-binding sites in the ADM promoter, and HIF-1α can regulate
the transcription of ADM upon binding to the ADM promoter. Mutation
of the HIF-1α-binding sites in the ADM promoter decreases the
expression of ADM; therefore, HIF-1α regulates ADM at a
transcriptional level. Numerous studies on adult rats have
hypothesized that hypoxia can induce the synthesis of ADM (18, 19). In adult rats, ADM mRNA expression
increases continuously in response to hypoxia, with the most
pronounced increases observed in arterial endothelial cells and
vascular smooth muscle cells. Increased ADM expression may be
involved in lowering vascular tension (20) and alleviating hypoxia-induced
damage to the lung tissues by inducing vasodilation. In the present
study, HIF-1α and ADM mRNA expression levels significantly
increased during the early stages of hypoxia when compared with the
control group. This observation indicates that the accumulation of
HIF-1α may upregulate ADM mRNA expression, which may enhance the
synthesis and secretion of ADM in endothelial and smooth muscle
cells. In response to prolonged hypoxia, vascular remodeling and
endothelial injury occur and the activity of the
endothelium-derived vasodilator pathway decreases, resulting in a
gradual decrease in ADM synthesis and suppression of the effects of
ADM on vascular smooth muscle cell proliferation (21). Finally, the potent vasoconstriction
and cell proliferation promoted by ET-1 lead to an imbalance
between vasoconstrictors and vasodilators, resulting in the
development of vascular remodeling and pulmonary hypertension
(22). These observations indicate
that the vasoconstrictor, ET-1, dominates during the development of
HPH in neonatal rats, whereas the vasodilator, ADM, exerts weaker
effects.
In the present study, no marked pathological changes
were observed in the pulmonary arterioles during the early stages
of hypoxia. However, vascular remodeling occurred in the pulmonary
arterioles after seven days of hypoxia. Therefore, the results
indicate that seven days of hypoxia is the cut-off point at which
the pulmonary arterioles undergo transition from functional change
to anatomical change in pulmonary hypertension. Thus, seven days
represents the appropriate period of time for early management to
prevent pulmonary vascular remodeling during the progression of
HPH. Further study investigating pulmonary vascular remodeling in
newborns is required in order to provide accurate clinical
guidelines for the treatment of HPH in neonates.
In conclusion, HIF-1α is an important factor in the
development of HPH in newborn rats, and ET-1 and ADM are also
involved in this process. The present study has confirmed that the
mRNA expression levels of HIF-1α, ET-1 and ADM are elevated in the
lung tissues during the early stages of hypoxia in newborn rats.
However, during prolonged hypoxia, although the mPAP is elevated
and vascular remodeling occurs in the pulmonary arterioles, the
mRNA expression levels of HIF-1α, ET-1 and ADM are not
significantly different when compared with control animals.
Acknowledgements
The study was supported by a grant from the National
Natural Science Foundation of China (no. 30960410).
References
|
1
|
Greenough A and Khetriwal B: Pulmonary
hypertension in the newborn. Peadiatr Respir Rev. 6:111–116. 2005.
View Article : Google Scholar
|
|
2
|
Preston IR: Clinical perspective of
hypoxia-mediated pulmonary hypertension. Antioxid Redox Signal.
9:711–721. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bonnet S, Michelakis ED, Porter CJ, et al:
An abnormal mitochondrial-hypoxia inducible factor-1a-Kv channel
pathway disrupts oxygen sensing and triggers pulmonary arterial
hypertension in fawn hooded rats: similarities to human pulmonary
arterial hypertension. Circulation. 113:2630–2641. 2006. View Article : Google Scholar
|
|
4
|
Wei HL, Zhang CY, Jin HF, Tang CS and Du
JB: Hydrogen sulfide regulates lung tissue-oxidized glutathione and
total antioxidant capacity in hypoxic pulmonary hypertensive rats.
Acta Pharmacol Sin. 29:670–679. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marzo F, Lavorgna A, Coluzzi G, et al:
Erythropoietin in heart and vessels: focus on transcription and
signaling pathways. J Thromb Thrombolysis. 26:183–187. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Higenbottam T: Pulmonary hypertension and
chronic obstructive pulmonary disease: a case for treatment. Proc
Am Thorac Soc. 2:12–19. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu J, Discher DJ, Bishopric NH and Webster
KA: Hypoxia regulates expression of the endothelin-1 gene through a
proximal hypoxia-inducible binding site on the antisense strand.
Biochem Biophys Res Commun. 245:894–899. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nagaya N, Nishikimi T, Uematsu M, et al:
Haemodynamic and hormonal effects of adrenomedullin in patients
with pulmonary hypertension. Heart. 84:653–658. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nishikimi T, Nagata S, Sasaki T, et al:
The active molecular form of plasma adrenomedullin is extracted in
the pulmonary circulation in patients with mitral stenosis:
possible role of adrenomedullin in pulmonary hypertension. Clin Sci
(Lond). 100:61–66. 2001. View Article : Google Scholar
|
|
10
|
Prass K, Scharff A, Ruscher K, et al:
Hypoxia-induced stroke tolerance in the mouse is mediated by
erythropoietin. Stroke. 34:1981–1986. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qing X and Keith IM: Targeted blocking of
gene expression for CGRP receptors elevates pulmonary artery
pressure in hypoxic rats. Am J Physiol Lung Cell Mol Physiol.
285:L86–L96. 2003.PubMed/NCBI
|
|
12
|
Fu H, Luo F, Yang L, Wu W and Liu X:
Hypoxia stimulates the expression of macrophage migration
inhibitory factor in human vascular smooth muscle cells via
HIF-1alpha dependent pathway. BMC Cell Biol. 11:662010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wiesner D, Merdian I, Lewerenz J, Ludolph
AC, Dupuis L and Witting A: Fumaric acid esters stimulate
astrocytic VEGF expression through HIF-1a and Nrf2. PLoS One.
8:e766702013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He ZH, Dai AG, Zhang XF and Tan XW:
Expressions of hypoxia inducible factor-1 alpha and inducible
nitric oxide synthase gene in the development of hypoxic pulmonary
hypertension in rats. Clin Rehab Tissue Eng Res. 11:7290–7294.
2007.
|
|
15
|
Smith TG, Robbins PA and Ratcliffe PJ: The
human side of hypoxia-inducible factor. Br J Haematol. 141:325–334.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shreenivas S and Oparil S: The role of
endothelin-1 in human hypertension. Clin Hemorheol Microcirc.
37:157–178. 2007.
|
|
17
|
Schindler MB, Hislop AA and Haworth SG:
Porcine pulmonary artery and bronchial responses to endothelin-1
and norepinephrine on recovery from hypoxic pulmonary hypertension.
Pediatr Res. 60:71–76. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maybin JA, Battersby S, Hirani N,
Nikitenko LL, Critchley HO and Jabbour HN: The expression and
regulation of adrenomedullin in the human endometrium: a candidate
for endometrial repair. Endocrinology. 152:2845–2856. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shimoda LA and Semenza GL: HIF and the
lung: role of hypoxia-inducible factors in pulmonary development
and disease. Am J Respir Crit Care Med. 183:152–156. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Westphal M, Booke M and Dinh-Xuan AT:
Adrenomedullin: a smart road from pheochromocytoma to treatment of
pulmonary hypertension. Eur Respir J. 24:518–520. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen L, Qiu JH, Zhang LL and Luo XD:
Adrenomedullin promotes human endothelial cell proliferation via
HIF-1α. Mol Cell Biochem. 365:263–273. 2012.PubMed/NCBI
|
|
22
|
Oladipupo S, Hu S, Kovalski J, et al: VEGF
is essential for hypoxia-inducible factor-mediated
neovascularization but dispensable for endothelial sprouting. Proc
Natl Acad Sci USA. 108:13264–13269. 2011. View Article : Google Scholar : PubMed/NCBI
|