Introduction
Deep hypothermic circulatory arrest (DHCA) has
become a common technique used to protect vital organs during
surgical interventions on the thoracic aorta in adults, neonates
and infants undergoing surgery for complex congenital heart disease
(1,2). Despite the excellent surgical results
that may be obtained with DHCA, neurological dysfunction, renal
failure and other organ damage remain common complications
following DHCA (1,3). Bleeding also remains a constant risk
following DHCA (3). An
understanding of the complex interactions between various cells,
such as leukocytes, platelets and endothelial cells, and the
coagulation, fibrinolytic and inflammatory pathways, in addition to
the impact of hypothermia on these pathways is essential for
further improvement of DHCA.
One parameter that is known to have a significant
impact on inflammatory cell activation is temperature (4). Hypothermia significantly decreases
the global cerebral metabolic rate of glucose and oxygen. The
metabolic rate of the human body at 18°C is only 12–25% of the
metabolic rate at normal temperature (2). Experimental data has demonstrated
that moderate hypothermia delays the production of proinflammatory
cytokines and nuclear factor-κB (NF-κB) activation. NF-κB plays a
pivotal role in regulating the transcription of cytokines, cell
adhesion molecules and other mediators involved in the inflammatory
response (5). Hypothermia may
affect many metabolic pathways, reactions of inflammation and
apoptosis processes, among many other pathways. A possible
explanation for the success of therapeutic hypothermia is the
multiple active mechanisms blocking the cascade of ischemia at a
number of levels (6). In addition,
hypothermia also affects hemostasis at various levels. It alters
platelet function, the enzymatic kinetics in the coagulation
cascade and the dynamic equilibrium of the fibrinolytic system
(5).
Activated leukocytes are key mediators of
inflammatory reactions due to their ability to release
tissue-damaging compounds following their adherence to endothelial
cells (7). Inflammatory cells,
such as monocytes, are recruited in response to injury cues. They
express a panoply of proinflammatory genes through a combination of
transcription factors of which NF-κB is the most fundamental
(8). The firm adhesion of
leukocytes to the endothelium and their subsequent transmigration
through the endothelium junctions represents the early stages of
the inflammatory response following conditions such as ischemic
injury, atherosclerosis and various other inflammatory disorders
(9). Monocytes play an important
role in regulating the thrombotic and fibrinolytic systems, and
cell adhesion molecules trigger cellular interactions at the
interface of thrombosis and fibrinolysis (10). Additionally, leukocyte-platelet
interactions play an important role in inflammatory reactions. Both
platelets and leukocytes are able to modulate each other’s
functions. Leukocytes enhance platelet-mediated aggregation via
interaction of their P-selectin ligand with P-selectin; binding of
leukocytes to platelets promotes leukocyte activation (1,11).
The cell-cell interaction between leukocytes, endothelial cells and
platelets is mediated by adhesion molecules on the surface of these
cells. Thus, modulation of the expression of cell adhesion
molecules on leukocytes during blood stasis and hypothermia may
have an impact on inflammatory responses during DHCA.
Studies concerning hypothermic conditions have
mainly focused on platelet activation, platelet-leukocyte
interactions or inflammatory markers; however, to the best of our
knowledge, no data are available concerning the expression of cell
adhesion molecules in leukocytes during DHCA. The purpose of the
present study was to investigate the expression of the cell
adhesion molecules cluster of differentiation (CD) molecule 11B
[CD11b; also known as α subunit of the β2-integrin macrophage-1
antigen (MAC-1)], CD54 [also known as intercellular adhesion
molecule 1 (ICAM-1)] and CD162 [also known as P-selectin
glycoprotein ligand 1 (PSGL-1)] on monocytes under normothermic
(36°C) and hypothermic (18°C) conditions using an in vitro
experimental model that simulates the conditions of circulatory
arrest.
Materials and methods
Subjects
Healthy volunteers were included in the present
study following the provision of informed consent. The study
protocol conforms to the ethical guidelines of the 1975 Declaration
of Helsinki and was approved by the local Ethics Committee
(University of Tuebingen, Tuebingen, Germany).
Blood sampling and sample
preparation
Blood from non-medicated healthy male volunteers
(n=11) was collected by venipuncture with a 21-gauge needle from an
antecubital vein. The first 5 ml of blood was discharged prior to
the drawing of additional blood samples for analysis. All blood
samples were anti-coagulated with 3 U/ml heparin.
One blood sample set from each subject was incubated
in stasis for 30 min at 18°C to simulate DHCA. The other blood
sample set was incubated in stasis for 30 min at 36°C. Each of the
sample sets (18°C and 36°C) consisted of three subsets: i) one
subset was incubated with an antibody combination of anti-CD45 and
anti-CD11b; ii) the second subset was incubated with anti-CD45 and
anti-CD54; and iii) the third subset was incubated with anti-CD45
and anti-CD162, as described below. For incubation of the blood
samples, a specially designed temperature-regulation device with a
heating and cooling function was used (11). The model was intended to mimic the
conditions of stasis and hypothermia.
Sub-samples preparation for
flow-cytometry
The following incubation steps were performed
immediately after the 30 min period of incubation in stasis using a
previously described method (12,13).
Expression of the following cell adhesion molecules
on monocytes was measured by flow cytometry (Coulter Epics XL-MCL™;
Beckman Coulter, Krefeld, Germany). Leukocytes were detected using
fluorescein isothiocyanate (FITC)-conjugated anti-lymphocyte common
antigen (anti-CD45; BD Biosciences, Heidelberg, Germany). The
following phycoerythrin (PE)-conjugated monoclonal antibodies were
used for cell detection in fluorescence-activated cell sorting
(FACS): Anti-CD11b, anti-CD54 (both from BD Biosciences) and
anti-CD162 (Beckman Coulter). Whole blood (100 μl) was incubated
with saturating concentrations of FITC-conjugated anti-CD45 and
PE-conjugated monoclonal antibodies for 20 min at room temperature.
Erythrocytes were lysed and leukocytes were fixed with a
commercially available solution (FACS Lysing Solution, BD
Biosciences). Samples were then incubated for 10 min in the dark.
Thereafter, samples were centrifuged at 200 × g for 10 min, the
pellet washed with phosphate-buffered saline (Gibco, Invitrogen
Life Technologies, Karlsruhe, Germany) and centrifuged again. The
pellet was then resuspended in phosphate-buffered saline and
applied to the flow cytometer equipped with a 488 nm argon laser.
Results are expressed as mean fluorescence intensity (MFI) of
CD11b, CD54 and CD162 on monocytes.
Statistical methods
The Mann-Whitney U test for independent samples was
used to analyze the significance of any differences between samples
incubated at 18°C and samples incubated at 36°C. Data are presented
as median and interquartile range (IQR, 25th to 75th percentiles).
A two-tailed P value <0.05 was considered to indicate a
statistically significant difference. The calculations were
performed using InStat (GraphPad Software, Inc., San Diego, CA,
USA) and IBM SPSS software (IBM, Ehningen, Germany).
Results
The volunteers (n=11) had no history of acute or
chronic disease and were non-medicated. Their mean age was 28.7
years. All donors were male (ethnic background: all Caucasian).
Following an incubation period of 30 min in a
circulatory arrest model simulating blood stasis with different
temperatures, a significant association between temperature and
surface expression of CD11b and CD162 on monocytes was
demonstrated.
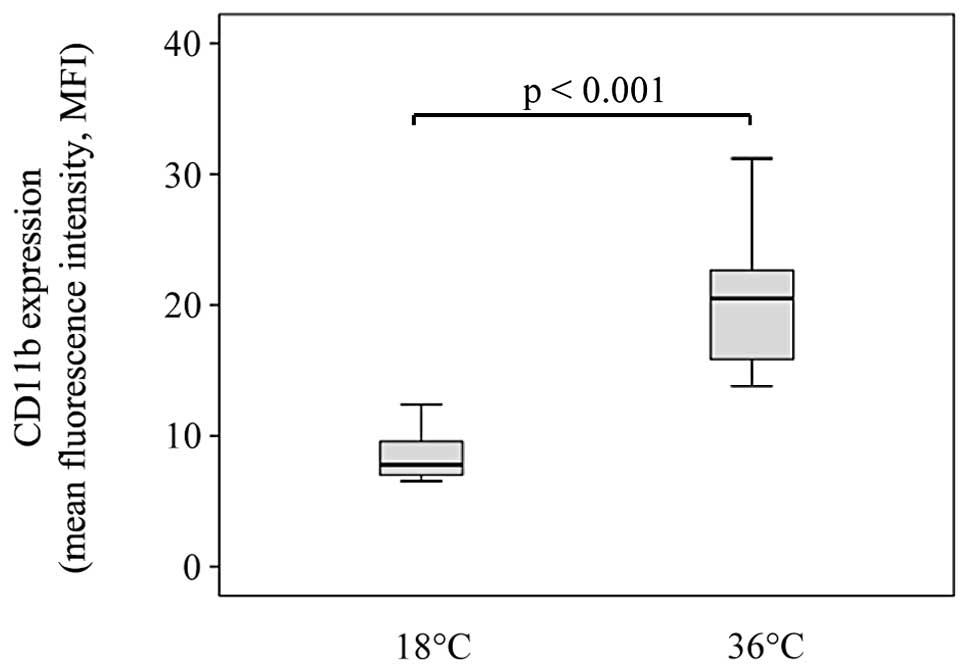
The expression of CD11b on monocytes was
significantly decreased following the incubation of blood at 18°C
compared with the expression of CD11b on monocytes after incubation
at 36°C (median MFI 7.78, IQR 7.0–9.6 vs. median MFI 20.5, IQR
15.4–22.7, respectively, P<0.001). After 30 min of blood stasis
in the circulatory arrest model, the expression of CD162 on
monocytes was significantly lower at 18°C compared with that at
36°C (median MFI 16.4, IQR 15.9–19.1 vs. median MFI 29.8, IQR
27.9–30.7, respectively, P<0.001). No association was identified
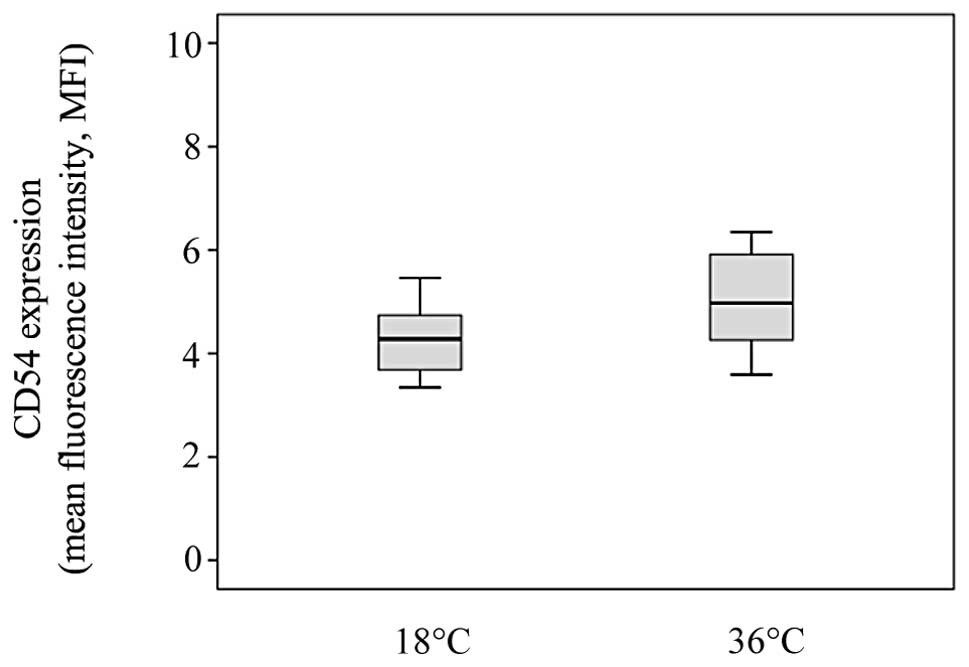
between temperature and the surface expression of CD54 on monocytes
after 30 min of stasis. The expression of CD54 on monocytes did not
differ between 18°C and 36°C (median MFI 4.28, IQR 3.7–4.7 vs.
median MFI 4.97, IQR 4.3–6.0, respectively, P>0.05). Medians
with quartiles for MFI are depicted in Figs. 1–3.
Discussion
During certain cardiac surgical procedures, such as
those on the thoracic aorta or during the repair of congenital
cardiac defects, extracorporeal circulation (ECC) in combination
with deep hypothermia (<20°C) may be performed, particularly
during deep hypothermic circulatory arrest (DHCA) (14). DHCA has been demonstrated to be an
effective and safe organ protection technique used during periods
of interrupted blood flow (2,15).
The physiological basis for the preservation of tissue integrity
during a period of interrupted circulation through hypothermia
centers on the lowering of the metabolic rate through a reduction
in temperature (15). Another
reason is a reduced systemic inflammatory response with DHCA since
temperature has a significant impact on inflammatory cell
activation (4,16). Activated leukocytes are key
mediators of inflammatory reactions due to their ability to release
tissue-damaging compounds following their adherence to endothelial
cells (7). Hypothermia is known to
have anti-inflammatory effects by inhibiting leukocyte response
following several types tissue insults such as ischemic brain or
liver injury (4). The interaction
between leukocytes, endothelial cells and platelets is mediated by
cell adhesion molecules that are expressed on the surface of
activated cells. In a prior investigation, a temperature-dependent
regulation of the surface expression of CD11b and CD162 on
monocytes in an ECC model was demonstrated (13). However, to the best of our
knowledge, no data are available to date regarding the expression
of cell adhesion molecules on leukocytes during DHCA.
In the present in vitro study, the effects of
blood temperature on the surface expression of cell adhesion
molecules on monocytes were investigated using an experimental
model simulating circulatory arrest. A difference in the expression
levels of CD11b and CD162 on monocytes was observed between blood
incubated at 18°C for 30 min compared with blood incubated at 36°C
for the same time period.
CD11b/CD18 (MAC-1) is a member of the β2-integrin
family. It is present in an inactive state on circulating
leukocytes; however, upon neutrophil stimulation with various
cytokines it undergoes a rapid conformational change that results
in its activation, which is required for optimal integrin function.
β2-integrins mediate leukocyte adhesion and transmigration across
the endothelium, through interactions with ICAM-1 on the activated
endothelium. Several adhesion-dependent neutrophil functions, such
as binding to fibrinogen, immune complexes and platelets, are
MAC-1-dependent owing to the large variety of ligands for MAC-1 and
its ability to cooperate functionally with a variety of other
surface receptors (17). The
P-selectin/MAC-1 cascade gives rise to heterotypic conjugates of
platelets with leukocytes. Leukocyte tethering by the P-selectin of
platelets not only induces rapid β2-integrin activation but also
triggers delayed responses through the induction of the expression
of transcription factors such as NF-κB. This activates gene
transcription for the synthesis of proinflammatory molecules; these
are fundamental for leukocytes to acquire an inflammatory phenotype
(18). Experimental data
demonstrate that hypothermia delays the production of
proinflammatory cytokines and NF-κB activation (5). Furthermore, studies on platelet
function have reported a DHCA-induced platelet dysfunction, and
hypothermia has been revealed to induce platelet-aggregate
formation (1,3). Additionally, soluble fibrinogen
promotes neutrophil activation in a MAC-1-dependent manner and the
interaction of platelets with leukocytes may result in fibrin
deposition through increased tissue-factor expression (17,18).
Tissue-type plasminogen activator (tPA) promotes the aggregation
and interaction of annexin A2 and MAC-1, leading to the clustering
and activation of MAC-1 signaling in macrophages (8). Thus, CD11b (MAC-1) is a surface
receptor on stimulated monocytes and neutrophils. It is very
important for cell-cell interactions which contribute to the
inflammatory response and also contribute to hemostasis. Several
clinical and experimental studies have suggested that the
expression of integrins on leukocytes is upregulated during or
after cardio-pulmonary bypass (CPB) (19–21).
Experimental data have demonstrated that cooling decreases the
upregulation of MAC-1 on monocytes (4,13).
The use of hypothermia has been shown to significantly reduce the
expression of MAC-1 during CPB compared with that during
normothermia in cardiac surgical patients (7). In the present investigation the
results revealed a significantly decreased expression of MAC-1 on
monocytes during conditions of DHCA compared with that during
normothermia. This suggests that hypothermia reduces the
upregulation of CD11b (MAC-1) expression on monocytes during
conditions of DHCA. This is consistent with data demonstrating that
hypothermia delays the production of proinflammatory cytokines and
NF-κB activation (5). Reduced
expression of MAC-1 on monocytes during circulatory arrest may be
associated with a decreased ability of the leukocytes to adhere to
endothelium and platelets, which is a prerequisite for reduced
leukocyte-induced tissue damage. Thus, hypothermia-induced
reduction of CD11b expression on monocytes is part of the
organ-protective effects during DHCA.
CD54 (ICAM-1) is a member of the immunoglobulin
superfamily. It is expressed on the cell surface of a wide variety
of cell types including endothelial cells and leukocytes,
functioning as a key receptor in the cell-cell interactions
(9,22). During inflammation, flowing
leukocytes roll onto vascular surfaces, arrest, spread, crawl to
endothelial junctions and then migrate into extravascular tissues.
Selectin-ligand interactions initiate rolling, whereas
integrin-ligand interactions mediate arrest and crawling. As the
neutrophils roll, P- or E-selectin expressed on activated
endothelial cells transduces signals that partially activate
integrin CD11a, which binds reversibly to ICAM-1 to decrease the
rolling velocities. Slow rolling facilitates neutrophil
interactions with endothelial cell-bound chemokines that fully
activate the integrins, leading to arrest (23). The binding of ICAM-1 to MAC-1
(CD11b/CD18) results in the adhesion of neutrophils and monocytes
to the endothelium (22). ICAM-1
is induced by cytokines and various stress stimuli such as hypoxia,
and is associated with a variety of inflammatory diseases and
conditions, including atherosclerosis and ischemia reperfusion
injury (9,22). Such inflammatory conditions are
present during DHCA. The NF-κB signaling cascade is pivotal in
ICAM-1 activation but NF-κB-independent pathways may also
participate (9). The NF-κB
signaling pathway is activated by the proinflammatory cytokines
tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), the
major inducers of ICAM-1 expression in most cell types (22). Experimental data demonstrate that
hypothermia delays the production of the proinflammatory cytokines
TNF-α and IL-1β as well as NF-κB activation (5). Thus, it may be speculated that
hypothermia decreases the expression of ICAM-1 on the cell surface
by attenuating the activation of proinflammatory cytokines and
NF-κB. In contrast to this hypothesis, a recent investigation
revealed no association between different blood temperatures and
the expression of CD54 on monocytes after 30 min of blood
circulation in an in vitro ECC model (13). In the present study, the effect of
deep hypothermia during blood stasis using an experimental setup
simulating DHCA was investigated. The data demonstrate no
significant difference in the surface expression of ICAM-1 (CD54)
on monocytes during normothermia and hypothermia. An explanation
for this may be that alternative activation pathways of ICAM-1
exist via NF-κB-independent pathways (9,22).
These NF-κB-independent signaling pathways are likely to be
unaffected by hypothermia. Nevertheless, the data demonstrate that
even though the expression of ICAM-1 on monocytes was unaffected by
hypothermia during stasis, the leukocyte-endothelial interaction
was reduced by deep hypothermia due to the decreased expression of
its ligand MAC-1 on monocytes. Thus, deep hypothermia leads to an
inhibited leukocyte response during circulatory arrest.
During acute inflammation, leukocytes are recruited
from the blood circulation to sites of infection and injury. This
multistep adhesion and signaling cascade is initiated by
interactions between selectins and their glycoconjugates that
mediate leukocyte tethering to and rolling on the surface of
endothelial cells. CD162 (PSGL-1) has been demonstrated to generate
a specific, high-affinity, biologically relevant ligand for
P-selectin (24,25). To trigger slow leukocyte rolling on
ICAM-1, neutrophils rolling on P-selectin engage PSGL-1 (23). In addition, ligation of PSGL-1 by
P-selectin may trigger intracellular events in some leukocytes
which enable them to respond to mediators elaborated at sites of
inflammation (24,25). P-selectin is an adhesion molecule,
which, besides generating a tether with PSGL-1, induces a signal
that activates leukocytes through a molecular cascade, finally
inducing the activated form of MAC-1. The P-selectin/MAC-1 cascade
gives rise to heterotypic conjugates of platelets with leukocytes
(18). It has been reported by
Straub et al, that hypothermia induces α-granule release
with increased expression of P-selectin on platelets, which
mediates platelet-leukocyte binding via interaction with the
leukocyte ligand PSGL-1. This finding reveals that hypothermia
induces platelet activation (1,11).
In a prior study, a decreased expression level of CD162 on
monocytes at 18°C was revealed using an in vitro ECC model
(13). In concordance with these
results, the current study demonstrated a decreased expression of
CD162 on monocytes at deep hypothermia during blood stasis. The
results of the present study suggest that hypothermia decreases
PSGL-1 expression on monocytes resulting in a decreased
platelet-leukocyte interaction. Additionally, hypothermia-induced
reduction of PSGL-1 on monocytes may further have an impact on
leukocyte-endothelial interaction, such as the tethering and
rolling of leukocytes, which affects the inflammatory response.
These data suggest that deep hypothermia has a beneficial effect on
the systemic inflammatory response and organ damage during
circulatory arrest.
In conclusion, to the best of our knowledge, the
findings of the present study reveal for the first time that deep
hypothermia decreases the expression of CD11b (MAC-1) and CD162
(PSGL-1) on monocytes in an experimental setup simulating the
conditions of DHCA. This may result in an inhibition of
leukocyte-endothelial and leukocyte-platelet interactions, which
may be a beneficial aspect of deep hypothermia that may influence
the inflammatory response and tissue damage during DHCA.
References
|
1
|
Straub A, Azevedo R, Beierlein W, Wendel
HP, Dietz K and Ziemer G: Glycoprotein IIb/IIIa inhibition reduces
prothrombotic events under conditions of deep hypothermic
circulatory arrest. Thromb Haemost. 94:115–122. 2005.PubMed/NCBI
|
|
2
|
Ziganshin BA and Elefteriades JA: Deep
hypothermic circulatory arrest. Ann Cardiothorac Surg. 2:303–315.
2013.
|
|
3
|
Mossad EB, Machado S and Apostolakis J:
Bleeding following deep hypothermia and circulatory arrest in
children. Semin Cardiothorac Vasc Anesth. 11:34–46. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rimmelé T, Bishop J, Simon P, et al: What
blood temperature for an ex vivo extracorporeal circuit? Artif
Organs. 35:593–601. 2011.PubMed/NCBI
|
|
5
|
Kimura A, Sakurada S, Ohkuni H, Todome Y
and Kurata K: Moderate hypothermia delays proinflammatory cytokine
production of human peripheral blood mononuclear cells. Crit Care
Med. 30:1499–1502. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
González-Ibarra FP, Varon J and López-Meza
EG: Therapeutic hypothermia: critical review of the molecular
mechanisms of action. Front Neurol. 2:42011.PubMed/NCBI
|
|
7
|
Le Deist F, Menasché P, Kucharski C, Bel
A, Piwnica A and Bloch G: Hypothermia during cardiopulmonary bypass
delays but does not prevent neutrophil-endothelial cell adhesion. A
clinical study. Circulation. 92(9 Suppl): II354–II358.
1995.PubMed/NCBI
|
|
8
|
Lin L, Wu C and Hu K: Tissue plasminogen
activator activates NF-κB through a pathway involving annexin
A2/CD11b and integrin-linked kinase. J Am Soc Nephrol.
23:1329–1338. 2012.
|
|
9
|
Gorgoulis VG, Zacharatos P, Kotsinas A, et
al: p53 activates ICAM-1 (CD54) expression in an
NF-kappaB-independent manner. EMBO J. 22:1567–1578. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simon DI, Ezratty AM, Francis SA, Rennke H
and Loscalzo J: Fibrin(ogen) is internalized and degraded by
activated human monocytoid cells via Mac-1 (CD11b/CD18): a
nonplasmin fibrinolytic pathway. Blood. 82:2414–2422.
1993.PubMed/NCBI
|
|
11
|
Straub A, Breuer M, Wendel HP, Peter K,
Dietz K and Ziemer G: Critical temperature ranges of
hypothermia-induced platelet activation: possible implications for
cooling patients in cardiac surgery. Thromb Haemost. 97:608–616.
2007.
|
|
12
|
Walter T, Suselbeck T, Borggrefe M,
Swoboda S, Hoffmeister HM and Dempfle CE: Effect of atorvastatin on
cellular adhesion molecules on leukocytes in patients with
normocholesterolemic coronary artery disease. In Vivo. 24:189–193.
2010.PubMed/NCBI
|
|
13
|
Swoboda S, Gruettner J, Lang S, et al:
Hypothermia inhibits expression of CD11b (MAC-1) and CD162 (PSGL-1)
on monocytes during extracorporeal circulation. In Vivo.
27:459–464. 2013.PubMed/NCBI
|
|
14
|
Straub A, Schiebold D, Wendel HP, Azevedo
R, Dietz K and Ziemer G: Platelet anaesthesia during extracorporeal
circulation: differential effects of GP IIb/IIIa blockers on
platelet activation marker P-selectin expression at hypothermia.
Thromb Res. 122:383–389. 2008. View Article : Google Scholar
|
|
15
|
Griepp RB and Di Luozzo G: Hypothermia for
aortic surgery. J Thorac Cardiovasc Surg. 145(3 Suppl): S56–S58.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tassani P, Barankay A, Haas F, et al:
Cardiac surgery with deep hypothermic circulatory arrest produces
less systemic inflammatory response than low-flow cardiopulmonary
bypass in newborns. J Thorac Cardiovasc Surg. 123:648–654. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mayadas TN and Cullere X: Neutrophil beta2
integrins: moderators of life or death decisions. Trends Immunol.
26:388–395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cerletti C, Tamburrelli C, Izzi B,
Gianfagna F and de Gaetano G: Platelet-leukocyte interactions in
thrombosis. Thromb Res. 129:263–266. 2012. View Article : Google Scholar
|
|
19
|
Tárnok A, Bocsi J, Rössler H, Schlykow V,
Schneider P and Hambsch J: Low degree of activation of circulating
neutrophils determined by flow cytometry during cardiac surgery
with cardiopulmonary bypass. Cytometry. 46:41–49. 2001.PubMed/NCBI
|
|
20
|
Greilich PE, Brouse CF, Rinder CS, et al:
Effects of epsilon-aminocaproic acid and aprotinin on
leukocyte-platelet adhesion in patients undergoing cardiac surgery.
Anesthesiology. 100:225–233. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Asimakopoulos G, Kohn A, Stefanou DC,
Haskard DO, Landis RC and Taylor KM: Leukocyte integrin expression
in patients undergoing cardiopulmonary bypass. Ann Thorac Surg.
69:1192–1197. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roebuck KA and Finnegan A: Regulation of
intercellular adhesion molecule-1 (CD54) gene expression. J Leukoc
Biol. 66:876–888. 1999.PubMed/NCBI
|
|
23
|
Shao B, Yago T, Coghill PA, et al:
Signal-dependent slow leukocyte rolling does not require
cytoskeletal anchorage of P-selectin glycoprotein ligand-1 (PSGL-1)
or integrin αLβ2. J Biol Chem. 287:19585–19598. 2012.
|
|
24
|
Zhang Y, Jiang N, Zarnitsyna VI, Klopocki
AG, McEver RP and Zhu C: P-selectin glycoprotein ligand-1 forms
dimeric interactions with E-selectin but monomeric interactions
with L-selectin on cell surfaces. PLoS One. 8:e572022013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moore KL: Structure and function of
P-selectin glycoprotein ligand-1. Leuk Lymphoma. 29:1–15. 1998.
View Article : Google Scholar : PubMed/NCBI
|