Introduction
The inflammatory response induced by
hypoxic-ischemic (HI) brain damage may further trigger the
activation of certain signaling pathways, finally leading to cell
apoptosis or necrosis (1,2). Various complex chemical
neurotransmitters and signaling pathways are involved in this
process. A large number of studies have confirmed that the
phosphatidylinositol 3-kinase (PI3K)-protein kinase B (Akt)
signaling pathway is one of the major pathways involved in neuronal
apoptosis in the brain (3,4). The PI3K/Akt signaling pathway is an
important mode of transducing cell membrane receptor signals into
intracellular signals, and thus serves a key function in
maintaining cell survival and inhibiting apoptosis. This pathway is
also involved in regulating cell proliferation and inhibiting
apoptosis by affecting the activation of effector molecules, such
as cyclin and apoptosis-related proteins downstream (4).
Glycogen synthase kinase-3β (GSK-3β) is an essential
protein downstream of Akt that is involved in the pathological
process of HI brain damage. The PI3K/Akt signal transduction
pathway may affect the activity of GSK-3β. Activated Akt inhibits
GSK-3β activity via phosphorylation (5). A study of cerebral ischemic injury
has confirmed that GSK-3β serves a pro-apoptotic function in HI
brain injury by activating the p53 gene that expresses the P53
protein. P53 protein inhibits anti-apoptotic Bcl-2 and activates
pro-apoptotic Bax, thereby promoting apoptosis. GSK-3β may also
inhibit heat shock factor 1 (HSF 1) to decrease HSP70 expression
and promote apoptosis (6).
Therefore, neuronal apoptosis due to HI brain injury in neonatal
rats may be alleviated through intervention in the PI3K/Akt/GSK-3β
signaling pathway, which may provide new solutions for the
treatment of brain injury.
Progesterone (PROG) is generated by endocrine
tissues outside the nervous system and acts on reproductive organs
to provide a natural regulating effect on reproductive function.
However, recent studies suggesting protective effects of PROG on
the brain, in spinal cord injury, as well as in peripheral nerve
repair after injury, have aroused considerable interest (7–9). A
number of studies have found that PROG can reduce brain edema and
scavenge free radicals, as well as function as an antioxidant to
enhance the cognitive abilities of animals with brain injury
(10–12). However, the specific brain
protection mechanism of PROG remains unclear, and studies of brain
damage in newborn rats are considerably limited. To the best of our
knowledge, current literature has not reported on whether PROG
alleviates the neuronal apoptosis associated with HI brain damage
in neonatal rats by modulating GSK-3β expression via activation of
the PI3K/Akt/GSK-3β signaling pathway. Studies investigating this
are thus warranted.
In the present study, molecular biology
technologies, including immunohistochemistry, western blotting and
enzyme-linked immunosorbent assay (ELISA), were used to explore the
mechanism by which the PI3K/Akt/GSK-3β pathway induces
neuroprotective effects following PROG intervention in HI brain
injury. This study also aimed to further clarify the molecular
mechanism of the protective effect of PROG and provide experimental
and theoretical evidence relevant to the clinical therapeutic
applications of PROG.
Materials and methods
Animal source and grouping
Ninety-six male and female Wistar rats, aged 7 days
and weighing 12–18 g were provided by the Experimental Animal
Centre of Xinxiang Medical University (Xinxiang, China). The
animals were randomly divided into four groups of 24: Sham surgery,
HI, drug prevention (PROG) and Akt inhibitor groups. Neck incision
without ischemia and hypoxia treatment was performed on animals in
the sham group. Animals in the HI group were subjected to the
animal model preparation process described below. Animals in the
drug prevention group were subjected to the animal modeling process
and injected with 0.5 g/l PROG solution (Sigma-Aldrich, St. Louis,
MO, USA) intraperitoneally at a dose of 8 mg/kg 30 min prior to
hypoxia. Animals in the Akt inhibitor group were also subjected to
the animal modeling process. However, 30 min prior to model
establishment, the rats underwent positioning of the left
hippocampus using a brain stereotaxic apparatus, and were injected
with dilute (0.16 μg/μl) wortmannin solution (Sigma-Aldrich) into
the left hippocampus at a dose of 16 μg/kg. This study was carried
out in strict accordance with the recommendations in the Guide for
the Care and Use of Laboratory Animals of the National Institutes
of Health: Eight Edition (2010). The animal use protocol was
reviewed and approved by the Institutional Animal Care and Use
Committee (IACUC) of Xinxiang Medical University.
Animal model preparation
Following isoflurane anesthesia, supine neonatal
rats were fixed on a rat bench. The skin of the neck was sliced in
the middle following disinfection with alcohol, and then the left
common carotid artery was isolated from within the
sternocleidomastoid muscle, ligated with silk and cut from the
middle. The wounds were sutured, and the animals were left to
recover at room temperature for 2–3 h. Animals without rotational
movement of the left side were removed and placed in a sealed
container at a constant temperature of 37°C, and ventilated with 8%
O2 and 92% N2 gas mixture at a rate of 1.5
l/min for 2.5 h to prepare the HI animal models (13). In the sham group, the left common
carotid artery was isolated only, and was not subjected to ligation
or the hypoxia treatment.
Immunohistochemistry
Experimental animals were sacrificed 24 h after
hypoxia-ischemia, and their brains were immediately taken,
segmented from the optic chiasm, placed in 10% formalin and fixed
overnight. Following conventional dehydration, the brains were
transparently embedded in paraffin and sliced into 5-μm sections.
Finally, the slices were dewaxed to water, baked and preserved in a
4°C refrigerator in readiness for p-Akt and GSK-3β
immunohistochemical staining.
The streptavidin biotin-peroxidase complex (SABC)
immunohistochemical technique was used. p-Akt (Ser 473) and GSK-3β
immunohistochemical staining was performed in strict accordance
with the instructions of the reagent kit provided by Wuhan Boster
Biotechnology Co., Ltd. (Wuhan, China). In the negative control
assay, phosphate-buffered saline (PBS) replaced the antibody, and
the remaining procedures were conducted as described in the kit’s
instructions. The positive expression of p-Akt and GSK-3β proteins
was indicated by the cytoplasm and membrane of the cells being
stained brownish yellow. Two different fields of the cortex of
three animal brain tissue slices were observed under a light
microscope (magnification, ×400), and images were randomly captured
using a digital camera. An HMIAS-2000 color image analysis system
(Tongji Medical College Clear Screen Imaging Inc., Wuhan, China)
was used to analyze the staining results and to calculate the mean
optical density (MOD).
Western blot analysis
In total, eight animals from each group were
sacrificed 24 h after hypoxia-ischemia; their brains were
immediately placed on ice and part of the cortex in the
brain-damaged side was obtained for protein detection. After adding
cell lysis buffer, the proteins were extracted by centrifugation,
with the total tissue proteins being contained in the supernatant.
The protein concentration was detected using the bicinchoninic acid
(BCA) method. Polyacrylamide gel electrophoresis was performed to
isolate the target protein, which was transferred to
nitrocellulose. Rabbit anti-rat p-Akt (Ser 473), rabbit anti-rat
GSK-3β or anti-β-actin antibodies were added as the primary
antibodies after sealing, and the kit used was provided by Beijing
Zhongshan Golden Bridge Biotechnology Co., Ltd. (Beijing, China).
The secondary antibody (Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA) was added and the sample was incubated at 4°C overnight. A
gel or film transient display system was used to scan the films,
and image analysis software (Tocan, Shanghai, China) was used to
analyze the optical density value of the target bands. The relative
protein content was expressed as the grey ratio of the band
representing the target protein to that of the band denoting the
internal reference protein.
Flow cytometry
Flow cytometry was used to determine the apoptosis
rate of neural cells. Ischemic brain tissues of eight mice from
each group were rapidly stripped on ice. Single-cell suspensions
were prepared mechanically, centrifuged at 4°C and 1,200 × g for 10
min, and then the supernatant was discarded. The cells were
suspended in 1 ml cold PBS by gentle shaking, centrifuged at 4°C
and 1,000 rpm for 10 min, and then the supernatant was discarded.
The cells were resuspended in 200 μl binding buffer and 10 μl
Annexin V-fluorescein isothiocyanate (FITC) and 5 μl propidium
iodide (PI) were then added. The cell suspension was gently mixed
and stored in the dark at room temperature for 15 min. Finally, 300
μl binding buffer was added to the suspension, which was analyzed
using flow cytometry (FACS 420; BD Biosciences, Franklin Lakes, NJ,
USA).
ELISA
A total of eight animals in each group were
sacrificed 24 h after hypoxia-ischemia. Following sacrifice, the
craniotomy raphe nucleus, stripped pia mater and ischemic brain
tissue were placed in a homogenizer. Following the addition of 2 ml
PBS solution to prepare a tissue homogenate, the tissues were
centrifuged at 1,000 rpm for 10 min. The supernatant was obtained,
the number of cells was adjusted to between 105 and
106, and then the GSK-3β content was measured by ELISA.
The kit was provided by Adlitteram Diagnostic Laboratories (San
Diego, CA, USA). A Hyperion MR III microplate reader was used
(Biotek Instruments Inc., Winooski, VT, USA) and the process was
conducted strictly in accordance with the instructions of the
manufacturer.
Statistical analysis
All experimental data were analyzed using SPSS 13.0
(SPSS, Inc., Chicago, IL, USA). Measurement data are expressed as
means ± standard deviations and comparisons between different
groups were analyzed using one-way analysis of variance (ANOVA).
Paired comparisons between two groups were analyzed using a t-test,
with P<0.05 considered to indicate a statistically significant
difference.
Results
Immunohistochemistry
Positive staining for p-Akt was revealed as
irregularly shaped brown particles, mainly located in the cytoplasm
and some in the nucleus. p-Akt-positive cells were occasionally
seen in the sham group with light staining, a sparse distribution
and low MOD. The expression of p-Akt in the HI group was
considerably less, with light staining, a sparse distribution and a
lower MOD than that of the sham group; however, the difference
between the two groups was not statistically significant
(P>0.05). p-Akt expression in the PROG group was the most
immunoreactive, with positive particles distributed densely with
dark staining and a significantly increased MOD compared with that
of the HI group (P<0.05). p-Akt expression in the Akt inhibitor
group was significantly decreased compared with that of the PROG
group (P<0.05; Table I;
Fig. 1A–D).
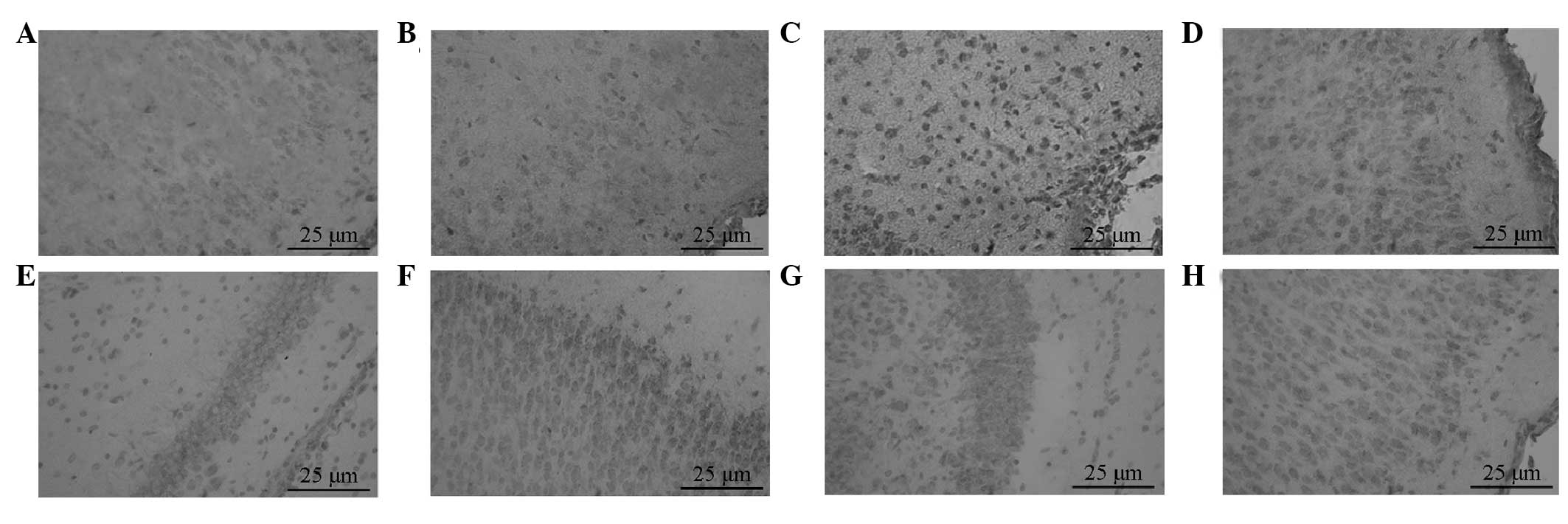 | Figure 1Expression of p-Akt and GSK-3β was
observed using immunohistochemistry (magnification, ×400). p-Akt
expression in the (A) sham, (B) HI, (C) PROG and (D) Akt inhibitor
groups, respectively. GSK-3β expression in the (A) sham, (B) HI,
(C) PROG and (D) Akt inhibitor groups, respectively. HI,
hypoxic-ischemic; PROG, progesterone; p-Akt, phosphorylated Akt;
GSK-3β, glycogen synthase kinase 3β. |
 | Table IEffect of PROG on the expression of
p-Akt and GSK-3β in the brain tissue of neonatal rats. |
Table I
Effect of PROG on the expression of
p-Akt and GSK-3β in the brain tissue of neonatal rats.
|
Immunohistochemistry | Western blotting |
|---|
|
|
|
|---|
| Group | p-Akt | GSK-3β | p-Akt | GSK-3β |
|---|
| Sham | 0.386±0.022 | 0.262±0.025 | 0.21±0.04 | 0.24±0.03 |
| HI | 0.342±0.031 | 0.738±0.035a | 0.19±0.06 | 0.49±0.12a |
| PROG | 0.754±0.046b | 0.432±0.028b | 0.55±0.16b | 0.25±0.02b |
| Akt inhibitor | 0.429±0.035c | 0.526±0.021c | 0.34±0.06c | 0.40±0.08c |
GSK-3β-positive cells were round- or oval-shaped and
were widely distributed in the neurons of the cerebral cortex.
GSK-3β expression was infrequently seen in the sham group, and the
MOD in this group was low. The expression of GSK-3β in the HI group
was significantly higher than that in the sham group (P<0.05).
The MOD of the PROG group was significantly lower than that of the
HI group, whereas the expression of GSK-3β in the Akt inhibitor
group was significantly higher than that in the PROG group
(P<0.05; Table I; Fig. 1E–H).
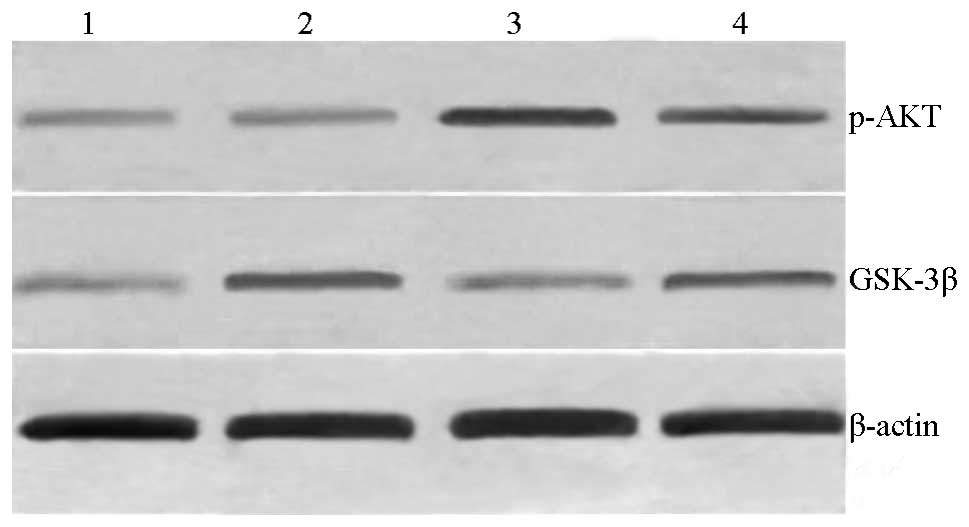
Western blotting
The western blot analysis results showed that the
molecular weight of p-Akt was ~60 kDa. Only a small amount of p-Akt
was expressed in the brain tissue of the sham group. The grey ratio
of the p-Akt bands with respect to the β-actin internal reference
bands in the sham group was insignificantly lower. The p-Akt
protein expression level of the PROG group was significantly
increased compared with that of the HI group (P<0.05). The level
of p-Akt expression in the rat brain tissues was significantly
lower compared with that of the PROG group following the use of the
Akt inhibitor, and the difference was statistically significant
(P<0.05; Table I; Fig. 2). The western blotting results were
comparable with the immunohistochemical results for p-Akt in these
groups.
The level of GSK-3β expression was low in the sham
group, and that of the HI group was significantly higher compared
with that of the sham group (P<0.05). The expression level of
GSK-3β in the PROG group was significantly lower than that in the
HI group (P<0.05). Following the use of the Akt inhibitor, the
GSK-3β expression level in the rat brain tissues was significantly
increased compared with that of the PROG group, and the difference
was statistically significant (P<0.05; Table I; Fig.
2).
GSK-3β content and neuronal apoptosis
rate
The GSK-3β content as determined by ELISA and the
neuronal apoptosis rate in the brain tissues of newborn rats in the
HI group were significantly higher than those of the sham group
(P<0.05). The GSK-3β content and neuronal apoptosis rate in the
brain tissue of the PROG-treated group was significantly lower than
that of the HI group (P<0.05), whereas the values of in the Akt
inhibitor group were significantly higher than those of the PROG
group (P<0.05; Table II).
| Table IIEffects of PROG on the levels of
GSK-3β and rate of apoptosis in brain tissue following HI brain
damage in neonatal rats. |
Table II
Effects of PROG on the levels of
GSK-3β and rate of apoptosis in brain tissue following HI brain
damage in neonatal rats.
| Group | GSK-3β (ng/ml) | Apoptosis rate |
|---|
| Sham | 3.26±0.21 | 2.49±0.23 |
| HI | 7.87±0.59a | 10.09±0.36a |
| PROG | 5.10±0.42b | 3.47±0.32b |
| Akt inhibitor | 6.65±0.37c | 6.32±0.56c |
Discussion
HI brain injury is a critical illness in the
neonatal period. Infants are prone to mortality, and survivors
easily retain neurological sequelae, which seriously affect their
health and quality of life. The pathogenesis of HI brain injury is
not fully understood, and no effective treatment is available
(14). Therefore, the
identification of methods to weaken the effects of cerebral
ischemia and protect against neuronal injury is necessary. The
pathophysiology of HI brain injury is a complex, multiply linked,
multi-factorial, multi-channel cascade of enzymatic activity and
injury processes (15), of which
nerve cell apoptosis is a major factor. Activation of signal
transduction processes is a necessary prerequisite for the
initiation of apoptosis.
Studies have suggested that the PI3K/Akt signaling
pathway is the main apoptosis-associated signal transduction
pathway that is involved following cerebral ischemia, and that this
signaling pathway is important in cell survival (4,16).
Under physiological conditions, Akt is located in the cytoplasm in
a low-activity form; when cells are stimulated by an extracellular
signal (for example, ischemia or hypoxia), the C-terminal
Ser 473 residues of Akt are activated by phosphorylation for the
regulation of the upstream molecule. The activated Akt promotes
cell survival through a variety of mechanisms, and one of the main
mechanisms involves the inhibition of GSK-3β activity to prevent it
from exerting an apoptotic effect (17,18).
That is, the activated Akt binds to GSK-3β, induces the
translocation of GSK-3β to the cell membrane, phosphorylates the
N-terminal Ser active site, and thereby inactivates GSK-3β
(19,20). Therefore, the identification of Akt
and GSK-3β as targets for the inhibition of neuronal apoptosis is
of great significance for reducing brain damage via the use of
neuroprotective agents.
PROG contributes to the protection of brain tissue
in the pathogenesis of HI brain injury, and the authors’ previous
studies have shown that PROG may antagonize the production of free
radicals, inhibit apoptosis, among other activities (9,21).
However, the molecular mechanisms and pathways by which PROG
inhibits apoptosis remain unclear. Whether the protective effect of
PROG against the apoptosis of damaged neurons in the HI brain is
realized by modulation of the PI3K/Akt/GSK-3β pathway was worthy of
study.
Different degrees of ischemia and various ischemia
models generated via the Akt/GSK-3β pathway following cerebral
ischemia show different disease states (22,23).
Previous studies (23,24) found that p-Akt expression in the
neurons of rat models following middle cerebral artery occlusion
increased in the 1–3 h after ischemia and reperfusion, with the
peak occurring at 1 h, indicating that the PI3K/Akt signaling
pathway is involved in the brain ischemia-induced stress response
in early ischemia-reperfusion. In transient global cerebral
ischemia and focal cerebral ischemia models, the level of p-Akt
increased to help to protect neurons against fatal ischemic damage.
Following use of a PI3K/Akt inhibitor, ischemic injury worsened.
GSK-3β is activated to promote apoptosis in 90-min focal cerebral
ischemia models (severe ischemia). Conversely, GSK-3β is
inactivated to promote cell survival in short 5-min global ischemia
models (mild cerebral ischemia models). Since previous studies
(24,25) have shown that the first 24 h for
newborn rats with hypoxic brain damage is the most critical, in
which the peak time of neuronal apoptosis and the most serious
stage of aggravated secondary brain injury occurs, 24 h HI
following brain injury was selected as the observation time point
in the present study. Changes in the PI3K/Akt/GSK-3β pathway at 24
h following HI brain damage in the neonatal rat brain tissue were
observed, and a PI3K/Akt pathway inhibitor, wortmannin, was used to
hinder HI brain damage in neonatal rats (wortmannin is a
phosphatase inhibitor that prevents Akt phosphorylation and
activity) to explore the neuroprotective effect mechanism of the
PI3K/Akt/GSK-3β pathway in the process by which PROG attenuates HI
brain injury.
To investigate the influence of the Akt/GSK-3β
pathway on the HI brain tissue of the neonatal rats,
immunohistochemistry and western blotting were used in the present
study to detect p-Akt and GSK-3β expression in the brain tissues 24
h following cerebral ischemia, ELISA was used to determine the
brain tissue GSK-3β content, and flow cytometry to determine the
rate of neuronal apoptosis. p-Akt expression levels were found to
be decreased and GSK-3β expression levels were increased in 24 h HI
brain tissues. Similarly, the GSK-3β content and neuronal apoptosis
rate were elevated, indicating that GSK-3β expression increased to
promote apoptosis in the late HI brain period. These results are
consistent with those of previous studies (19,20),
which suggest that the phosphorylation of Akt decreases following
long-term cerebral ischemia, and that GSK-3β is activated to
promote apoptosis. Following PROG administration, the highest p-Akt
expression level was observed in the PROG group, which exhibited a
dense immunoreactive particle distribution, dark staining and
significantly increased MOD values compared with those in the HI
group. GSK-3β expression levels were significantly reduced, as well
as the GSK-3β content of brain tissue, and the neuronal apoptosis
rate was lowered, indicating that PROG is able to elevate the
levels of p-Akt, inhibit GSK-3β expression and reduce its activity
to prevent apoptosis, thus further decreasing HI brain damage. The
current study further investigated whether the protective effect of
PROG in the brain was achieved through the PI3K/Akt/GSK-3β
signaling pathway. Following the intraventricular injection of
wortmannin to inhibit the PI3K/Akt pathway, the expression level of
p-Akt in the Akt inhibitor group was significantly decreased
compared with that of the PROG group, the expression level of
GSK-3β was significantly higher than that of the PROG group, and
the GSK-3β content and neuronal apoptosis rate were significantly
higher than those of the PROG group. These results indicate that
the injection of the Akt inhibitor wortmannin aggravates brain
damage and confirms that PROG can exert a protective effect during
ischemic brain injury through the activation of the PI3K/Akt/GSK-3β
pathway, and that the Akt/GSK-3β signaling pathway may mediate the
protective effect of PROG on neurons. The protective effect of PROG
against brain injury may occur through multiple pathways;
therefore, further studies on the underlying associated mechanism
are required to provide a theoretical basis for the future
prevention and treatment of HI brain disease.
In conclusion, Akt protein activity was decreased in
newborn rats in the 24 h following HI brain damage, thereby
triggering the activity of its downstream effector GSK-3β, which
may be one of the signaling pathways underlying neuronal apoptosis.
PROG is able to increase Akt activity, inhibit GSK-3β activity and
decrease brain damage. Therefore, PROG is indicated to exert a
protective effect on ischemic brain injury by activating the
PI3K/Akt/GSK-3β pathway, which provides an experimental basis for
its potential use as a drug for treating ischemic brain injury.
Acknowledgements
This study was supported by the Natural Science
Research Project of Education Department of Henan Province
(2008A180029) and the Funding Program for Young Backbone Teachers
in Colleges and Universities of Henan (2012GGJS-134).
References
|
1
|
Liu F and McCullough LD: Inflammatory
responses in hypoxic ischemic encephalopathy. Acta Pharmacol Sin.
34:1121–1130. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kasdorf E and Perlman JM: Hyperthermia,
inflammation, and perinatal brain injury. Pediatr Neurol. 49:8–14.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gu Q, Zhai L, Feng X, et al: Apelin-36, a
potent peptide, protects against ischemic brain injury by
activating the PI3K/Akt pathway. Neurochem Int. 63:535–540. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang B, Ji X, Zhang S, Ren H, Wang M, Guo
C and Li Y: Hemin-mediated neuroglobin induction exerts
neuroprotection following ischemic brain injury through PI3K/Akt
signaling. Mol Med Rep. 8:681–685. 2013.
|
|
5
|
Xiong T, Tang J, Zhao J, et al:
Involvement of the Akt/GSK-3β/CRMP-2 pathway in axonal injury after
hypoxic-ischemic brain damage in neonatal rat. Neuroscience.
216:123–132. 2012.
|
|
6
|
Valerio A, Bertolotti P, Delbarba A, et
al: Glycogen synthase kinase-3 inhibition reduces ischemic cerebral
damage, restores impaired mitochondrial biogenesis and prevents ROS
production. J Neurochem. 116:1148–1159. 2011. View Article : Google Scholar
|
|
7
|
Singh M and Su C: Progesterone-induced
neuroprotection: factors that may predict therapeutic efficacy.
Brain Res. 1514:98–106. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baudry M, Bi X and Aguirre C:
Progesterone-estrogen interactions in synaptic plasticity and
neuroprotection. Neuroscience. 239:280–294. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Zhang J, Yang Y, Dong W, Wang F,
Wang L and Li X: Progesterone attenuates cerebral edema in neonatal
rats with hypoxic-ischemic brain damage by inhibiting the
expression of matrix metalloproteinase-9 and aquaporin-4. Exp Ther
Med. 6:263–267. 2013.PubMed/NCBI
|
|
10
|
Sarkaki AR, Khaksari Haddad M, Soltani Z,
Shahrokhi N and Mahmoodi M: Time- and dose-dependent
neuroprotective effects of sex steroid hormones on inflammatory
cytokines after a traumatic brain injury. J Neurotrauma. 30:47–54.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsuji M, Taguchi A, Ohshima M, Kasahara Y
and Ikeda T: Progesterone and allopregnanolone exacerbate
hypoxic-ischemic brain injury in immature rats. Exp Neurol.
233:214–20. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shahrokhi N, Khaksari M, Soltani Z,
Mahmoodi M and Nakhaee N: Effect of sex steroid hormones on brain
edema, intracranial pressure, and neurologic outcomes after
traumatic brain injury. Can J Physiol Pharmacol. 88:414–421. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Taniguchi H and Andreasson K: The
hypoxic-ischemic encephalopathy model of perinatal ischemia. J Vis
Exp. 19:3791–3792. 2008.
|
|
14
|
Doi K, Sameshima H, Kodama Y, Furukawa S,
Kaneko M and Ikenoue TK: Perinatal death and neurological damage as
a sequential chain of poor outcome. J Matern Fetal Neonatal Med.
25:706–709. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Northington FJ, Chavez-Valdez R and Martin
LJ: Neuronal cell death in neonatal hypoxia-ischemia. Ann Neurol.
69:743–758. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu YM, Wang CC, Chen L, et al: Both
PI3K/Akt and ERK1/2 pathways participate in the protection by
dexmedetomidine against transient focal cerebral
ischemia/reperfusion injury in rats. Brain Res. 494:1–8. 2013.
View Article : Google Scholar
|
|
17
|
Zhang X, Zhang X, Wang C, et al:
Neuroprotection of early and short-time applying berberine in the
acute phase of cerebral ischemia: up-regulated pAkt, pGSK and
pCREB, down-regulated NF-κB expression, ameliorated BBB
permeability. Brain Res. 1459:61–70. 2012.PubMed/NCBI
|
|
18
|
Zhou C, Tu J, Zhang Q, et al: Delayed
ischemic postconditioning protects hippocampal CA1 neurons by
preserving mitochondrial integrity via Akt/GSK3β signaling.
Neurochem Int. 59:749–758. 2011.PubMed/NCBI
|
|
19
|
Sun B, Chen L, Wei X, Xiang Y, Liu X and
Zhang X: The Akt/GSK-3β pathway mediates flurbiprofen-induced
neuroprotection against focal cerebral ischemia/reperfusion injury
in rats. Biochem Biophys Res Commun. 409:808–813. 2011.
|
|
20
|
Collino M, Aragno M, Castiglia S,
Tomasinelli C, Thiemermann C, Boccuzzi G and Fantozzi R: Insulin
reduces cerebral ischemia/reperfusion injury in the hippocampus of
diabetic rats: a role for glycogen synthase kinase-3beta. Diabetes.
58:235–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang XY, Li XJ, Li DL, Li XJ, Zhu XQ and
Guo XP: Neuroprotective effect of progesterone in newborn rats with
hypoxic-ischemic encephalopathy. Int J Phys Sci. 6:2894–2900.
2011.
|
|
22
|
Zhao H, Shimohata T, Wang JQ, Sun G,
Schaal DW, Sapolsky RM and Steinberg GK: Akt contributes to
neuroprotection by hypothermia against cerebral ischemia in rats. J
Neurosci. 25:9794–9806. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Endo H, Nito C, Kamada H, Nishi T and Chan
PH: Activation of the Akt/GSK3beta signaling pathway mediates
survival of vulnerable hippocampal neurons after transient global
cerebral ischemia in rats. J Cereb Blood Flow Metab. 26:1479–1489.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li L, Qu Y, Mao M, Xiong Y and Mu D: The
involvement of phosphoinositide 3-kinase/Akt pathway in the
activation of hypoxia-inducible factor-1alpha in the developing rat
brain after hypoxia-ischemia. Brain Res. 1197:152–158. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yao D, He X, Wang JH and Zhao ZY: Effects
of PI3K/Akt signaling pathway on learning and memory abilities in
neonatal rats with hypoxic-ischemic brain damage. Zhongguo Dang Dai
Er Ke Za Zhi. 13:424–427. 2011.(In Chinese).
|