Introduction
Artemisinin is a sesquiterpene lactone isolated from
the Artemisia annua plant that has been used extensively as
an antimalarial drug (1).
Dihydroartemisinin (DHA) is the active metabolite of artemisinin
compounds, and a semisynthetic derivative of artemisinin (2). DHA is an effective, water-soluble
antimalarial drug, with fewer side-effects compared with other
drugs (3). In addition, DHA
exhibits strong antitumor and antiangiogenesis effects (4). However, the underlying mechanisms of
DHA remain poorly understood.
Angiogenesis, the formation of new capillaries
branching from existing vessels, plays a critical role in embryonic
development, wound-healing and the menstrual cycle (5). Abnormal angiogenesis is associated
with tumor growth, diabetes, rheumatoid arthritis and
atherosclerosis (6). Under hypoxic
conditions, tumor cells or other growing cells secrete growth
factors, including vascular endothelial growth factor (VEGF) and
basic fibroblast growth factor (bFGF). These growth factors
activate endothelial cells (ECs) in nearby capillaries, leading to
the migration of ECs branching out of the vessel (7). The cells proliferate and
differentiate to form a network of new capillaries. It has been
well established that solid tumor growth requires a neovascular
network, which enables rapid proliferation of tumor cells by
providing oxygen and nutrients (8).
EC migration is an essential component of
angiogenesis (7). This motile
process is regulated by chemotactic, haptotactic and mechanotactic
stimuli (7). Typically, chemotaxis
of ECs is driven by growth factors, including VEGF and bFGF,
whereas haptotaxis is associated with increased EC motility in
response to integrins binding to the extracellular matrix (5). The process requires the activation of
several signaling pathways that converge on cytoskeletal remodeling
(9).
Mitogen-activated protein kinase (MAPK) pathways
constitute a large network of signaling cascades that regulate
diverse physiological processes, including cell migration (10). The functions of MAPKs are mediated
through the phosphorylation of substrates, including
phospholipases, transcription factors and cytoskeletal proteins
(11). To date, three major
subfamilies of MAPKs have been well-characterized: Extracellular
signal-regulated kinases (ERKs), p38 MAPKs and c-Jun N-terminal
kinases (JNKs). The p38 pathway conveys the VEGF signal to
microfilaments, inducing rearrangements in the actin cytoskeleton
that regulate cell migration (12). Through the modulation of cell
migration, p38 MAPK is an important regulator of angiogenesis
(13).
In the present study, DHA was hypothesized to
inhibit EC migration via the p38 MAPK pathway. Cell migration
assays were performed on human umbilical vein ECs (HUVECs) with the
addition of DHA, and the phosphorylation of the p38 MAPK was
examined by western blot analysis. The role of p38 MAPK was further
examined using an electrical cell-substrate impedance sensing
(ECIS) system with the specific inhibitor, SB203850.
Materials and methods
Cell culture
HUVECs were purchased from the American Type Culture
Collection (Manassas, VA, USA) and maintained in endothelial growth
medium (EGM-2), supplemented with the EGM-2-MV bullet kit (Lonza,
Basel, Switzerland) and antibiotics (100 IU/ml penicillin and 100
μg/ml streptomycin), in a humidified atmosphere at 37°C and 5%
CO2. DHA (sc-211332; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA), anisomycin (sc-3524, Santa Cruz
Biotechnology, Inc.) and SB203850 (Cell Signaling Technology, Inc.,
Beverly, MA, USA) were dissolved in dimethyl sulfoxide (DMSO).
Boyden chamber migration assay
Cell migration assays were performed using modified
24-well Boyden chambers (Costar, Acton, MA, USA), containing a
polycarbonate membrane with 8.0-μm pores. HUVECs were starved in
basic EGM-2 (serum/growth factor free) overnight at 37°C, prior to
being harvested with trypsin and resuspended in basic EGM-2. The
single cell suspensions with 20 μM DHA or 20 μM SB203850 were
seeded at 1×105 cells/well in the upper chamber, while
0.5 ml EGM-2 with 20 ng/ml VEGF was added to the bottom chamber as
chemoattractants. After 24 h incubation, the migrated cells on the
bottom surface were stained with 0.1% crystal violet (Santa Cruz
Biotechnology, Inc.) and counted under an Olympus LCX100 Imaging
system (Olympus Corporation, Tokyo, Japan).
Wound healing migration assay
HUVECs were grown to confluence in 24-well plates
and starved for 2 h. The media were changed to basic endothelial
growth basal medium (EBM-2), supplemented with 100 ng/ml VEGF, and
a scratch (wound) was made across the monolayer using a sterile
pipette tip. DHA was added to the culture medium with a final
concentration of 20 μM. Images of the wells were captured at fixed
points to record the area of clearing at time 0 and 8 h, and ImageJ
software (NIH, Bethesda, MD, USA) was used to quantitate the
cleared area.
Western blot analysis
HUVECs treated with 20μM DHA were collected at
different time points. To generate a positive control for p38 MAPK
activation, a group of HUVECs were treated with 1 μg/ml anisomycin
for 1 h. Cell lysates were prepared in radioimmunoprecipitation
assay buffer [20 mM Tris (pH 7.5), 150 mM NaCl, 50 mM NaF, 1%
NP-40, 0.1% deoxycholate, 0.1% SDS and 1 mM EDTA] (Santa Cruz
Biotechnology, Inc.), supplemented with 1 mM phenylmethylsulfonyl
fluoride and 1 μg/ml leupeptin (Santa Cruz Biotechnology, Inc.).
Cleared cell lysates were subjected to SDS-PAGE using 10%
polyacrylamide gel and transferred to polyvinylidene fluoride
membranes. Membranes were blocked with 2.5% non-fat milk, and
incubated with primary antibodies at 4°C overnight in
phosphate-buffered saline Tween-20 (Santa Cruz Biotechnology,
Inc.). The primary antibodies used were total-p38, phospho-p38
(Cell Signaling Technology, Inc.) and β-actin (Sigma-Aldrich, St.
Louis, MO, USA). Immunoreactivity was visualized with horseradish
peroxidase-conjugated secondary antibodies and an enhanced
chemiluminescence reagent (Santa Cruz Biotechnology, Inc.). The
blots were analyzed using a Bio-Rad imaging system (Bio-Rad,
Hercules, CA, USA).
ECIS migration analysis
Real-time EC migration was measured using the ECIS
technique (ECIS model 1600; Applied BioPhysics, Troy, NY, USA).
Briefly, eight-well ECIS arrays (8W10E+) were coated with
fibronectin (Invitrogen Life Technologies, Carlsbad, CA, USA).
HUVECs were plated at a confluent density to form monolayers
directly on top of the electrodes. Next, an elevated voltage pulse
of 40 kHz was applied to the arrays for 30 sec, which resulted in
the death and detachment of cells from the electrodes. As a result,
the wound was healed by the surrounding cells. Following treatment
with DHA or SB203850, an alternating current was applied to the
cells across the electrodes and the electrical resistance was
recorded. Data plots are representative of triplicate experiments,
with each graph showing the resistance readings from a separate
well, at 40 distinct electrodes per well.
Statistical analysis
Statistically significant differences were assessed
using a paired-samples t-test. All statistical analyses were
performed using SPSS 19.0 statistical software (Chicago, IL, USA)
with a significance level of P<0.05.
Results
DHA inhibits EC migration
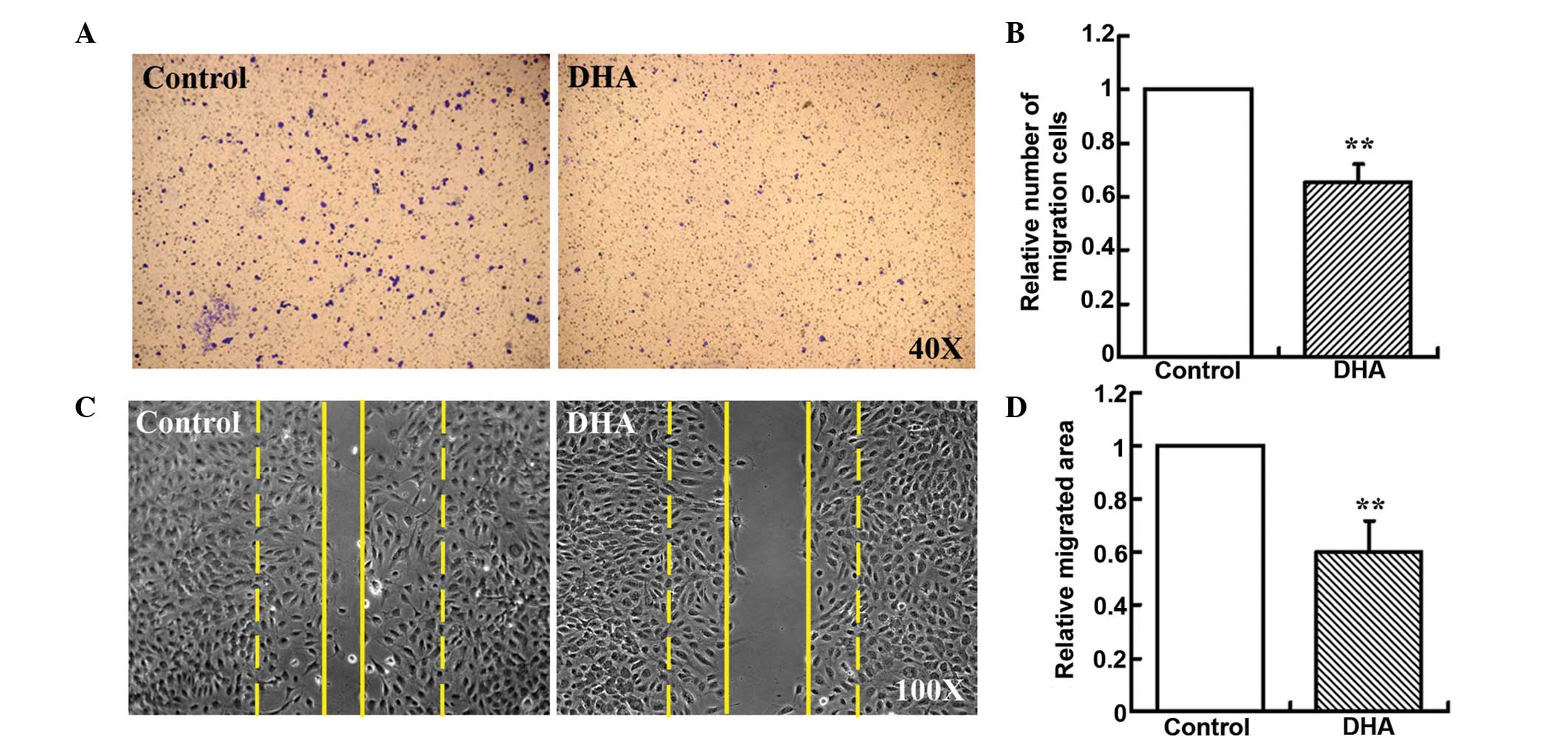
Boyden chamber-type cell migration assays were used
to assess the effect of DHA on EC migration. A low concentration of
20 μM DHA was used, since it had been demonstrated to be sufficient
to inhibit angiogenesis in vitro (14). The number of HUVECs migrating
across the polycarbonate membrane was significantly reduced in the
groups treated with 20 μM DHA (33.76%, P<0.01; Fig. 1A and B). Cell migration during
in vitro wound healing mimics the process of EC motility
in vivo (12,15). Thus, wound healing migration assays
were also performed, and the migrated area of HUVECs was
significantly reduced in the DHA-treated groups when compared with
the groups treated with vehicle DMSO alone (40.97%, P<0.01;
Fig. 1C and D). Thus, these in
vitro assays indicated that DHA induced the reduction of EC
migration.
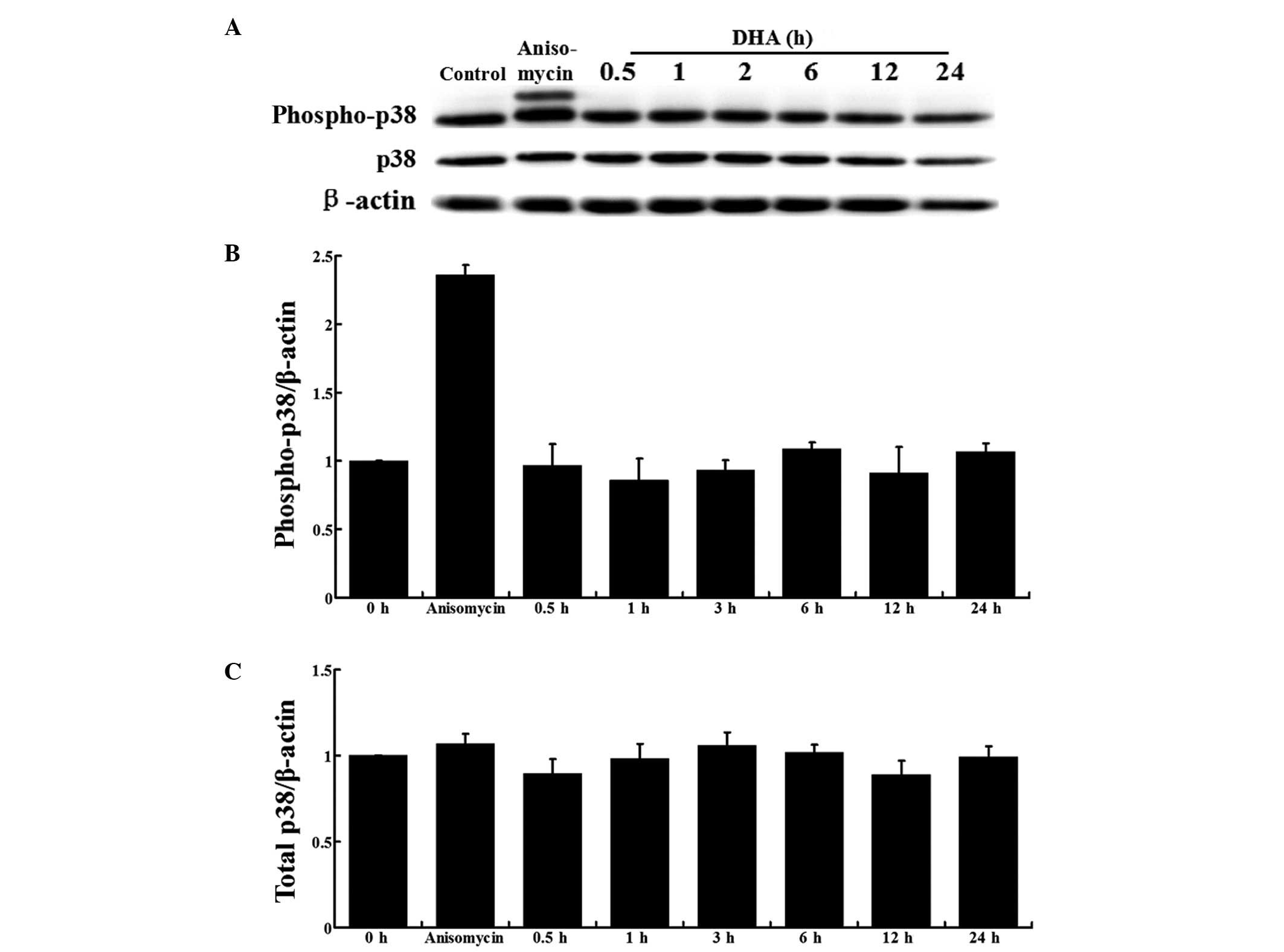
Activation of p38 MAPK is unaffected by
DHA in ECs
In ECs, p38 MAPK activation by VEGF mediates actin
reorganization and cell migration (12). To examine the effects of DHA on p38
MAPK activation, HUVECs cultured in EBM-2, containing 100 ng/ml
VEGF, were treated with 20 μM DHA and the protein expression was
analyzed at different time points. HUVECs treated with anisomycin,
a known activator of p38 MAPK, were used as a positive control
(16). Western blot analysis
showed that the total p38 and phospho-p38 MAPK protein expression
levels remained unchanged during DHA treatment at all the indicated
time points (Fig. 2A).
Densitometric analysis further confirmed that DHA did not affect
p38 MAPK activation or expression (Fig. 2B and C).
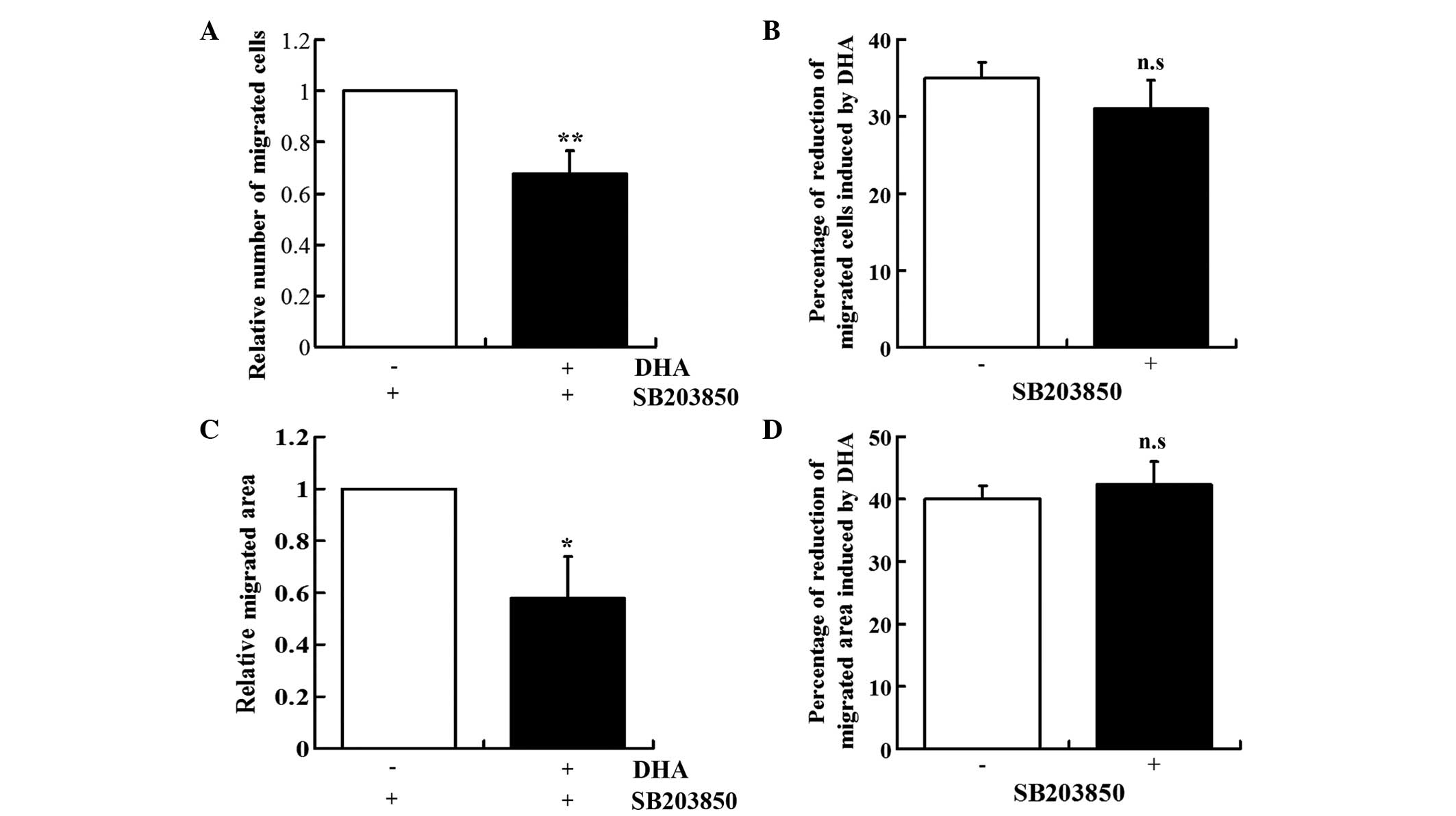
DHA-induced repression of EC migration is
unaffected by SB203850
SB203850 is a pyridinyl imidazole derivative that
inhibits p38 MAPK phosphorylation, but does not reduce p38 MAPK
expression (17). The role of p38
MAPK in mediating the DHA-induced inhibition of EC migration was
ascertained by examining the effect of SB203580 on the DHA-treated
HUVECs. In the presence of SB203580, DHA also induced the reduction
of EC migration in the Boyden chamber assay (30.38%, P<0.01) and
wound healing assay (43.04%, P<0.05; Fig. 3A and C). The levels of reduction
induced by DHA in the two assays were similar in the absence or
presence of SB203850 (Fig. 3B and
D).
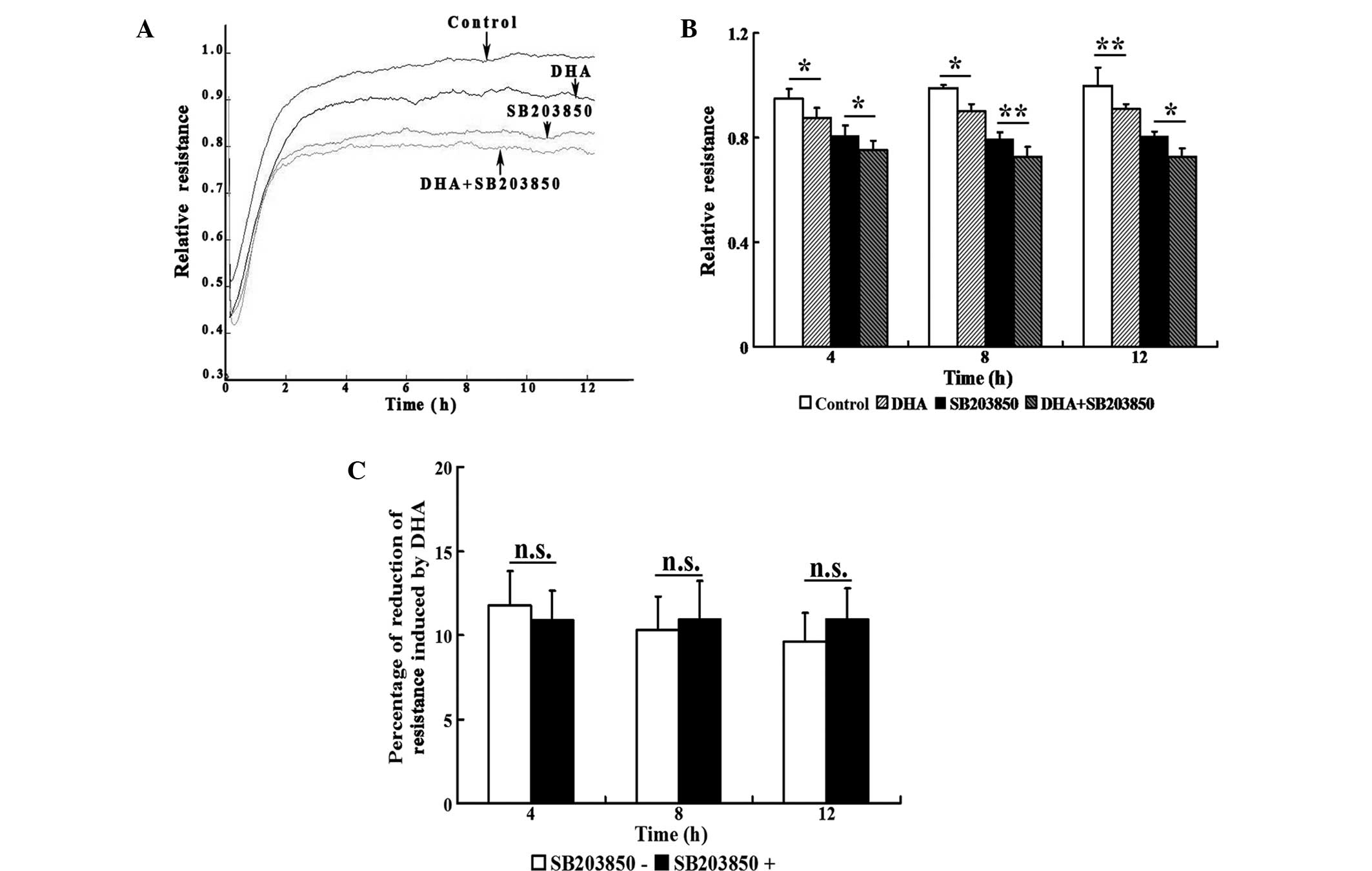
To capture the subtle alterations of cell migration,
an ECIS system was implemented, which allowed for real-time
measurements of the resistance caused by the migration of ECs. The
transendothelial resistance was significantly reduced when HUVECs
were treated with 20 μM DHA (P<0.05; Fig. 4A and B). Treatment with 20 μM
SB203580 also significantly decreased the transendothelial
resistance (P<0.05); however, treatment with DHA and SB203850
induced an additional reduction in transendothelial resistance at
4, 8 and 12 h (Fig. 4A and B). In
the absence or presence of SB203850, the reduction of
transendothelial resistance induced by DHA showed no statistically
significant difference at any of the time points (Fig. 4C). Therefore, the inhibitory
effects of DHA on EC migration are not mediated by the p38 MAPK
pathway.
Discussion
Antiangiogenic therapy targeting ECs to block
neovascularization has become an effective anticancer strategy
(18). DHA, a widely used
antimalarial drug with minimal side-effects, has been reported to
be highly effective against tumor angiogenesis (14). The aim of the present study was to
investigate the mechanisms underlying the effect of DHA on EC
migration, a key process in angiogenesis. DHA was shown to inhibit
EC migration in three types of migration assays. In addition, the
role of the p38 MAPK signaling pathway in the effect of DHA was
further investigated, and p38 MAPK activation was shown to not be
affected by DHA. Furthermore, the p38 MAPK inhibitor, SB203850, did
not inhibit or augment the DHA-induced reduction of EC migration.
To the best of our knowledge, the present study is the first to
investigate the signaling transduction pathways mediating the
effects of the artemisinin family of drugs on EC migration.
EC migration is a dynamic and multistep process that
plays a crucial role in the initiation and progression of
angiogenesis (19). Previous
studies have demonstrated the antiangiogenetic effects of DHA and
other artemisinin derivatives (20,21).
DHA has been shown to inhibit HUVEC migration in a dose-dependent
manner (14), and also to reduce
the migration of murine lymphatic ECs (22). In the current study, HUVEC
migration was examined through Boyden transwell assays, wound
healing assays and ECIS analysis. Results from all three methods
confirmed the inhibitory role of DHA on EC migration, which may
significantly contribute towards the antiangiogenetic effects.
Initiation of cell migration in response to
cytokines, including VEGF, is a highly controlled process requiring
the co-operation of signaling pathways, involving the extracellular
matrix, transmembrane receptors, such as integrins, and actin
cytoskeleton-associated motile apparatus (23,24).
MAPK pathways are important components of the signaling network,
transducing the migratory signals generated by VEGF and playing an
important role in the regulation of angiogenesis (12). VEGF activates the p38 MAPK pathway
via KDR/Flk-1 in ECs, while SB203580 induces the inhibition of p38
activity and cell migration (12).
Artemisinin and its derivatives have been shown to increase or
suppress p38 MAPK activity in different experimental settings
(25,26). In the present study, DHA was shown
to not affect the VEGF-induced activation of p38 MAPK, indicating
that DHA exerts antimigratory effects independent of the p38 MAPK
pathway.
The ECIS system provides a highly sensitive method
to monitor real-time cell migration (27). Consistent with previous
observations, ECIS analysis revealed that SB203850 significantly
inhibited cell migration. However, the application of DHA to
SB203850-treated HUVECs resulted in an additional decrease in cell
migration. Thus, inhibiting the p38 MAPK pathway failed to prevent
the DHA-induced reduction of cell migration. These results further
confirm that p38 MAPK does not mediate the DHA-induced inhibition
of EC migration.
Although p38 MAPK plays a key role in promoting cell
migration, other signaling pathways are also involved in regulating
this process. The nuclear factor-κB (NF-κB) family comprises a wide
range of transcription factors that are involved in numerous
cellular functions (28).
Activation of the NF-κB pathway promotes cell migration.
Extracellular stimuli, such as high glucose concentrations, inhibit
EC migration via the suppression of the NF-κB pathway (29). DHA inhibits the NF-κB pathway by
preventing the nuclear translocation of the p65/p50 complex
(30). In addition, the protein
kinase B (PKB)signaling pathway positively regulates cell
migration, and oxidized low-density lipoprotein inhibits EC
migration through the suppression of this pathway (31). Artemisinin and its derivatives have
been shown to inhibit PKB activity (32). Therefore, the DHA-induced
inhibition of EC migration may be mediated by these p38-independent
signaling pathways.
In conclusion, the results of the present study
demonstrated that DHA inhibits EC migration, which is independent
of p38 MAPK signaling. These observations provide an insight into
the mechanisms underlying the antiangiogenetic effects of the
artemisinin family of drugs, which may develop into therapeutic
agents against cancer and other vascular-associated diseases.
Acknowledgements
This study was supported by a grant from the Medical
Science and Technology Development Plan of Shandong Province, China
(no. 2013WS0137). The authors are grateful for the financial
support from the Shandong Taishan Scholarship (Ju Liu).
References
|
1
|
Klayman DL: Qinghaosu (artemisinin): an
antimalarial drug from China. Science. 228:1049–1055. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tu Y: The development of new antimalarial
drugs: qinghaosu and dihydro-qinghaosu. Chin Med J (Engl).
112:976–977. 1999.PubMed/NCBI
|
|
3
|
Gordi T and Lepist EI: Artemisinin
derivatives: toxic for laboratory animals, safe for humans? Toxicol
Lett. 147:99–107. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crespo-Ortiz MP and Wei MQ: Antitumor
activity of artemisinin and its derivatives: from a well-known
antimalarial agent to a potential anticancer drug. J Biomed
Biotechnol. 2012:2475972012.PubMed/NCBI
|
|
5
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Costa C, Soares R and Schmitt F:
Angiogenesis: now and then. APMIS. 112:402–412. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klagsbrun M and Moses MA: Molecular
angiogenesis. Chem Biol. 6:R217–R224. 1999. View Article : Google Scholar
|
|
8
|
Steeg PS: Tumor metastasis: mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hoefen RJ and Berk BC: The role of MAP
kinases in endothelial activation. Vascul Pharmacol. 38:271–273.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Page C and Doubell AF: Mitogen-activated
protein kinase (MAPK) in cardiac tissues. Mol Cell Biochem.
157:49–57. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rousseau S, Houle F, Landry J and Huot J:
p38 MAP kinase activation by vascular endothelial growth factor
mediates actin reorganization and cell migration in human
endothelial cells. Oncogene. 15:2169–2177. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harris VK, Coticchia CM, Kagan BL, Ahmad
S, Wellstein A and Riegel AT: Induction of the angiogenic modulator
fibroblast growth factor-binding protein by epidermal growth factor
is mediated through both MEK/ERK and p38 signal transduction
pathways. J Biol Chem. 275:10802–10811. 2000. View Article : Google Scholar
|
|
14
|
Wang SJ, Sun B, Cheng ZX, et al:
Dihydroartemisinin inhibits angiogenesis in pancreatic cancer by
targeting the NF-κB pathway. Cancer Chemother Pharmacol.
68:1421–1430. 2011.PubMed/NCBI
|
|
15
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: a convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu J and Kapron CM: Differential
induction of MAP kinase signalling pathways by cadmium in primary
cultures of mouse embryo limb bud cells. Reprod Toxicol.
29:286–291. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cuenda A, Rouse J, Doza YN, et al: SB
203580 is a specific inhibitor of a MAP kinase homologue which is
stimulated by cellular stresses and interleukin-1. FEBS Lett.
364:229–233. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vasudev NS and Reynolds AR:
Anti-angiogenic therapy for cancer: current progress, unresolved
questions and future directions. Angiogenesis. 17:471–494. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lamalice L, Le Boeuf F and Huot J:
Endothelial cell migration during angiogenesis. Circ Res.
100:782–794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang JL, Wang Z, Hu W, Chen SS, Lou XE
and Zhou HJ: DHA regulates angiogenesis and improves the efficiency
of CDDP for the treatment of lung carcinoma. Microvasc Res.
87:14–24. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng R, Li C, Li C, et al: The
artemisinin derivative artesunate inhibits corneal
neovascularization by inducing ROS-dependent apoptosis in vascular
endothelial cells. Invest Ophthalmol Vis Sci. 54:3400–3409. 2013.
View Article : Google Scholar
|
|
22
|
Wang J, Guo Y, Zhang BC, Chen ZT and Gao
JF: Induction of apoptosis and inhibition of cell migration and
tube-like formation by dihydroartemisinin in murine lymphatic
endothelial cells. Pharmacology. 80:207–218. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nobes CD and Hall A: Rho, rac, and cdc42
GTPases regulate the assembly of multimolecular focal complexes
associated with actin stress fibers, lamellipodia, and filopodia.
Cell. 81:53–62. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lauffenburger DA and Horwitz AF: Cell
migration: a physically integrated molecular process. Cell.
84:359–369. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang JX, Hou LF, Yang Y, Tang W, Li Y and
Zuo JP: SM905, an artemisinin derivative, inhibited NO and
pro-inflammatory cytokine production by suppressing MAPK and
NF-kappaB pathways in RAW 264.7 macrophages. Acta Pharmacol Sin.
30:1428–1435. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu JJ, Meng LH, Cai YJ, et al:
Dihydroartemisinin induces apoptosis in HL-60 leukemia cells
dependent of iron and p38 mitogen-activated protein kinase
activation but independent of reactive oxygen species. Cancer Biol
Ther. 7:1017–1023. 2008. View Article : Google Scholar
|
|
27
|
Hong J, Kandasamy K, Marimuthu M, Choi CS
and Kim S: Electrical cell-substrate impedance sensing as a
non-invasive tool for cancer cell study. Analyst. 136:237–245.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jacobs MD and Harrison SC: Structure of an
IkappaBalpha/NF-kappaB complex. Cell. 95:749–758. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hamuro M, Polan J, Natarajan M and Mohan
S: High glucose induced nuclear factor kappa B mediated inhibition
of endothelial cell migration. Atherosclerosis. 162:277–287. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hwang YP, Yun HJ, Kim HG, Han EH, Lee GW
and Jeong HG: Suppression of PMA-induced tumor cell invasion by
dihydroartemisinin via inhibition of PKCalpha/Raf/MAPKs and
NF-kappaB/AP-1-dependent mechanisms. Biochem Pharmacol.
79:1714–1726. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chavakis E, Dernbach E, Hermann C, Mondorf
UF, Zeiher AM and Dimmeler S: Oxidized LDL inhibits vascular
endothelial growth factor-induced endothelial cell migration by an
inhibitory effect on the Akt/endothelial nitric oxide synthase
pathway. Circulation. 103:2102–2107. 2001. View Article : Google Scholar
|
|
32
|
Cheng C, Ho WE, Goh FY, et al:
Anti-malarial drug artesunate attenuates experimental allergic
asthma via inhibition of the phosphoinositide 3-kinase/Akt pathway.
PloS One. 6:e209322011. View Article : Google Scholar : PubMed/NCBI
|