Introduction
Autophagy exists in higher eukaryotes ranging from
yeast to humans. The process is highly conserved and maintains
intracellular stability (1).
Distinct from the ubiquitin-proteasome pathway, which degrades
short-life proteins, autophagy degrades long-life bioplasmin and
malfunctional organelles. There a five steps in the autophagy
process (2–4): i) Activation of a dual-membrane
structure, known as a phagocytic vacuole; ii) amplification of the
phagocytic vacuole; iii) maturation of the phagosome; iv)
combination between phagosome and lysosome; v) autophagy. In normal
physiological conditions, autophagy is maintained at a low level
and preserves normal physiological function by cleaning up injured
organelles and biomolecules, such as mitochondria and proteins.
Dysfunctional autophagy can lead to cancer and degenerative
disease; however, whether autophagy participates in the
pathogenesis of kidney disease remains unclear. Considerable
evidence shows a close association between autophagy and a number
of kidney diseases, including acute kidney injury (AKI), diabetic
nephropathy and polycystic kidney disease (5). This association could be utilized in
the development of novel treatments for kidney diseases.
Erythropoietin (EPO) is a hormone that regulates the
generation of erythrocytes. It is primarily produced by the renal
cortex and peritubular fibroblasts. The levels of renal, hepatic
and cerebral endogenous EPO increase when blood supply or oxygen
insufficiency occurs (6). EPO
assists oxygen delivery and erythropoiesis and reduces apoptosis,
oxidative stress and inflammation (7,8).
Several studies have attempted to show the effects of EPO on the
kidney, but the mechanism has remained unclear (Jackevicius,
Moeini).
The precursor LC3 is divided and formed into LC3-I
by autophagy-related gene 4 (Atg4). LC3-I is transformed into the
membrane-bound form LC3-II through the activation of Atg7, which is
located on the membrane of autophagosomes and autolysosomes
(9). LC3-I and LC3-II are
indicators of autophagy, and show the activation of autophagy
(10). p62/sequestosome-1 (SQSTM1)
is an ubiquitin-binding protein, and can induce the autophagy of
ubiquitinated proteins by lysosomes in mammalian cells, through
cooperation with LC3 (11). The
high expression of p62/SQSTM1 can cause an upregulation of the
autophagy process and can act as an indicator of autophagy
(12).
The aim of the present study was to induce autophagy
in rat renal glomerular mesangial cells (GMCs) through the use of
lipopolysaccharide (LPS) and to explore the effect of EPO on GMC
autophagy to provide novel insight into EPO-mediated renal
protection.
Materials and methods
Isolation and culture of rat renal
GMCs
A total of 32 two-month-old male Sprague Dawley rats
were supplied by the Henan Laboratory Animal Center (Zhengzhou,
China). This study was carried out in strict accordance with the
recommendations in the Guide for the Care and Use of Laboratory
Animals of the National Institutes of Health. The animal use
protocol was reviewed and approved by the Institutional Animal Care
and Use Committee of the First Affiliated Hospital of Xinxiang
Medical University (Weihui, China). Both kidneys were isolated from
the two-month-old Sprague Dawley rats aseptically. The renal
capsule was exfoliated, minced into small pieces and placed on a
three-layered stainless steel screen, prior to being washed and
ground with a pestle simultaneously. The washing process was
terminated when kidney tubules could no longer be observed under
the microscope and when >98% of the glomeruli were not
associated with the Bowman’s capsule. The glomerular tissue was
then collected from the second screen (<75 μm) into a centrifuge
tube for centrifugation at 15,000 × g for 5 min. The supernatant
was subsequently discarded and the sediment was treated with
collagenase V (Sigma, St. Louis, MO, USA) for 10–15 min at 37°C.
The reaction was terminated with the addition of RPMI-1640
medium/15% fetal bovine serum (FBS) (Gibco-BRL, Grand Island, NY,
USA). The sample was then centrifuged at 15,000 × g for a further 5
min prior to the supernatant being discarded. The glomeruli were
collected and seeded in a gelatinized culture flask with RPMI-1640
medium/15% FBS at 37°C and 5% CO2. The first passage was
conducted after 7–10 days. Twenty-four hours after passage, the
cells adhered and formed fusiform or irregular stellate cells.
After 3–4 days of culture, the cells formed a tablet.
Immunofluorescent staining
Immunofluorescent staining was performed by
initially activating the cells in vitro, staining the
surface for antigens and fixing with paraformaldehyde in order to
stabilize the cell membrane. The cell membrane was then
permeabilized with the detergent saponin in order to allow the
antibodies to stain intracellularly. Immunofluorescence showed
positive results for monoclonal mouse anti-human α-smooth muscle
actin (1:200 dilution; Beyotime Institute of Biotechnology,
Shanghai, China) and rabbit anti-human monoclonal vimentin (1:200
dilution; Beyotime Institute of Biotechnology), and negative
results for mouse monoclonal cytokeratin (1:200 dilution; Beyotime
Institute of Biotechnology) and mouse monoclonal factor VIII (1:200
dilution; Beyotime Institute of Biotechnology). Cells cultured over
three passages were used for examination.
Grouping
The cells were divided into four groups as follows:
i) Control (cells were cultured in RPMI-1640 medium/15% FBS and 5
mmol/l glucose; ii) high-glucose (cells were cultured in RPMI-1640
medium/15% FBS and 30 mmol/l glucose; iii) LPS (cells were cultured
in RPMI-1640 medium/15% FBS and 10 nmol/ml LPS (Sigma); iv) LPS +
EPO (cells were cultured in RPMI-1640 medium/15% FBS, 10 ng/ml LPS
and 10 nmol/ml EPO (Sigma). Cells were harvested for examination
after 24- and 72-h culture periods.
Western blot analysis
A total of 0.1 ml pre-chilled
radioimmunoprecipitation assay buffer [50 mmol/l Tris-Cl (pH 7.6),
150 mmol/l NaCl, 1% NP-40, 0.1% sodium dodecyl sulfate (SDS), 0.5%
deoxycholate, 1 μl/ml leupeptin, 1 μl/ml aprotinin and 0.5 mmol/l
phenylmethylsulfonyl fluoride] (Beyotime Institute of
Biotechnology) was added to the homogenate and chilled on ice for
30 min. The sample was then centrifuged at 15,000 × g for 30 min,
the supernatant was pipetted and the concentration of protein was
measured with a bicinchoninic acid protein assay kit (Baiwang
Biotechnology Corporation, Shenzhen, China). SDS loading buffer (10
ml) [2X; 2 ml 0.5 mol/l Tris (pH 6.8), 2 ml glycerin, 2 ml 20% SDS,
0.5 ml 0.1% bromophenol blue, 1 ml 1 mol/l dithiothreitol and 2.5
ml ddH2O] (TaKaRa, Dalian, China) was added to 30–50 μg
protein, and the sample was heated at 100°C for 5 min to denature
the protein. Following SDS-PAGE and protein transfer to a
nitrocellulose membrane, the membrane was blocked with 1X blocking
buffer [5% skimmed milk powder 0.5 g; 20 mM Tris-Cl, (pH 7.5–8.0);
150 mM NaCl; 0.05% Tween-20; Baiwang Biotechnology Corporation] at
room temperature for 1 h. The membrane was then immersed in the
primary antibody (LC3, dilution ratio 1:2,000; p62, dilution ratio
1:200) (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and
incubated overnight at 4°C. Following incubation with the
polyclonal primary antibody (catalogue no. F2426), the membrane was
rinsed three times with Tris-buffered saline and Tween®
20 (Baiwang Biotechnology Corporation), with each rinse lasting 5
min. The membrane was then immersed in mouse anti-rabbit secondary
antibody (dilution ratio 1:2,000) and incubated at room temperature
for 1 h, prior to being rinsed a further three times. A
chemiluminescent graph was used to be colored and detected. All of
the experiments were performed according to the manufacturer’s
instructions (Baiwang Biotechnology Corporation). β-actin was used
as an internal control.
Statistical analysis
All data are expressed as the mean ± standard
deviation. Comparisons between two groups were conducted with an
independent samples t-test. SPSS 17.0 (SPSS, Inc., Chicago, IL,
USA) was utilized for the analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
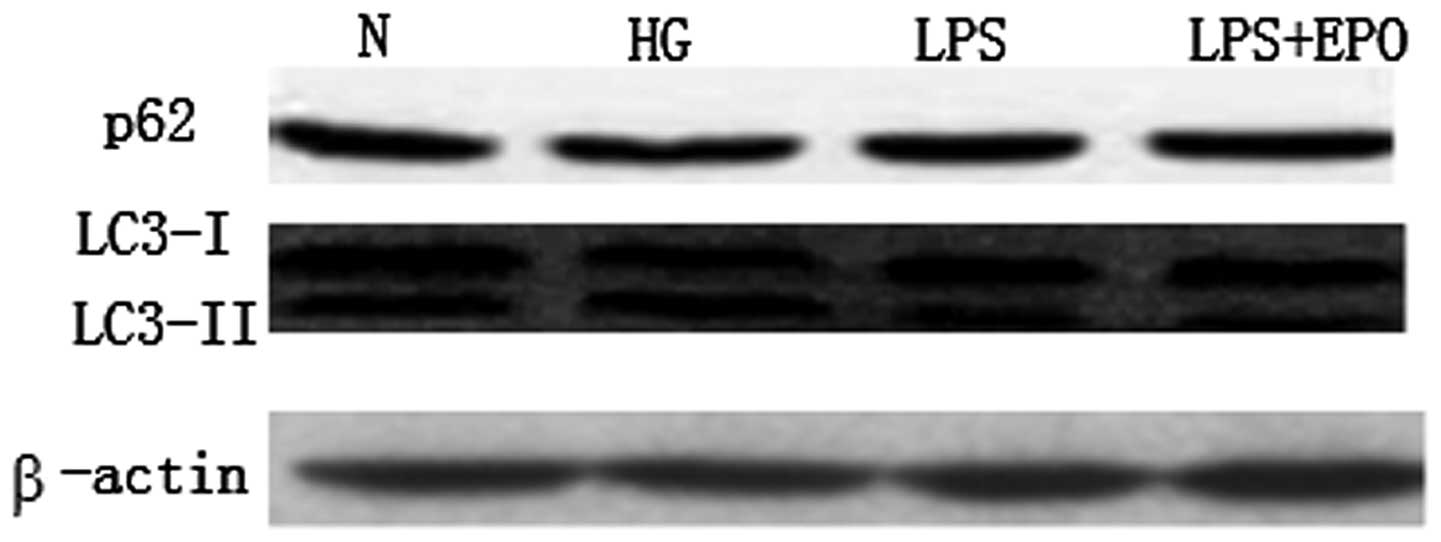
In the present study, no significant differences in
the protein expression of LC3-I and II, and p62/SQSTM1 were found
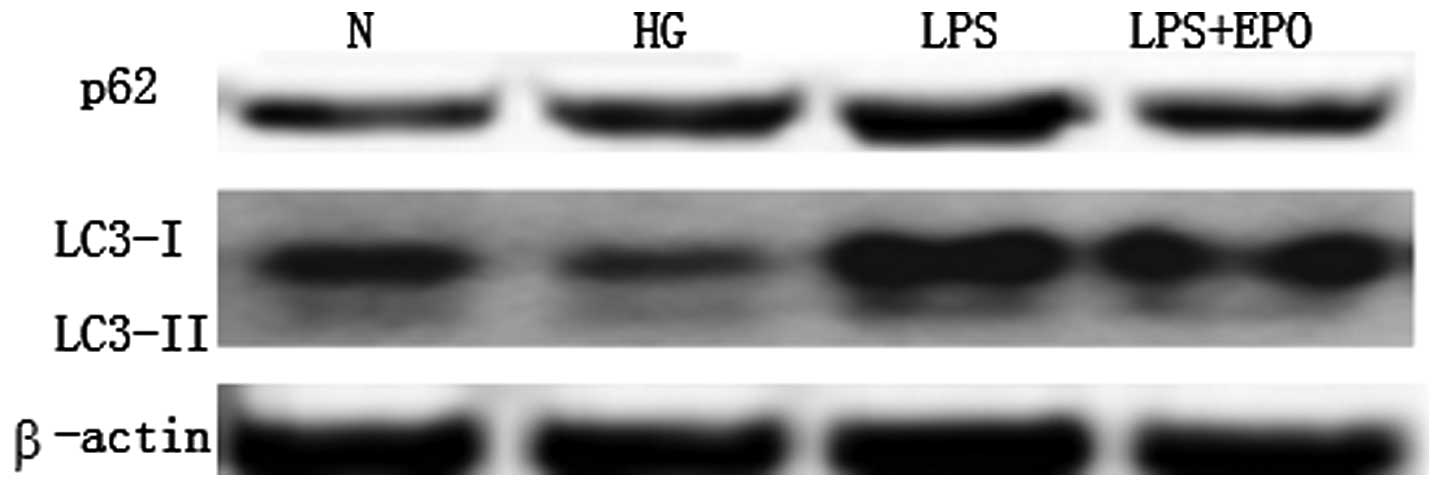
among the four groups in the 24-h culture experiment (Fig. 1). After 72 h of culture, the
expression of p62/SQSTM1 was increased, and that of LC3-I and II
was decreased in the high-glucose group compared with the
expression in the control group (Fig.
2). By contrast, the expression of p62/SQSTM1 was increased and
that of LC3-I and II was increased in the LPS group. The changes
were reversed in the LPS + EPO group (Fig. 2). These results suggest that
high-glucose conditions can inhibit autophagy in mesangial cells;
LPS can induce autophagy in mesangial cells and EPO can suppress
this induction.
Discussion
Several factors, such as diabetes and bacterial
infection, can lead to a renal disease, which can seriously affect
human health. Autophagy is a common defense mechanism that exists
widely in eukaryotes. Autophagy can maintain the intracellular
stability by cleaning up damaged or aged organelles and
biomolecules, such as mitochondria, peroxisomes, endoplasmic
reticulum and paraprotein (13,14).
Autophagy therefore plays an important role in pathology. It has
been shown that autophagy participates in renal
ischemia/reperfusion injury. In a study by Li et al
(15) it was suggested that high
glucose concentrations could inhibit autophagy in mesangial cells,
while rapamycin could suppress the high glucose-induced autophagy
and N3-methyladenine could upregulate the high glucose-induced
autophagy in renal mesangial cells. Renal diseases usually cause
anemia, due to an EPO deficiency originating from the renal cortex
and renal tubule fibroblasts. As a result, EPO is often
administered in renal therapy. In a recent study, Yamaleyeva et
al (16) treated chronic renal
injury with EPO-positive cells and revealed promising results.
Furthermore, in a study by Oh et al (17) it was found that kidney function
could be conditioned using EPO following the occurrence of AKI
(17), mainly by improving oxygen
transport and erythropoiesis, as well as reducing autophagy,
oxidative stress and inflammation (7,8).
In the present study, it was found that high-glucose
conditions could suppress autophagy in GMCs; this finding was
consistent with the results in the study by Li et al
(15). The present results
suggested that LPS could induce autophagy in GMCs, and that EPO
could inhibit the LPS-induced autophagy and protect the renal cells
from damage. In an oxidative toxicity rat model, EPO was previously
shown to block the autophagy signals in the brain to keep the brain
from damage (18). In a recent
study by Yu et al (19), it
was reported that EPO could protect endothelial cells through the
same mechanism in a neonatal necrotizing enterocolitis rat model.
This indicated that inflammation could induce autophagy and
apoptosis and that EPO could block this pathway to protect the
cells. The inhibition of autophagy has certain value in clinical
practice. A previous study by Zhang et al (20) showed that the treatment of
folliculin-deficient renal cancer was enhanced through the use of a
combination paclitaxel and autophagy inhibitor treatment. Both
autophagy inhibition and efforts to target the B-cell lymphoma-2
family of proteins are strategies that could overcome drug
tolerance (21). By contrast,
Kimura et al (22) reported
that mitochondrial metabolic stress in renal proximal tubular
epithelial cells can lead to a defensive autophagy. Furthermore,
the study by Yamahara et al (23) showed that obesity-mediated
autophagy cannot exacerbate proteinuria-induced tubulointerstitial
injury. The role of autophagy in renal disease therefore remains to
be fully elucidated.
In conclusion, the present study showed that
high-glucose conditions can inhibit autophagy in renal cells, and
that LPS can induce autophagy. Furthermore, EPO can reverse this
LPS-induced autophagy and protect renal mesangial cells from
damage.
References
|
1
|
Levine B and Klionsky DJ: Development by
self-digestion: molecular mechanisms and biological functions of
autophagy. Dev Cell. 6:463–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang Z and Klionsky DJ: Eaten alive: a
history of macroautophagy. Nat Cell Biol. 12:814–822. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weide T and Huber TB: Implications of
autophagy for glomerular aging and disease. Cell Tissue Res.
343:467–473. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mizushima N and Levine B: Autophagy in
mammalian development and differentiation. Nat Cell Biol.
12:823–830. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huber TB, Edelstein CL, Hartleben B, et
al: Emerging role of autophagy in kidney function, diseases and
aging. Autophagy. 8:1009–1031. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weidemann A and Johnson RS: Nonrenal
regulation of EPO synthesis. Kidney Int. 75:682–688. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Toba H, Nakashima K, Oshima Y, et al:
Erythropoietin prevents vascular inflammation and oxidative stress
in subtotal nephrectomized rat aorta beyond haematopoiesis. Clin
Exp Pharmacol Physiol. 37:1139–1146. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Malgorzewicz S, Lichodziejewska-Niemierko
M, Lizakowski S, et al: Oxidative stress, inflammation and
nutritional status during darbepoetin alpha treatment in peritoneal
dialysis patients. Clin Nephrol. 73:210–215. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kadowaki M and Karim MR: Cytosolic LC3
ratio as a quantitative index of macroautophagy. Methods Enzymol.
452:199–213. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pankiv S, Clausen TH, Lamark T, et al:
p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of
ubiquitinated protein aggregates by autophagy. J Biol Chem.
282:24131–24145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mizushima N and Yoshimori T: How to
interpret LC3 immunoblotting. Autophagy. 3:542–545. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kondo-Okamoto N, Noda NN, Suzuki SW, et
al: Autophagy-related protein 32 acts as an autophagic degron and
directly initiates mitophagy. J Biol Chem. 287:10631–10638. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xie Z and Klionsky DJ: Autophagosome
formation: core machinery and adaptations. Nat Cell Biol.
9:1102–1109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Bai XY, Cui SY, Fu B and Chen X:
Effect of rapamycin on high glucose-induced autophagy impairment,
oxidative stress and premature senescence in rat mesangial cells in
vitro. Nan Fang Yi Ke Da Xue Xue Bao. 32:467–471. 2012.(In
Chinese). PubMed/NCBI
|
|
16
|
Yamaleyeva LM, Guimaraes-Souza NK, Krane
LS, et al: Cell therapy with human renal cell cultures containing
erythropoietin-positive cells improves chronic kidney injury. Stem
Cells Transl Med. 1:373–383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oh SW, Chin HJ, Chae DW and Na KY:
Erythropoietin improves long-term outcomes in patients with acute
kidney injury after coronary artery bypass grafting. J Korean Med
Sci. 27:506–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bendix I, Schulze C, Haefen Cv, et al:
Erythropoietin modulates autophagy signaling in the developing rat
brain in an in vivo model of oxygen-toxicity. Int J Mol Sci.
13:12939–12951. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu Y, Shiou SR, Guo Y, et al:
Erythropoietin protects epithelial cells from excessive autophagy
and apoptosis in experimental neonatal necrotizing enterocolitis.
PLoS One. 8:e696202013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Q, Si S, Schoen S, et al:
Suppression of autophagy enhances preferential toxicity of
paclitaxel to folliculin-deficient renal cancer cells. J Exp Clin
Cancer Res. 32:992013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mei H, Lin Z, Wang Y, Wu G and Song Y:
Autophagy inhibition enhances pan-Bcl-2 inhibitor AT-101-induced
apoptosis in non-small cell lung cancer. Neoplasma. 61:186–192.
2014. View Article : Google Scholar
|
|
22
|
Kimura T, Takahashi A, Takabatake Y, et
al: Autophagy protects kidney proximal tubule epithelial cells from
mitochondrial metabolic stress. Autophagy. 9:1876–1886. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamahara K1, Kume S, Koya D, et al:
Obesity-mediated autophagy insufficiency exacerbates
proteinuria-induced tubulointerstitial lesions. J Am Soc Nephrol.
24:1769–1781. 2013. View Article : Google Scholar : PubMed/NCBI
|