Introduction
The most prominent feature of chronic kidney disease
(CKD) is renal interstitial fibrosis (RIF). The pathological
changes associated with RIF include cellular changes, changes in
the extra cellular matrix (ECM) and changes in growth factor
interactions (1).
Studies have revealed that renal ischemia and
hypoxia are caused by microvessel loss during CKD. Adenosine (ADO)
is an important signaling molecule that is induced under ischemic
and hypoxic conditions (2,3) and which plays a specific biological
role following adenosine receptor (AR) binding on the cell surface.
In renal tissues, ADO reduces renal blood flow by constricting
afferent arterioles, thereby reducing renal transport load
(4–8) and protecting short-term renal
function. A previous study observed that adenosine deaminase
(ADA)-deficient mice with prolonged exposure to high concentrations
of ADO developed RIF lesions and renal damage, indicating that ADO
plays an important role in promoting RIF damage (9).
Based on these facts, the present study considered
the hypothesis that ADO may be an important signaling molecule in
the development of RIF. It was hypothesized that under long-term
ischemic and hypoxic conditions, intracellular proinflammatory
cytokine release is induced by increased ADO concentrations in the
renal tissues. Fibroblasts and collagen deposition are activated
and regulated, which ultimately results in RIF and leads to kidney
damage. Therefore, the present study selected the classic mouse
unilateral ureteral obstruction (UUO) model mechanism of RIF.
Following modeling, hypoxia of the renal tissue, ADO concentration
changes and pathological changes in the obstructed renal tissues
were observed at different time points in order to assess the
degree of RIF. Changes in the deposition of related profibrogenic
factors and interstitial collagen were detected through regulation
of the ADO signaling pathway in order to investigate the
association between ADO and RIF.
Materials and methods
Animals
The present study was carried out in strict
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of the National Institutes of Health,
Second Edition (2011). The protocol was approved by the Committee
on the Ethics of Animal Experiments of the Third Xiangya Hospital
of Central South University (Changsha, China). All surgery was
performed under sodium pentobarbital anesthesia and all efforts
were made to minimize suffering. A total of 44 Kunming male mice
with an average weight of ~40 g were randomly divided into three
groups: Sham group (n=12), model group (UUO group; n=16) and
intervention group (PT group; n=16). The mice in the intervention
group were intraperitoneally injected with 10 mg/kg/day
8-(p-sulfophenyl)-theophylline (8-PT), a non-selective AR
blocker, once each day following UUO surgery (1 time/day) (10). The mice in the model group were
intraperitoneally injected with normal saline. On days 1, 3, 7, and
14 following each surgery, a number of mice were sacrificed,
specifically, four each from the UUO and PT groups and three from
the sham group. Prior to sacrifice, the mice were placed in
metabolic cages for 24 h to enable the collection of urine. A
solution of Hypoxyprobe™-1 (pimonidazole HCl; 60 mg/kg)
from a Hyproxyprobe-1 kit (Hypoxyprobe, Burlington, MA, USA) was
intravenously injected into the penile region of the mice in order
to observe the degree of tissue hypoxia 1.5 h prior to the mice
being sacrificed.
UUO model
Mice were intravenously injected with 10% sodium
pentobarbital (40 mg/kg). A 1.5-cm left upper quadrant midline
incision was made to locate the left ureter. The renal pelvis near
the ureter and the middle and upper junctions were ligated with no.
1 silk thread. The ureter between the two ligatures was cut. In the
sham group, the left ureter was left free without ligation and the
other steps were the same as for the UUO group.
Quantitative polymerase chain reaction
(qPCR)
The mouse cDNA sequence was obtained from GenBank.
Primers were designed using Primer Premier software, version 3.0
(Premier Biosoft, Palo Alto, CA, USA) and were synthesized by
ProMab (Richmond, CA, USA). Respective primer sequences are shown
in Table I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Genes | Upstream and
downstream primers (5′-3′) | Amplified fragment
length (bp) | Temperature (°C) |
|---|
| α1(I)
procollagen-F |
GTTCTCCTGGCAAAGACGGA | 199 | 58 |
| α1(I)
procollagen-R |
CGGCCACCATCTTGAGACTT | | |
|
TGF-β1-F |
AGGGCTACCATGCCAACTTC | 168 | 58 |
|
TGF-β1-R |
CCACGTAGTAGACGATGGGC | | |
| α-SMA-F |
GGACTCTGGAGATGGTGTGAC | 167 | 58 |
| α-SMA-R |
CAATCTCACGCTCGGCAGTA | | |
| GAPDH (mouse)-F |
AACTTTGGCATTGTGGAAGG | 132 | 58/59 |
| GAPDH (mouse)-R |
GGATGCAGGGATGATGTTCT | | |
The total RNA in the renal tissue was extracted
using TRIzol® reagent (15596-026; Invitrogen Life
Technologies, Carlsbad, CA, USA) and RiboLock™
ribonuclease inhibitor (EO0381; Thermo Fisher Scientific,
Pittsburgh, PA, USA) was used to eliminate genomic DNA. The reverse
transcription (RT) reaction was performed according to the
instructions of the RevertAid™ H Minus First Strand cDNA
Synthesis kit (K1631, Thermo Fisher Scientific). The mRNA
transcript levels of tumor growth factor β1
(TGF-β1) and α1(I) procollagen were
quantified using qPCR according to instructions provided with
SYBR® Green PCR Master Mix (4309155; Applied Biosystems,
Carlsbad, CA, USA).
Pathological analysis
The degree of tubular injury was scored by
hematoxylin and eosin (H&E) staining; the degree of RIF was
judged by Masson’s trichrome staining (11,12).
Renal tissue TGF-β1 and α-smooth muscle actin (α-SMA)
levels were detected at each time point by immunohistochemistry
using anti-TGF-β1 (1:50, Bioss, Ltd., Woburn, MA, USA)
and anti-α-SMA (1:200, Wuhan Boster Biological Technology, Ltd.,
Wuhan, China) antibodies. The degree of renal tissue hypoxia was
semi-quantitatively determined using a Hypoxyprobe-1 kit (13).
A single-blind pathological examination was
performed according to a multi-step procedure. Renal tissue samples
were collected from the mice, paraffin-embedded and sectioned into
5-μm slices. From each renal sample four slices were randomly
selected. Routine H&E and Masson’s trichrome staining was
performed and the morphology observed using a microscope. Each
slice was analyzed by the same individual with five non-overlapping
fields randomly selected in each slice. Positive cells or areas
were represented by an average optical density, separately
calculated in the selected four parts of the renal tissue sample in
each mouse. The average value of the four parts was calculated. The
average optical density for the positive areas was automatically
determined by Image-Pro Plus 6.0 software (Media Cybernetics,
Rockville, MD, USA).
High performance liquid chromatography
(HPLC) assay
Approximately 1/3 of the left kidney was cut
immediately upon being extracted from the mouse and preserved in a
liquid nitrogen tank to determine the ADO concentration. The HPLC
assay was performed on a reversed phase custom ocadecyl-silica
(ODS) column (4.6×254 mm) with a detection wavelength of 260 nm at
30°C. Following adenine nucleotide extraction, the ADO
concentration was designated the abscissa (x) and its corresponding
peak area as the vertical axis (y). The regression equation was
obtained; the ADO concentration was determined by conversion from
the corresponding peak areas of the test sample.
Statistical analyses
All data are expressed as the mean ± standard error
of the mean and were analyzed for statistical significance using
GraphPad Prism 5 software (GraphPad Software, San Diego, CA, USA).
Student’s t-tests were applied for two-group analysis. The
statistical significance of the differences in multiple groups of
mice was assessed by analysis of variance (ANOVA). The comparison
between two groups was performed using Tukey’s test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Continuous renal tissue hypoxia during
RIF with increased ADO levels
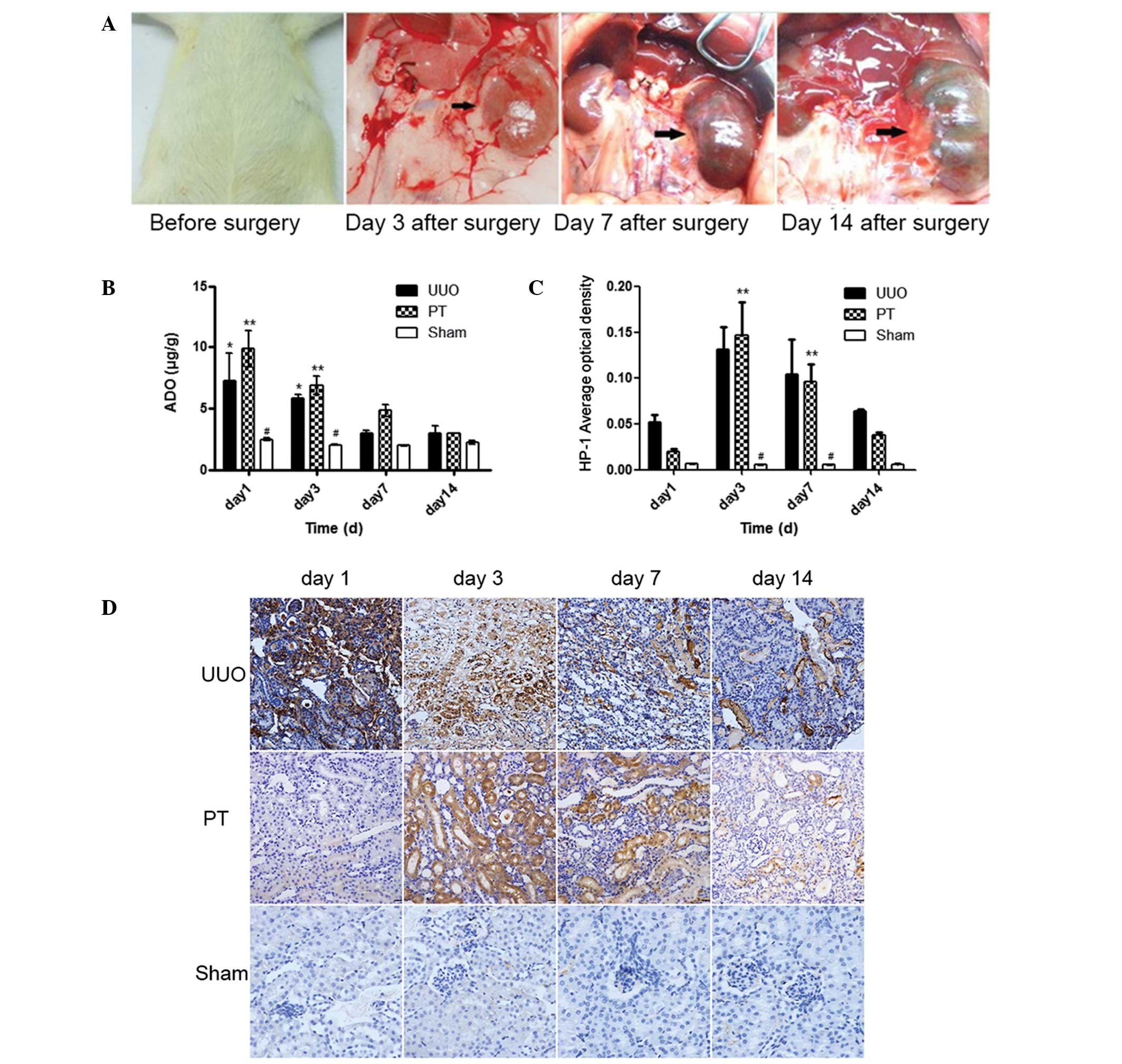
Morphological changes in the kidney were observed on
days 1, 3, 7 and 14 following modeling. As shown in Fig. 1A, hydronephrosis progressively
increased over time following the UUO procedure.
Hydronephrosis progressively increases as
the duration of UUO is extended
The ADO concentrations in the renal tissue were
determined through HPLC assays at different times following the UUO
procedure. In the UUO and PT groups, the ADO concentrations on days
1 and 3 after modeling were revealed to be significantly higher
than those in the sham group (P<0.05). In the PT group, the ADO
concentrations on days 1, 3 and 7 after modeling were higher than
those in the UUO group at the same time points. As the time
subsequent to the UUO procedure extended, the ADO concentrations of
the two groups converged on day 14 after modeling (Fig. 1B). These results indicated that
after the AR was blocked by PT, extracellular ADO binding to the AR
was reduced and metabolism slowed down, resulting in a progressive
increase in the extracellular ADO concentration. However, following
the UUO procedure, extracellular ADO concentration was decreased
due to increased apoptosis and ATP depletion.
The degree of renal tissue hypoxia was measured
following the intravenous injection of Hypoxyprobe-1 solution into
the mouse penile region. Changes in the degree of hypoxia were
observed following the UUO procedure. The degree of hypoxia in the
UUO and PT groups was significantly higher than that in the sham
group (P<0.05), particularly on days 3 and 7. On day 3, the
degree of hypoxia peaked and subsequently gradually declined. No
significant difference in the degree of hypoxia was identified
between the PT and UUO groups (Fig. 1C
and D). The brown-stained parts of the renal tissue represented
the degree of renal tissue hypoxia. On day 3, the degree of hypoxia
in the UUO and PT groups peaked and gradually declined
thereafter.
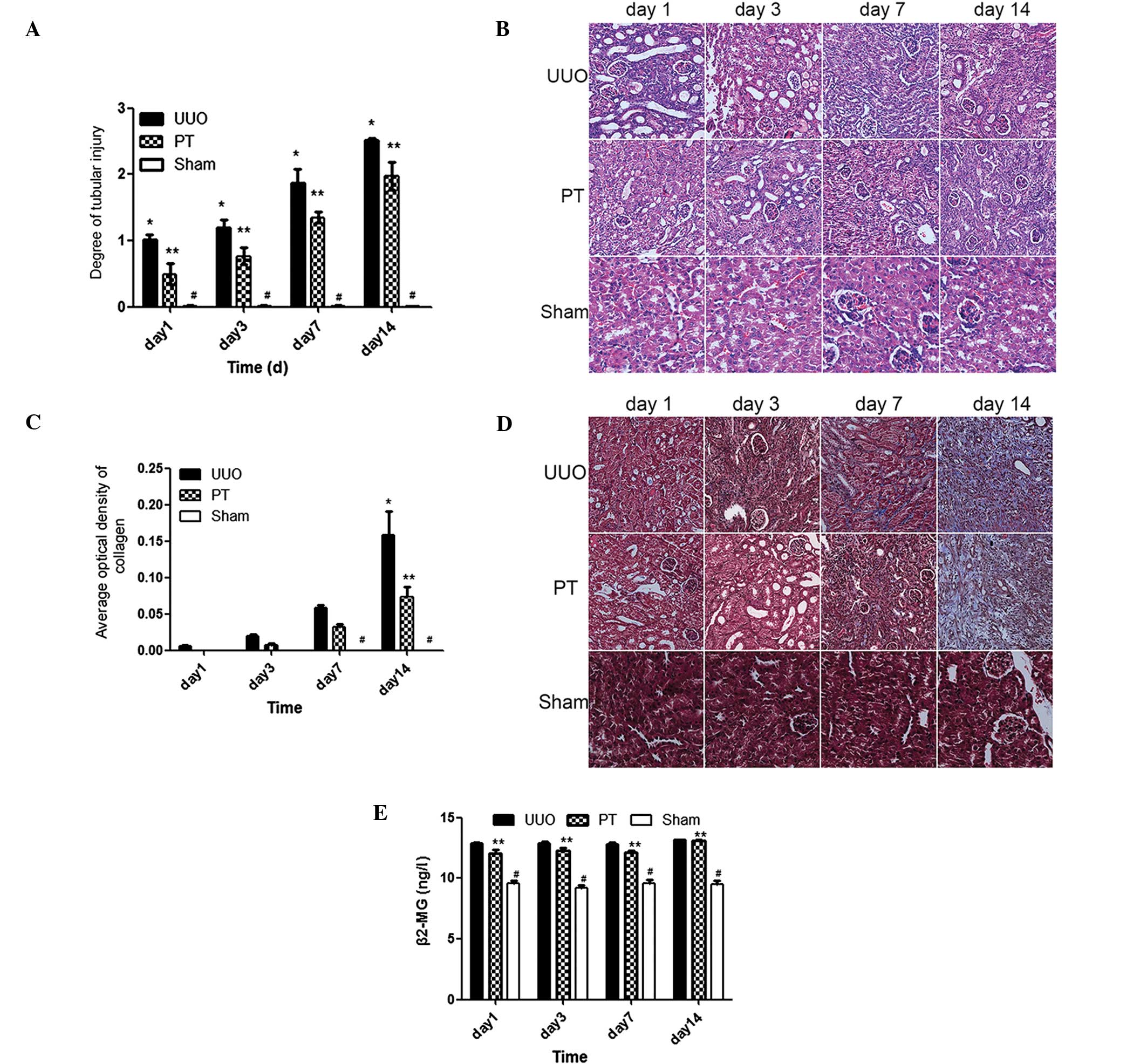
Promotion of RIF progress by increased
ADO concentration is reduced by blocking the ADO pathway
H&E staining revealed that the tubular injury
score in the UUO group was significantly higher than that in sham
group at the same time point (P<0.001). Tubular dilation,
accompanied by vacuolar degeneration, was observed on day 3 and
progressively increased over time following the UUO procedure; this
revealed a correlation between the post-UUO procedure time span and
the degree of injury (P<0.0001). The PT group demonstrated a
significantly reduced degree of tubular injury compared with that
in the UUO group (P<0.05); however, the degree of injury
remained higher than that in the sham group (Fig. 2A and B).
Tubular injury and the infiltration of interstitial
inflammatory cells was observed in the UUO group on day 3. This
progressively increased with time. In the PT group, the injury was
significantly lower compared with that in the UUO group.
Collagen expression in the renal tissue was detected
by Masson’s staining. On day 3, small amounts of collagen were
observed in the renal interstitial tissue in the UUO group. These
had significantly increased on day 7; on day 14, more collagen
tracts were observed, revealing typical interstitial fibrotic
changes. Collagen content in the UUO group on days 7 and 14 was
significantly higher compared with that in the sham group
(P<0.01). In the PT group, the collagen content was
significantly lower than that in the UUO group (P<0.001) and
significantly higher than that in the sham group (P<0.01;
Fig. 2C and D).
The parts of the renal tissue stained blue
represented collagen. Renal interstitial collagen content increased
with time. At each time point, the collagen content of the PT group
was lower than that of the UUO group.
The 24-h mouse urine sample was used for the
detection of β-2-microglobulin (β2-MG). In the UUO group
(Fig. 2E), the β2-MG
concentration in the 24-h urine was significantly higher than that
in the sham group (P<0.05) at each time point. In the PT group,
the β2-MG concentration was lower than that in the UUO
group; however, this difference was not statistically significant
(Fig. 2).
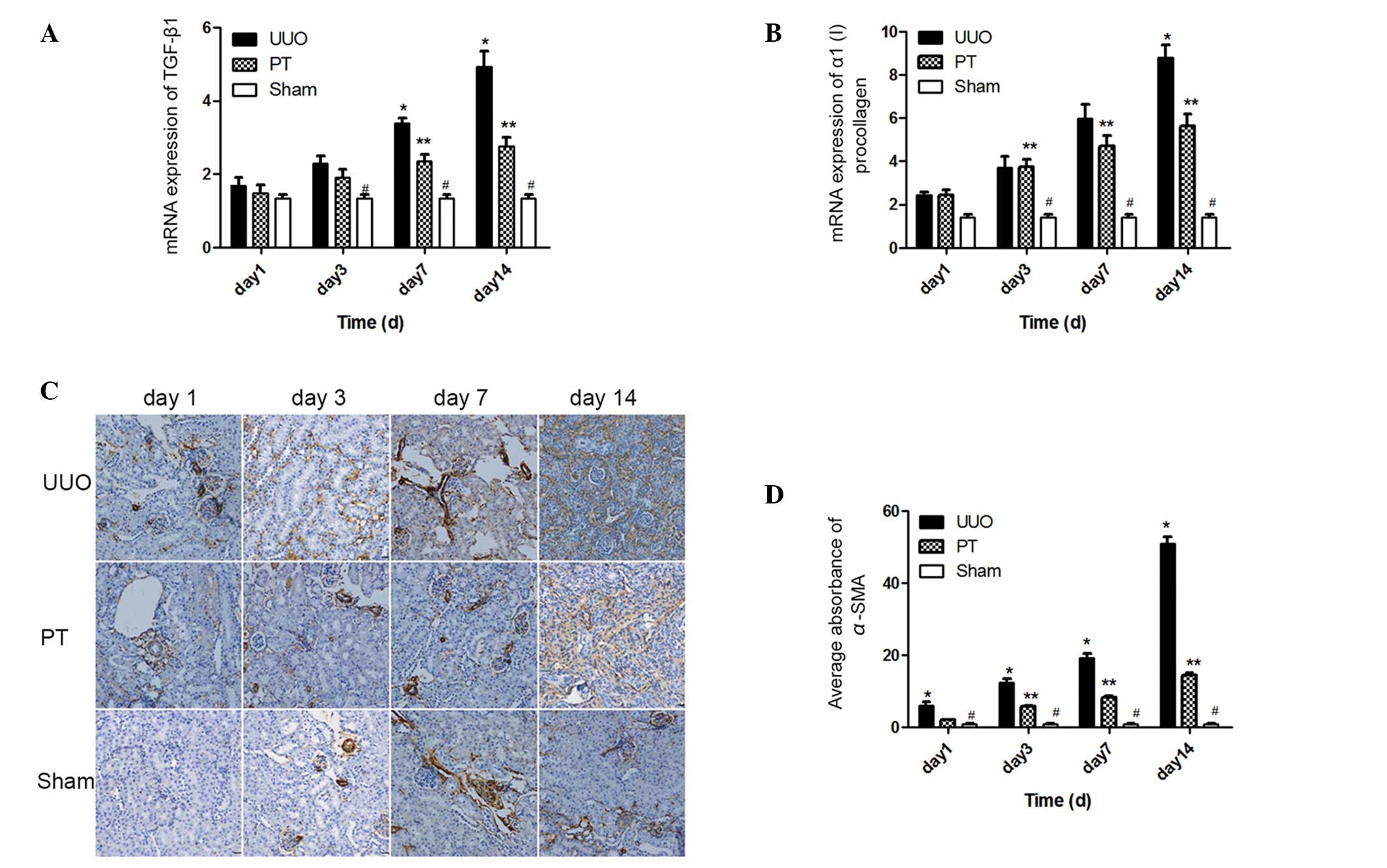
Role of the ADO signaling pathway
TGF-β1 and α1(I) procollagen
are two common profibrotic cytokines. The mRNA expression levels of
these two cytokines significantly increased with time
[TGF-β1, F=29.8; α1 (I) procollagen, F=32.85;
P<0.0001] in the UUO and PT groups. On days 3, 4 and 7, the mRNA
expression levels of TGF-β1 and α1(I)
procollagen in the UUO group were significantly higher than those
in the sham group (P<0.05). Following the blocking of the ADO
signaling pathway, the mRNA expression levels of the two cytokines
in the PT group were lower than those in the UUO group; however,
the expression levels were higher than those in the sham group
(P<0.05; Fig. 3A and B).
Compared with the UUO group, the t-statistic of TGF-β1
mRNA expression on days 7 and 14 was 3.490 and 7.375, respectively
(P<0.01); the t-statistic of α1(I) procollagen mRNA
expression on day 14 was 5.705 (P<0.001). These results indicate
that the ADO signaling pathway has a role in the regulation of
these profibrotic cytokines, and that TGF-β1 and
α1(I) procollagen expression was inhibited by ADO
pathway blockage.
The expression and distribution of α-SMA in the
renal tissue was dynamically observed by immunohistochemistry. In
the sham group, only a small amount of α-SMA was expressed in the
renal interstitial tissue blood vessels. In the UUO group, α-SMA
was widely present in the mesenchymal and renal tubular epithelial
cells. However, the expression was stronger than that in the sham
group (P<0.001). With the prolonging of the time following UUO,
α-SMA expression in the UUO group progressively increased.
Following ADO pathway blockage, the distribution of α-SMA was
consistent with that in the UUO group, but with a lower intensity
(P<0.001). However, it remained higher than in the sham group on
days 3, 7, and 14 (P<0.01; Fig. 3C
and D). Thus, the ADO pathway regulated α-SMA expression in the
renal tissue.
The parts of the renal tissue that were stained
brown represented α-SMA expression. From the first day following
modeling, the α-SMA expression in the UUO group was significantly
higher than that in the other two groups and increased with time.
The α-SMA expression in the PT group was lower than that in the UUO
group at all time periods, but remained higher than that in the
sham group.
Discussion
RIF is the most important factor in CKD progression
(14–16). Recent studies have indicated that
tubulointerstitial hypoxia may stimulate kidney disease and it is
considered to be one of the most important internal mechanisms of
RIF (17–19). ADO is an important signaling
molecule induced under ischemic and hypoxic conditions (2,3). In
the present study, marked tubular injury was observed following the
UUO procedure; the amount of interstitial collagen progressively
increased with time. Along with renal damage, the β2-MG
concentration in the UUO group was significantly higher than in the
sham group at each time period. The degree of renal tissue hypoxia
in the UUO group was significantly higher than in the sham group,
which peaked on day 3 following UUO and subsequently decreased with
time. A reason for this may be that in the time following the UUO
procedure, normal tissue cells were replaced by large amounts of
collagen, resulting in progressively decreased tissue protein
content and the reduction of its ability to bind pimonidazole.
Thus, the immunohistochemical staining decreased and the degree of
hypoxia exhibited an unusual reduction. Tissue hypoxia was present
during RIF in this part of the study.
Renal ischemia and hypoxia are conditions that are
able to induce ADO. It was confirmed by HPLC assay that following
the UUO procedure, the ADO concentration in the renal tissue was
significantly higher than in the sham group and that this was most
evident on days 1 and 3 after modeling. The concentration of ADO
decreased with time after UUO but remained higher than in the sham
group. The decreased ADO concentration was associated with
increased apoptosis, ATP depletion and increased renal interstitial
collagen levels. 8-PT was used in the current study as a
non-selective AR blocker to block the ADO signaling pathway.
Following the blocking of the ADO pathway, the ADO concentration in
the renal tissue of the PT group was higher than that in the UUO
group at the same time point. This result was related to the
reduced binding of ADO with AR following blockage and a slowed
metabolism, leading to extracellular ADO accumulation. Although the
ADO concentration increased in the PT group, the degree of tubular
injury, generation of renal interstitial collagen and
β2-MG concentration in the 24 h urine significantly
decreased compared with those in the UUO group. The ADO signaling
pathway is associated with RIF and following the ADO signaling
pathway blockage, RIF was effectively reduced and renal function
was protected.
The RIF process not only controls cell activation
and ECM deposition, but also maintains a close association with
cytokine interactions (1).
Presently, TGF-β1 is the most widely studied cytokine.
Increased TGF-β1 levels have been revealed to have a
causal association with tissue fibrosis (20). TGF-β1 has been confirmed
as the strongest accelerator of ECM accumulation currently known.
It functions through the TGF-β1/Smad signal transduction
pathway and is recognized as a target for RIF therapy (21,22).
In the current study, TGF-β1 mRNA content was detected
using qPCR, and was higher in the UUO group than in the sham group.
Following the blocking of the ADO pathway, the TGF-β1
mRNA level was significantly reduced. It was significantly lower in
the PT group than in the UUO group (P<0.01), particularly on
days 7 and 14. The above results indicate that hypoxia caused
persistently elevated ADO concentrations while inducing the
synthesis and secretion of large amounts of TGF-β1.
Following the blocking of the ADO signaling pathway,
TGF-β1 synthesis and secretion were inhibited, thus
protecting renal function.
Renal fibrosis is caused by the excessive synthesis
and decreased degradation of ECM (23), which is an important type of
collagen constituting the structural renal tissue framework. The
α-polypeptide chain is the basic subunit of collagen and
α1(I) procollagen is one of the major components
involved in collagen deposition. Thus, in the present study, the
degree of renal fibrosis in renal tissue was assessed by the
α1(I) procollagen content. From day 3 after modeling,
α1(I) procollagen content in the renal tissue in the UUO
group was consistently higher than in the sham group (P<0.05),
indicating that large amounts of collagen had accumulated outside
of the cells; thus, the renal tissue gradually experienced
fibrosis. Following the blocking of the ADO pathway, the mRNA
content of α1(I) procollagen was lower in the PT group
than in the UUO group from day 7 post-modeling. The content
continued to decrease and was significantly lower than that in the
UUO group on day 14 (P<0.001). This result indicates that the
ADO signaling pathway maintains a balance between the synthesis and
degradation of the ECM and that this has positive effects in
protecting renal function and repairing renal damage.
Fibroblasts are cells that are inherently present in
the renal interstitium. They are relatively sparse under normal
physiological conditions but greatly increase in number in fibrotic
states. Fibroblasts are important in the development of RIF as they
are ECM-secreting cells. When stimulated by inflammation, toxins or
an immune response, fibroblasts in the renal interstitial region
are activated, causing proliferative and phenotypic changes. The
most representative change is the activation and transformation of
fibroblasts into myofibroblasts (MFBs), which express α-SMA. The
amount of collagen produced by MFB is 4–5 times as much as that
produced by fibroblasts and this significantly influences the RIF
process. Thus, a key step in RIF is the transformation of
fibroblasts into α-SMA-expressing myofibroblasts (MFB). A study by
Iwano et al reported that in the UUO model, ~36% of the
fibroblasts were derived from the epithelial-interstitium
transdifferentiation processes (24). In the present study, the expression
of α-SMA in the renal tissue at different times was observed by
immunohistochemistry. Following the UUO procedure, α-SMA expression
in the renal interstitial and tubular tissue in the UUO group was
significantly increased compared with that in the sham group
(P<0.001). Following the blocking of the ADO pathway, α-SMA
expression in the UUO group was significantly reduced compared with
that in the UUO group (P<0.01). These results indicate that the
ADO signaling pathway is associated with the proliferation and
activation of the epithelial-mesenchymal transition (EMT) and
fibroblasts. Notably, these processes were inhibited following ADO
blockage.
In summary, the current study confirmed that ADO
regulates the pathological processes of RIF using the mice UUO
model. This provides an important experimental basis for further
investigations of RIF pathogenesis. Furthermore, the association
between AR types and RIF in the ADO signaling pathway, as well as
the biological changes of important fibrotic effector cells, merits
further study in order to provide novel antifibrotic therapy
proposals for patients with CKD.
Acknowledgements
This study was supported by the Science and
Technology Program of Hunan Scientific Committee and the ‘125’
Excellent Youth Foundation of the Third Xiangya Hospital of Central
South University (grant no. 2012FJ3134).
References
|
1
|
Suzuki K, Wang R, Kubota H, Shibuya H,
Saegusa J and Sato T: Kinetics of biglycan, decorin and
thrombospondin-1 in mercuric chloride-induced renal
tubulointerstitial fibrosis. Exp Mol Pathol. 79:68–73. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fredholm BB: Adenosine, an endogenous
distress signal, modulates tissue damage and repair. Cell Death
Differ. 14:1315–1323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eltzschig HK, Thompson LF, Karhausen J, et
al: Endogenous adenosine produced during hypoxia attenuates
neutrophil accumulation: coordination by extracellular nucleotide
metabolism. Blood. 104:3986–3992. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lasley RD, Rhee JW, Van Wylen DG and
Mentzer RM Jr: Adenosine A1 receptor mediated protection of the
globally ischemic isolated rat heart. J Mol Cell Cardiol. 22:39–47.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fishman P, Bar-Yehuda S, Farbstein T,
Barer F and Ohana G: Adenosine acts as a chemoprotective agent by
stimulating G-CSF production: a role for A1 and A3 adenosine
receptors. J Cell Physiol. 183:393–398. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Linden J: Adenosine in tissue protection
and tissue regeneration. Mol Pharmacol. 67:1385–1387. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu B, Rajakumar SV, Robson SC, et al: The
impact of purinergic signaling on renal ischemia-reperfusion
injury. Transplantation. 86:1707–1712. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hatfield S, Belikoff B, Lukashev D,
Sitkovsky M and Ohta A: The antihypoxia-adenosinergic pathogenesis
as a result of collateral damage by overactive immune cells. J
Leukoc Biol. 86:545–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dai Y, Zhang W, Wen J, Zhang Y, Kellems RE
and Xia Y: A2B adenosine receptor-mediated induction of IL-6
promotes CKD. J Am Soc Nephrol. 22:890–901. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grenz A, Baier D, Petroktistis F, et al:
Theophylline improves early allograft function in rat kidney
transplantation. J Pharmacol Exp Ther. 317:473–479. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sobh M, Sabry A, Moustafa F, Foda MA,
Sally S and Ghoneim M: Effect of colchicine on chronic ciclosporin
nephrotoxicity in Sprague-Dawley rats. Nephron. 79:452–457. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mizuguchi Y, Miyajima A, Kosaka T, Asano
T, Asano T and Hayakawa M: Atorvastatin ameliorates renal tissue
damage in unilateral ureteral obstruction. J Urol. 172:2456–2459.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu C, Ding X, Zhu J, Fu C and Zhong Y:
Preliminary study of the role of hypoxia in the ligated kidney of
unilateral ureteral obstruction rat model. Fudan University Journal
of Medical Sciences. 34:732–736. 2007.(In Chinese).
|
|
14
|
Coresh J, Astor BC, Greene T, Eknoyan G
and Levey AS: Prevalence of chronic kidney disease and decreased
kidney function in the adult US population: Third National Health
and Nutrition Examination Survey. Am J Kidney Dis. 41:1–12. 2003.
View Article : Google Scholar
|
|
15
|
Zhang L, Zhang P, Wang F, et al:
Prevalence and factors associated with CKD: a population study from
Beijing. Am J Kidney Dis. 51:373–384. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hewitson TD: Renal tubulointerstitial
fibrosis: common but never simple. Am J Physiol Renal Physiol.
296:F1239–1244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fine LG, Orphanides C and Norman JT:
Progressive renal disease: the chronic hypoxia hypothesis. Kidney
Int Suppl. 65:S74–78. 1998.PubMed/NCBI
|
|
18
|
Fine LG, Bandyopadhay D and Norman JT: Is
there a common mechanism for the progression of different types of
renal diseases other than proteinuria? Towards the unifying theme
of chronic hypoxia. Kidney Int Suppl. 75:S22–26. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fine LG and Norman JT: The breathing
kidney. J Am Soc Nephrol. 13:1974–1976. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Y: Renal fibrosis: new insights into
the pathogenesis and therapeutics. Kidney Int. 69:213–217. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Zhang Z, Wang D, Wang Y, Li Y and Wu
G: TGF-beta 1/Smads signaling stimulates renal interstitial
fibrosis in experimental AAN. J Recept Signal Transduct Res.
29:280–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Phanish MK, Wahab NA, Colville-Nash P,
Hendry BM and Dockrell ME: The differential role of Smad2 and Smad3
in the regulation of pro-fibrotic TGFbeta1 responses in human
proximal-tubule epithelial cells. Biochem J. 393:601–607. 2006.
View Article : Google Scholar :
|
|
23
|
van Kleef EM, Zurcher C, Oussoren YG, et
al: Long-term effects of total-body irradiation on the kidney of
Rhesus monkeys. Int J Radiat Biol. 76:641–648. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Iwano M, Plieth D, Danoff TM, Xue C, Okada
H and Neilson EG: Evidence that fibroblasts derive from epithelium
during tissue fibrosis. J Clin Invest. 110:341–350. 2002.
View Article : Google Scholar : PubMed/NCBI
|