Introduction
Epithelial tissue, in addition to forming a physical
barrier to external stimuli, plays a major role in the regulation
of the adaptive immune response. The physical barrier function of
the epithelium is attained via cell-cell interactions and via
impermeable tight junctions (TJs) that are located subapically
(1).
Claudins, occludin and junctional adhesion molecules
(JAMs) are major constituent proteins of TJs. The claudin family
comprises 24 members, and determines the TJ charge selectivity and
the location of the paracellular space (2). In addition, human fetal lung cells have
been shown to use differential claudin expression to regulate TJ
permeability (3). Occludin is an
additional protein that is important in barrier formation and
regulation (2). JAM-1, which is a
member of the immunoglobulin superfamily, has been shown to
interact with zonula occludens (ZO)-1, occludin and cingulin, which
form the TJ; JAM-1 is hypothesized to affect signal cascades
resulting from homophilic and heterophilic adhesion (4). Epithelial barrier function is impaired
in individuals with asthma, which is associated with increased
permeability to macromolecules, such as allergens. In addition,
epithelial barrier impairment in asthmatic bronchi has been shown
to be associated with a patchy intercellular distribution of TJs
and a decrease in the expression levels of TJ proteins, including
ZO-1 and occludin (5).
Tumor necrosis factor (TNF)-α has been shown to
reduce claudin expression in a variety of tissues (6–8).
However, the mechanism underlying this effect remains unclear with
different studies reporting variable results. One study reported
that the inflammatory cytokines, TNF-α and interferon-γ, cause a
dose- and time-dependent increase in intestinal epithelial
permeability by inducing the redistribution of TJ proteins,
including claudin-1, claudin-4 and JAM-2, without causing a
significant decrease in their total levels (7). However, an additional study reported
that the levels of claudin-1 and claudin-4 did not markedly change,
while the downregulation of JAM was observed in a human airway
epithelial cell line (9).
Furthermore, a previous study reported that TNF-α causes the
decreased expression of active occludin molecules, which leads to
the disruption of TJ structure in Caco-2 cells (8). However, an additional study reported
that TNF-α causes an increase in the levels of active occludin in
human airway epithelial cells, which was hypothesized to have
occurred due the presence of a leaky junction (8,9). Thus,
TNF-α antagonism is hypothesized to improve epithelial barrier
integrity.
There is little knowledge with regard to the
influence of vascular endothelial growth factor (VEGF) antagonism
on epithelial barrier function and TJ structure. The majority of
data on the issue are from studies using the retinal endothelium,
which have indicated that VEGF exposure leads to a decrease in
transendothelial resistance, an increase in vascular permeability,
and a decrease in the expression levels of claudin-1 and occludin
(10,11). Furthermore, VEGF antagonism has been
demonstrated to reverse the decrease in transendothelial resistance
and claudin-1 levels induced by VEGF in retinal endothelial cells
(12).
Steroid treatment has been shown to improve
epithelial barrier integrity, increase transepithelial resistance
and lead to a more integrated expression of the TJ proteins,
occludin and ZO-1. These effects have been proposed to be mediated
through epidermal growth factor receptor phosphorylation (13).
Investigating the effects of different asthma
treatments on the epithelial barrier proteins, occludin, claudin
and JAM, may identify possible therapeutic alternatives and
targets. Therefore, the aim of the present study was to investigate
the change in the levels of the TJ proteins, occludin, claudin and
JAM, in models of healthy and asthmatic mice, and to compare the
influence of TNF and VEGF inhibition and steroid treatment on these
changes.
Materials and methods
Experimental animals
A total of 38 BALB/c mice (age, 6–8 weeks; weight,
18–20 g) were used in the experiment. The mice were housed in
hygienic macrolene cages in air-conditioned rooms on a 12-h
light/dark cycle. Food and water were provided ad libitum.
The animals were maintained at a constant temperature of 22°C in
the Experimental Animal Unit of Dokuz Eylul University (Izmir,
Turkey).
This study was conducted in the Dokuz Eylul
University multidisciplinary laboratory and approved by the
university ethical board. Furthermore, the study was conducted in
accordance with the recommendations outlined in the Guidelines for
the Care and Use of Experimental Animals (14).
Study design
The study comprised five groups of mice. Control
group (n=6) mice were not exposed to ovalbumin (OVA) or any other
medication. A chronic asthma model was established in the other
four groups (n=8 per group) through application of intraperitoneal
(IP) and inhaled OVA exposure. Each of these latter four groups
received as follows: IP saline injection once per week for 2 weeks;
IP injection of etanercept (Wyeth Europa Ltd, Maidenhead, UK) at
0.01 mg/dose (0.5 mg/kg) twice per week for two weeks; IP injection
of bevacizumab (Avastin; Roche Diagnostics, Basel, Switzerland) at
0.15 mg (5 mg/kg) once per week for two weeks; or 1 mg/kg/dose IP
injection of dexamethasone (Dekort; Deva Holding AS, Istanbul,
Turkey) for 7 days during the final week of OVA inhalation.
Sensitization procedure and inhalation
exposure
A chronic murine model of asthma, as described by
Temelovski et al (15), was
successfully established. An injection of 10 µg OVA (grade V, ≥98%
pure; Sigma-Aldrich, St. Louis, MO, USA), at a volume of 0.1 ml,
was administered (IP) to the BALB/c mice (with the exception of the
Group 1 mice) at days 21 and 7 prior to OVA inhalation, with alum
as the adjuvant. Consequently, the mice were exposed to a 2.5% OVA
aerosol in sterile saline for 30 min/day for three days/week for a
total of eight weeks, starting on day 21 of the experiment.
Exposure was conducted using a whole body inhalation exposure
system in a plexiglass chamber with 40×60×120 cm volume, designed
for the placement of cages. A particle concentration of 10–20
mg/m3 was achieved, and ≥80% of the particles were
expected to have a diameter of ≤4 µm, due to the characteristics of
the system used.
The mice were anesthetized by an IP injection of
ketamine hydrochloride (200 mg/kg) at day 1 following the
administration of the final medication, after which the mice were
sacrificed via exsanguination cardiac arrest.
Histopathological and
immunohistochemical evaluations
Formalin-fixed, paraffin-embedded lung samples
collected from the control and experimental mice were prepared by
routine paraffin-embedding procedures. In brief, tissue samples
were fixed in 10% formalin in phosphate buffer (pH 7.4) for 48 h,
dehydrated in a graded series of ethanol (70%, 80%, 90% and
absolute ethanol), cleared in xylene and embedded in paraffin. The
paraffin blocks were serially sectioned using a Leica RM2245 rotary
microtome (Leica Microsystems Gmbh, Wetzlar, Germany) at a
thickness of 5 µm, and prepared for histochemical and
immunohistochemical staining. For histological assessment, the
sections were stained with hematoxylin and eosin.
For immunohistochemical staining, the sections were
incubated at 60°C overnight, cleared in xylene for 30 min, and
washed with decreasing concentrations of ethanol, followed by
washing with distilled water. After incubation in
phosphate-buffered saline PBS (P4417; Sigma-Aldrich) for 10 min,
the sections were treated with 2% trypsin (TSS155; Scytek
Laboratories, Inc., Logan, UT, USA) at 37°C for 10 min and washed
with PBS. The sections were delineated with a Dako pen (s2002;
Dako, Glostrup, Denmark), incubated in a solution of 3% hydrogen
peroxide (K31355100; Merck, Darmstadt, Germany) for 15 min to
inhibit endogenous peroxidase activity, and washed with PBS.
Subsequently, primary antibodies were added and the sections were
incubated in a humidified chamber at 4°C overnight. The following
primary antibodies were used at a 1:100 dilution: Claudin-1
antibody (A-9; sc-166338; mouse monoclonal IgG2b; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), occludin antibody
(sc-133256; mouse monoclonal IgG2b; Santa Cruz Biotechnology, Inc.)
and JAM-A antibody (H-80; sc-25629; rabbit polyclonal IgG; Santa
Cruz Biotechnology, Inc.).
Following removal of the primary antibodies, the
sections were washed with PBS three times for 5 min each and
incubated with a biotinylated secondary antibody for 30 min, using
a Histostain®-Plus-Peroxidase kit (85–9043; Invitrogen Life
Technologies, Carlsbad, CA, USA). Subsequently, the sections were
incubated with streptavidin conjugated to horseradish peroxidase in
PBS for 30 min, using a Histostain®-Plus Bulk kit, Invitrogen 2nd
Generation, labeled-(strept)avidin-biotin Detection System
(85–8943; Invitrogen Life Technologies). The sections were
subsequently washed with PBS three times and incubated for 5 min
with 3,3′-diaminobenzidine (00–2020; Invitrogen Life Technologies)
as a chromogen. After washing with distilled water, the sections
were counterstained with Mayer's hematoxylin solution
(#02274390059; J.T. Barker, Deventer, The Netherlands), washed with
tap water, dehydrated through a graded series of alcohol and
cleared in xylene. The sections were mounted with Entellan mounting
medium (HX265767; Merck Millipore, Darmstadt, Germany) and
evaluated under a light microscope (BX40; Olympus Corporation,
Tokyo, Japan). Control samples were processed in an identical
manner, with the exception that IgG was used instead of a primary
antibody.
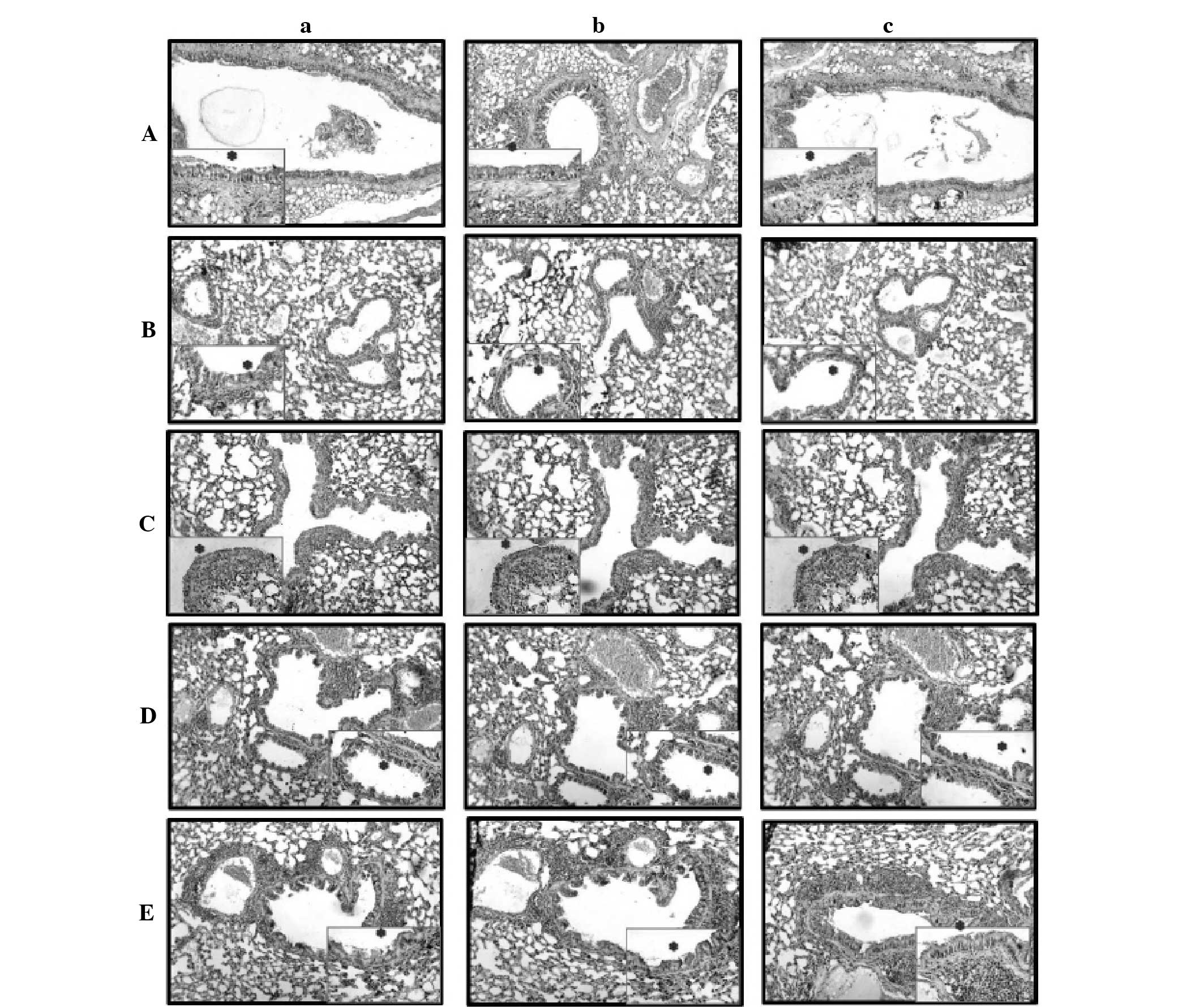
The distribution of immunohistochemical intensities
of the primary antibodies was scored as mild (+), moderate (++),
strong (+++) and very strong (++++) brown staining. Staining
intensity was graded semiquantitatively using H-scores, which were
calculated using the following equation: H-score = ΣPi (i + 1),
where i was equal to the intensity of immunohistochemical staining
with a value of 1–4, and Pi was the percentage of epithelial cells
stained with each intensity, varying between 0–100% (16).
Statistical analysis
Statistical analysis was performed using SPSS 15.0
software (SPSS, Inc., Chicago, IL, USA). The Kruskal-Wallis
non-parametric test was used to compare the results among all four
treatment groups, while the Mann-Whitney U test was used for
pairwise comparison of the groups. P<0.05 was considered to
indicate a statistically significant difference. The Bonferroni
correction was applied for the post-hoc analysis results, where
P<0.005 was regarded as a statistically significant difference
for the two group comparisons.
Results
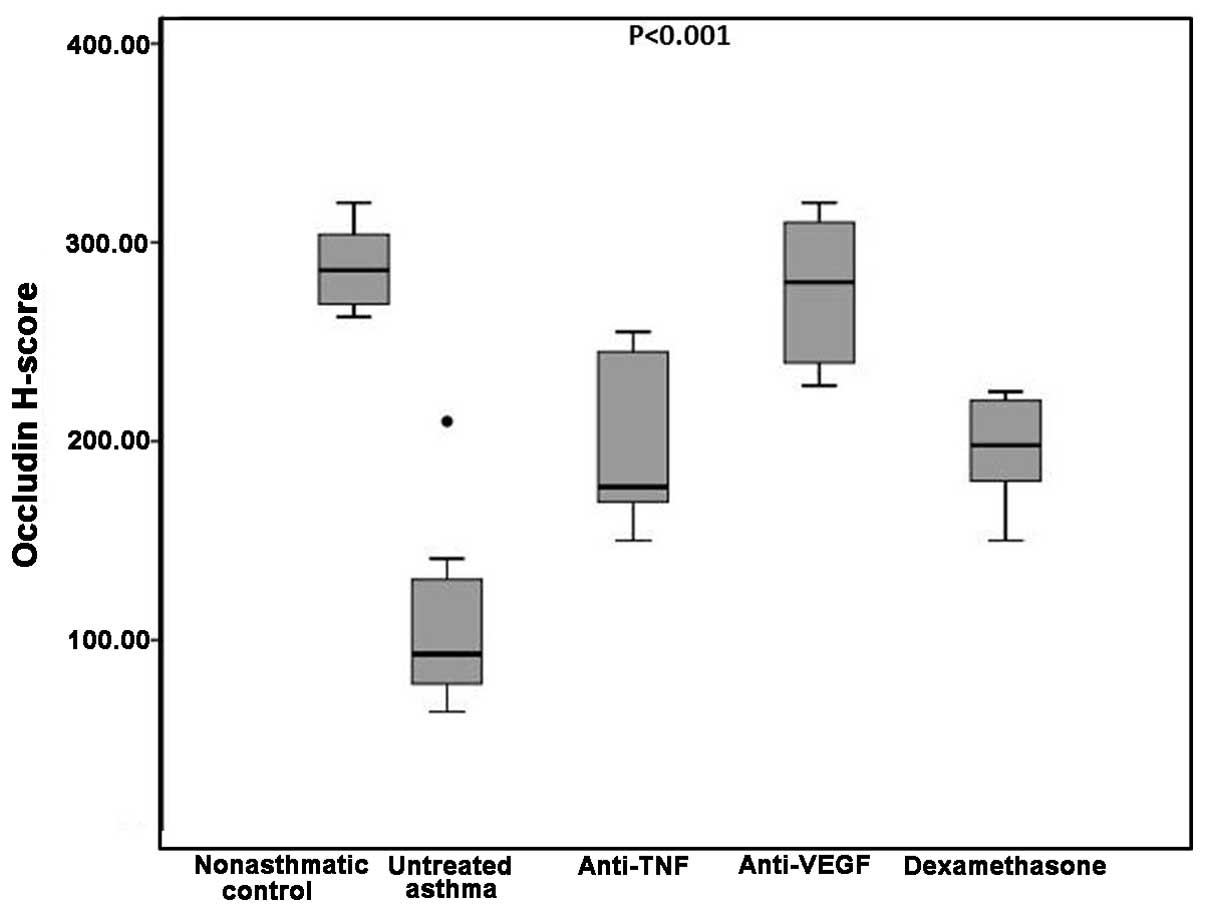
Occludin H-scores among the
groups
The difference in the occludin H-scores was
determined to be statistically significant when the five groups
were compared (P<0.001). In the non-asthmatic control group, the
median occludin H-score was 286, which was significantly higher
compared with the untreated asthma, etanercept and dexamethasone
groups (93, 177 and 198 respectively; P=0.002 for all). By
contrast, the difference in the occludin H-scores between the
bevacizumab group and non-asthmatic control group was not
statistically significant (280 vs. 286 respectively; P=0.60;
Table I).
 | Table I.Occludin, claudin and JAM H-scores in
the study groups. |
Table I.
Occludin, claudin and JAM H-scores in
the study groups.
| Study groups | Occludin H-score | Claudin H-score | JAM H-score |
|---|
| Non-asthmatic
control | 286 (269–304) | 306 (300–320) | 274 (259–304) |
| Untreated asthma | 93 (78–130.5) | 82 (76.5–108) | 130 (106.3–160) |
| Etanercept | 177 (169.5–245) | 193.5
(186–238.3) | 210 (184.5–236) |
| Bevacizumab | 280 (239.5–310) | 274 (229.8–306) | 288 (243–304) |
| Dexamethasone | 198 (180–220.5) | 202.5 (195–210) | 210 (186–231) |
| P-value | P<0.001 | P<0.001 | P<0.001 |
Occludin H-scores in the untreated asthma group were
significantly lower compared with the etanercept, bevacizumab and
dexamethasone groups (P=0.005, P=0.001 and P=0.004,
respectively).
Comparison of the treatment groups with each other
revealed that the differences in the occludin H-scores between the
etanercept group and the bevacizumab or dexamethasone groups were
not statistically significant (P=0.01 and P=0.79, respectively). By
contrast, the bevacizumab group had a significantly higher median
occludin H-score when compared with the dexamethasone group (280
vs. 198 respectively; P=0.001; Table
I and Figs. 1 and 2).
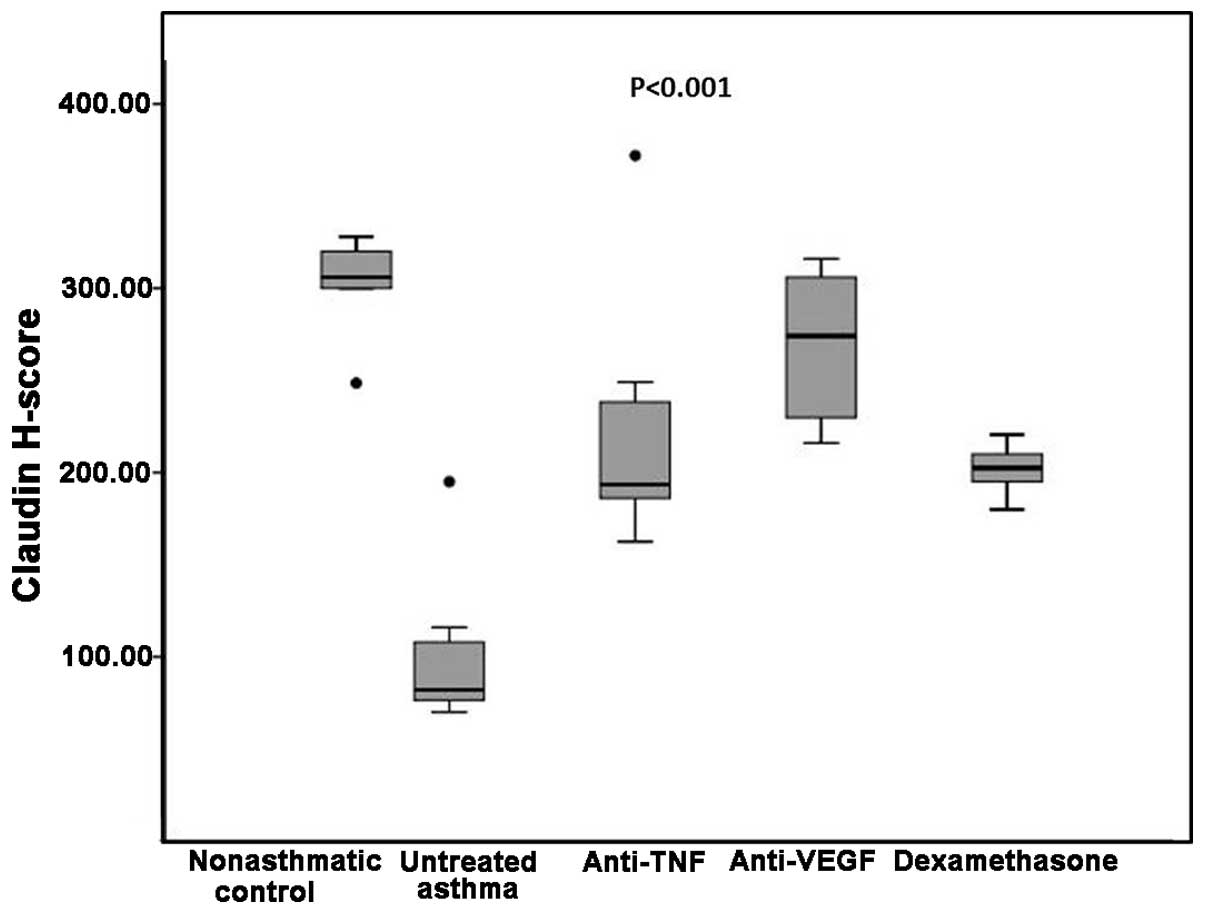
Claudin H-scores among the groups
Median claudin H-scores were significantly different
among the groups. In the non-asthmatic control group, the claudin
H-score was significantly higher compared with the untreated asthma
and dexamethasone groups (306 vs. 82 and 202.5, respectively;
P=0.002 for the two comparisons); however, there were no
statistically significant differences when comparing the
non-asthmatic control with the etanercept or bevacizumab groups
(193.5 and 274, respectively; P=0.03 and P=0.11 respectively;
Table I). The untreated asthma group
had a significantly lower claudin H-score compared with the three
treatment groups (82 vs. 193.5 in the etanercept, 274 in the
bevacizumab and 202.5 in the dexamethasone groups; P=0.004, P=0.001
and P=0.002, respectively; Table
I).
Comparison of the three treatment groups with each
other revealed that the dexamethasone group had similar claudin
H-scores compared with the etanercept group, but lower claudin
H-scores when compared with the bevacizumab group (P=0.75 and
P=0.001, respectively). Similarly, etanercept treatment was not
significantly different from the bevacizumab treatment in terms of
the levels of claudin (P=0.05; Table
I and Figs. 1 and 3).
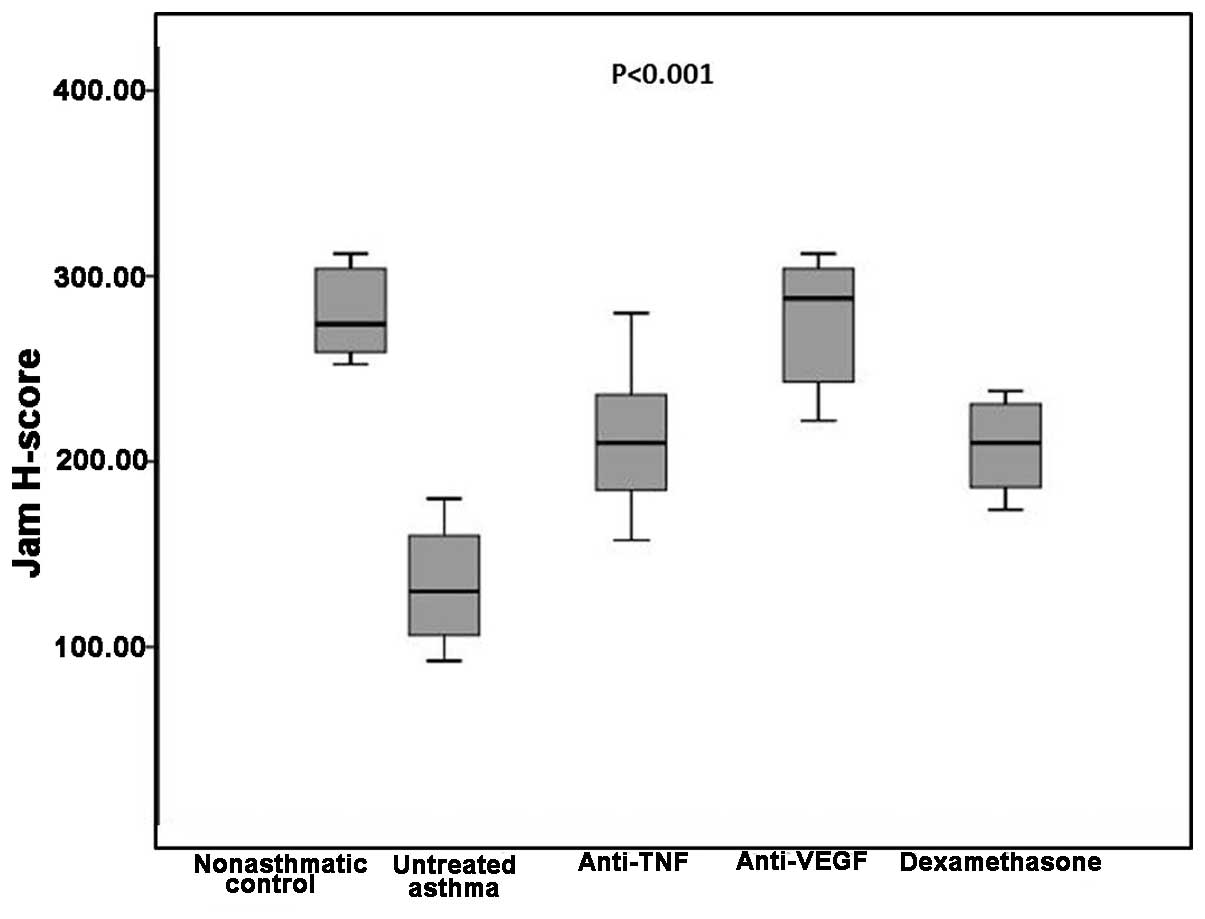
JAM H-scores among the groups
The median JAM H-score in the non-asthmatic control
group was 274, which was significantly higher compared with the
untreated asthma and dexamethasone groups (P=0.002 for both).
However, no statistically significant difference was detected when
this was compared with the etanercept and bevacizumab groups
(P=0.01 and P=0.89, respectively). The untreated asthma group had a
median JAM H-score of 130, which was significantly lower compared
with all three treatment groups (P=0.002, P=0.001 and P=0.001 for
the etanercept, bevacizumab and dexamethasone groups, respectively;
Table I).
Comparisons between the three treatment groups
revealed that the etanercept group had similar JAM H-scores when
compared with the bevacizumab and dexamethasone groups (P=0.007 and
P=0.87 respectively). However, the bevacizumab group had
significantly higher JAM H-scores compared with the dexamethasone
group (P=0.004; Table I and Figs 1 and 4).
Discussion
The present study demonstrated that the levels of
the TJ proteins, occludin, claudin and JAM, were decreased in a
murine model of asthma. Immunohistochemical evaluation of the
expression levels of occludin, claudin and JAM following the
different asthmatic treatment modalities revealed that treatment
with steroids, as well as TNF and VEGF antagonists, increased the
levels of the proteins when compared with untreated asthma group.
The only difference among the three treatment groups was between
dexamethasone and bevacizumab (VEGF antagonism), which was in favor
of the latter. Furthermore, TNF and VEGF antagonism resulted in
levels of claudin and JAM that were similar to the non-asthmatic
control, while only VEGF antagonism led to levels of occludin that
were similar to the non-asthmatic control.
Asthma is a chronic inflammatory disease of the
airways that originally was attributed to a deviated adaptive
immune system, primarily affecting T helper 2 lymphocytes; however,
novel pathogenetic results have indicated the role of the innate
immune system, including the epithelium, in the pathogenesis of
asthma (17). Under physiological
conditions, the airway epithelium forms a barrier to inhaled
antigens and allergens; however, this barrier is impaired in
individuals with asthma and the permeability to inhaled allergens
and pollutants is increased, which is hypothesized to be the
mechanism underlying the development of asthma in certain atopic
individuals (18,19). Concordantly, the present study
demonstrated significantly decreased levels of the TJ proteins,
occludin, claudin and JAM, in the bronchial epithelium of the
asthmatic mice, as compared with the non-asthmatic mice, indicating
their role in the pathogenesis of asthma.
Epithelial barrier function is predominantly
controlled by TJs that constitute a continuous structure between
the apical and basolateral membranes, which regulate the exchange
of substances between intracellular and extracellular environments.
Occludin, claudin and JAM-1 are the main components controlling the
barrier function of TJs through sealing the paracellular space.
Occludin copolymerases into claudin-based TJ strands; however, the
protein is not required for the formation of TJ strands, but for
the regulation of permeability and transepithelial resistance. The
claudin family, which comprises 24 members, forms the backbone of
the TJ structure and is considered to play a role in the formation
of the ion selective pores within the TJ strands. JAMs belong to
the immunoglobulin superfamily, and are not part of the TJ strands;
however, these proteins interact with TJ proteins, including ZO-1
and cingulin. JAM 1 is hypothesized to play a role in the
propagation of signals as a result of homophilic and heterophilic
adhesion to other molecules (4). The
aim of the current study was to investigate the changes in the
levels of these three major proteins, which play a role in the
formation of the epithelial barrier, in a mouse model of asthma
subjected to different treatment conditions, in an attempt to
identify potential therapeutic targets and agents.
Glucocorticoids have been shown to increase
endothelial barrier properties by increasing the gene expression
levels of occludin and claudin-5 in the retinal endothelium
(20). Similarly, the results of the
current study revealed increased levels of occludin, claudin and
JAM in a mouse model of asthma treated with steroids. These
observations indicate the role of conventional steroid treatment in
epithelial repair and barrier improvement in asthma.
The role of VEGF has been commonly studied in models
of diabetic retinopathy and retinal endothelial cells (11,21).
Considering the important role of VEGF in asthma and remodeling,
the current study aimed to investigate the effects of VEGF
antagonism on bronchial epithelial barrier function (22). VEGF has been observed to cause the
phosphorylation of occludin and a decrease in the level of occludin
in retinal endothelial cells in diabetic retinopathy, subsequently
increasing endothelial permeability (11,22).
This finding is concordant with the present results that
demonstrated an increase in the level of occludin in the asthmatic
mice treated with bevacizumab, resulting in a level that was
similar to the non-asthmatic control mice. Comparison of VEGF
antagonistic treatment with conventional steroid treatment in the
asthmatic mice revealed a higher level of bronchial epithelial
occludin in the former group. Furthermore, VEGF antagonism has been
shown to increase transendothelial electrical resistance in bovine
retinas and reverse VEGF-induced loss of claudin-1 protein. In
addition, VEGF antagonists have been shown to cause the
colocalization of claudin-1 and claudin-5 to the plasma membrane,
supporting a stable barrier function (12). The present study detected that the
level of claudin in the bronchial epithelium increased with VEGF
antagonistic treatment in a murine asthma model, resulting in
levels that were similar to those of the non-asthmatic controls.
Furthermore, the increase in the level of occludin observed with
VEGF antagonism was significantly more prominent compared with
conventional steroid treatment. The influence of VEGF antagonism on
the levels of bronchial JAM was also investigated. JAM levels were
shown to increase to a similar level to that observed in the
non-asthmatic mice, and this effect was significantly higher
compared with the conventional steroid treatment. Overall, VEGF
antagonistic treatment appeared to reverse the decrease in the
levels of the epithelial barrier TJ proteins, occludin, claudin and
JAM, in a mouse model of asthma, and this effect was more
pronounced when compared with conventional steroid treatment. Thus,
VEGF may be a potential therapeutic target for improving epithelial
barrier dysfunction in asthma.
TNF-α expression is known to result in a decrease in
the levels of occludin and claudin, and the redistribution of p120
catenin and E-cadherin, which subsequently compromises the
properties of TJs and adherens junctions. In turn, the epithelial
barrier function is compromised in different tissues, including the
brain and lungs (6,23). Subsequently, a decrease in
transepithelial electrical resistance occurs, and increased
epithelial permeability in asthmatic air-liquid interface cultures
has been observed (23). TNF-α is
not only effective on bronchial epithelial TJs, but also on other
tissues, including retinal epithelial cells, where the cytokine
increases the levels of matrix metalloproteinases, which in turn
degrade claudin-1 and occludin, increasing permeability (24). In concordance with these previously
reported effects of TNF-α, antagonism of TNF-α in the present mouse
model of asthma resulted in an increase in the levels of occludin
and claudin in the bronchial epithelium. Furthermore, the level of
an additional TJ protein, JAM, was also shown to increase with
TNF-α antagonistic treatment in asthma. These effects of TNF-α
antagonistic treatment appeared to be similar to conventional
steroid treatment, but inferior to VEGF antagonistic treatment.
Therefore, TNF-α antagonistic treatment in steroid-resistant cases
of asthma may not be expected to provide additional therapeutic
effects on bronchial epithelial barrier function.
One of the main limitations of the present study was
the use of the semiquantitative immunohistochemical H-scores. In an
attempt to avoid potential bias, the histologist was blinded to the
groups when scoring. Furthermore, the use of a mouse model limits
the generalization to humans. However, mouse models of asthma are
generally accepted to correlate well with clinical human results
(25). Finally, the use of animal
models precludes the prediction of side effects when these agents
are used in humans; however, dexamethasone and etanercept are
already widely administered. With regard to bevacizumab, further
clinical research is required in order to propose any
recommendations for human administration.
The major contribution of the present study to the
current literature is the comparison of the influence of different
therapeutic agents on epithelial barrier proteins, which are of
current interest in investigations into the pathogenesis of asthma.
Furthermore, the effect of VEGF inhibition on the bronchial
epithelium in asthma has not previously been studied
extensively.
In conclusion, the present study demonstrated that
the epithelial barrier proteins, occludin, claudin and JAM, are
significantly decreased in mouse models of asthma, which results in
the impairment of epithelial barrier function. Conventional
treatment with steroids was shown to increase the levels of these
proteins to a similar degree as TNF-α antagonistic treatment, which
subsequently raised the question of the efficacy of TNF-α
antagonistic treatment on barrier enhancement in steroid-resistant
cases. VEGF antagonists may be potential novel therapeutic agents,
since treatment with bevacizumab was shown to increase the
expression levels of the bronchial epithelium TJ proteins. However,
further clinical research investigating the in vivo efficacy
and side-effect profile of bevacizumab in cases of asthma is
required prior to the establishment of specific
recommendations.
References
|
1
|
Holgate ST: Epithelium dysfunction in
asthma. J Allergy Clin Immunol. 120:1233–1244. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marchiando AM, Graham WV and Turner JR:
Epithelial barriers in homeostasis and disease. Annu Rev Pathol.
5:119–144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daugherty BL, Mateescu M, Patel AS, Wade
K, Kimura S, Gonzales LW, Guttentag S, Ballard PL and Koval M:
Developmental regulation of claudin localization by fetal alveolar
epithelial cells. Am J Physiol Lung Cell Mol Physiol.
287:L1266–L1273. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Förster C: Tight junctions and the
modulation of barrier function in disease. Histochem Cell Biol.
130:55–70. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao C, Puddicombe SM, Field S, Haywood J,
Broughton-Head V, Puxeddu I, Haitchi HM, Vernon-Wilson E, Sammut D,
Bedke N, Cremin C, Sones J, Djukanović R, Howarth PH, Collins JE,
Holgate ST, Monk P and Davies DE: Defective epithelial barrier
function in asthma. J Allergy Clin Immunol. 128:549–556. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aslam M, Ahmad N, Srivastava R and Hemmer
B: TNF-α induced NFκB signaling and p65 (RelA) overexpression
repress Cldn5 promoter in mouse brain endothelial cells. Cytokine.
57:269–275. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bruewer M, Luegering A, Kucharzik T,
Parkos CA, Madara JL, Hopkins AM and Nusrat A: Proinflammatory
cytokines disrupt epithelial barrier function by
apoptosis-independent mechanisms. J Immunol. 171:6164–6172. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cui W, Li LX, Sun CM, Wen Y, Zhou Y, Dong
YL and Liu P: Tumor necrosis factor alpha increases epithelial
barrier permeability by disrupting tight junctions in Caco-2 cells.
Braz J Med Biol Res. 43:330–337. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Coyne CB, Vanhook MK, Gambling TM, Carson
JL, Boucher RC and Johnson LG: Regulation of airway tight junctions
by proinflammatory cytokines. Mol Biol Cell. 13:3218–3234. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Deissler HL, Deissler H, Lang GK and Lang
GE: VEGF but not PIGF disturbs the barrier of retinal endothelial
cells. Exp Eye Res. 115:162–171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Antonetti DA, Barber AJ, Khin S, Lieth E,
Tarbell JM and Gardner TW: Penn State Retina Research Group:
Vascular permeability in experimental diabetes is associated with
reduced endothelial occludin content: Vascular endothelial growth
factor decreases occludin in retinal endothelial cells. Diabetes.
47:1953–1959. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deissler HL, Deissler H and Lang GE:
Actions of bevacizumab and ranibizumab on microvascular retinal
endothelial cells: Similarities and differences. Br J Ophthalmol.
96:1023–1028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sekiyama A, Gon Y, Terakado M, Takeshita
I, Kozu Y, Maruoka S, Matsumoto K and Hashimoto S: Glucocorticoids
enhance airway epithelial barrier integrity. Int Immunopharmacol.
12:350–357. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council: Committee for
the Update of the Guide for the Care and Use of Laboratory
AnimalsGuide for the Care and Use of Laboratory Animals. 8th.
Washington (DC), USA: National Academies Press; 2011
|
|
15
|
Temelkovski J, Hogan SP, Shepherd DP,
Foster PS and Kumar RK: An improved murine model of asthma:
Selective airway inflammation, epithelial lesions and increased
methacholine responsiveness following chronic exposure to
aerosolised allergen. Thorax. 53:849–856. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuksel H, Yilmaz O, Baytur YB and Ozbilgin
K: Prenatal administration of granulocyte-macrophage
colony-stimulating factor increases mesenchymal vascular
endothelial growth factor expression and maturation in fetal rat
lung. Exp Lung Res. 34:550–558. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lambrecht BN and Hammad H: Asthma: The
importance of dysregulated barrier immunity. Eur J Immunol.
43:3125–3137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holgate ST: Pathogenesis of asthma. Clin
Exp Allergy. 38:872–897. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holgate ST: The sentinel role of the
airway epithelium in asthma pathogenesis. Immunol Rev. 242:205–219.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Felinski EA, Cox AE, Phillips BE and
Antonetti DA: Glucocorticoids induce transactivation of tight
junction genes occludin and claudin-5 in retinal endothelial cells
via a novel cis-element. Exp Eye Res. 86:867–878. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Harhaj NS, Felinski EA, Wolpert EB,
Sundstrom JM, Gardner TW and Antonetti DA: VEGF activation of
protein kinase C stimulates occludin phosphorylation and
contributes to endothelial permeability. Invest Ophthalmol Vis Sci.
47:5106–5115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuksel H, Kose C, Yilmaz O, Ozbilgin K,
Degirmenci PB, Pinar E and Kirmaz C: Increased expression of tissue
vascular endothelial growth factor and foetal liver kinase-1
receptor in seasonal allergic rhinitis and relevance to asthma
component. Clin Exp Allergy. 37:1183–1188. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hardyman MA, Wilkinson E, Martin E,
Jayasekera NP, Blume C, Swindle EJ, Gozzard N, Holgate ST, Howarth
PH, Davies DE and Collins JE: TNF-α-mediated bronchial barrier
disruption and regulation by src-family kinase activation. J
Allergy Clin Immunol. 132:665–675. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamada H, Yoneda M, Inaguma S, Watanabe D,
Banno S, Yoshikawa K, Mizutani K, Iwaki M and Zako M: Infliximab
counteracts tumor necrosis factor-α-enhanced induction of matrix
metalloproteinases that degrade claudin and occludin in
non-pigmented ciliary epithelium. Biochem Pharmacol. 85:1770–1782.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arnett HA and Viney JL: Considerations for
the sensible use of rodent models of inflammatory disease in
predicting efficacy of new biological therapeutics in the clinic.
Adv Drug Deliv Rev. 59:1084–1092. 2007. View Article : Google Scholar : PubMed/NCBI
|