Introduction
Myocardial ischemia-reperfusion injury (IRI) is a
common pathophysiological event that occurs in a variety of
clinical settings, including cardiopulmonary resuscitation,
myocardial transplantation, trauma and shock (1,2). The
molecular mechanisms underlying myocardial IRI are complex,
specific details remain unclear. However, there is evidence to
suggest that inflammation plays an important role in myocardial IRI
(3,4). Therefore, suppressing inflammation may
be an effective strategy for the treatment of myocardial IRI.
Phosphoinositide 3-kinase (PI3K) is composed of a
catalytic subunit (p110) and a regulatory subunit (p85) (5); activation of the catalytic subunit
relies on p85 phosphorylation. The activation of p85 by
phosphorylation leads to the phosphorylation of protein kinase B
(Akt), which subsequently results in the production of
proinflammatory mediators, such as tumor necrosis factor (TNF)-α,
interleukin (IL)-6 and IL-1β (6).
However, further activation of PI3K signaling has been demonstrated
to significantly suppress inflammation in IRI (7).
Erythropoietin (EPO) is a glycoprotein cytokine that
is produced primarily in the kidneys (8). Previously, EPO has been widely used in
clinical practice due to its ability to stimulate erythropoiesis
(8,9). In addition to this well-known and
widely recognized effect, EPO, has been shown to significantly
attenuate inflammation in kidney and brain IRI through regulating
the PI3K/Akt signaling pathway (10,11).
However, the impact of EPO, and the associated PI3K/Akt signaling
pathway, on myocardial IRI has not, to the best of our knowledge,
been previously studied. Therefore, the aim of the present study
was to establish a rat model of IRI in order to investigate this
area further.
Materials and methods
Animals
A total of 18 male Sprague Dawley (SD) rats (6–8
weeks-old; weight, 200–250 g) were purchased from Hua Fukang
Experimental Animal Center (Beijing, China). The rats were housed
in a specific pathogen-free facility at 18–29°C on a 14 h light/10
h dark cycle. The rats were provided with standard laboratory chow
and water ad libitum. Following a minimum acclimatization
period of 7 days, the rats were randomly allocated to three groups
(n=6 per group): i) IRI-saline group (IRI-SAL), in which the rats
were administered intraperitoneal (IP) saline injections 24 h prior
to IRI induction; ii) IRI-EPO group, in which the rats were
administered EPO (5,000 U/kg, IP) 24 h prior to IRI induction; and
iii) sham-operated group (Sham), in which the rats were subjected
to the same surgical procedures without occlusion of the left
descending coronary artery. The dosage of EPO was selected based on
that of a previous study (12). The
study was approved by the Institutional Animal Care and Use
Committee at the West China Hospital of Sichuan University
(Chengdu, China).
IRI induction in the heart
Rats were anesthetized by an IP injection of 1%
pentobarbital sodium solution (6 ml/kg; Qiao Xing, Shanghai,
China), and their body temperature was regulated by a lamp (Pang
Kang, Shanghai, China) during the procedure. Myocardial IRI was
induced as follows. The rats were placed in the supine position and
an incision was made in the left side of the chest. The heart was
exposed and exteriorized. A slipknot (4-0 silk; Ai De, Anhui,
China) was placed around the left anterior descending coronary
artery. After 30 min of ischemia, the slipknot was released and the
myocardium was reperfused for 3 h. Rats in the Sham group underwent
the same surgical procedure as the other groups, with the exception
of coronary artery occlusion.
Histological analysis
After 3 h of reperfusion, the excised hearts were
fixed with 10% formalin, embedded in paraffin and stained with
hematoxylin and eosin. The hearts were visualized under a CX31
microscope (1,000x magnification; Olympus Corporation, Tokyo,
Japan). Myocardial IRI was scored in the most obvious part of the
lesion, according to previously published morphological criteria
(13) as follows: 0, no damage; 1
(mild), interstitial edema and localized necrosis; 2 (moderate),
widespread myocardial cell swelling and necrosis; 3 (severe),
necrosis with contraction bands and compressed capillaries; and 4
(highly severe), diffuse necrosis with contraction bands,
compressed capillaries and hemorrhage.
Western blot analysis
Myocardial samples (50 mg), were mechanically
homogenized in 1 ml hypotonic buffer (200 µl HEPES, pH 7.5; 25
mmol/l magnesium chloride; 5 mmol/l dithiothreitol; 5 mmol/l
phenylmethylsulfonyl fluoride; 2 mmol/l pepstatin A; 10 µg/ml
leupeptin; 5 mmol/l EDTA; 10 µg/ml aprotinin; and 100 µl
anti-phosphatases; Roche Diagnostics, Basel, Switzerland). Protein
concentrations were determined using a bicinchoninic assay kit (Bi
Yun-tian, Wuhan, China), according to the manufacturer's
instructions. Samples of 80 µg protein were resolved using 12%
SDS-PAGE, and transferred to polyvinylidene fluoride membranes (San
Yin-Tan, Beijing). The membranes were blocked with 5% non-fat skim
milk in tris-buffered saline solution containing Tween-20 (TBST; 10
mmol/l tris, pH 7.5; 140 mmol/l sodium chloride; 0.1% Tween-20) for
1 h at 37°C, washed, and incubated with the corresponding primary
antibody in TBST containing 3% bovine serum albumin (Gibco Life
Technologies, Carlsbad, CA, USA). The membranes were incubated for
12 hours with the following primary antibodies: Rabbit anti-EPOR
(1:1,000, #41346), rabbit anti-phosphorylated (p)-EPOR (1:500,
#BY-1424P), rabbit anti-p85 (1:1,000, #2883S), rabbit anti-p-p85
(1:500, #4292S), rabbit anti-Akt (1:1,000, #9037S) and rabbit
anti-p-Akt (1:500, #8475S) (Cell Signaling Technology, Inc.,
Danvers, MA, USA) at 4°C. The membranes were then washed
extensively with TBS, prior to being incubated with a horseradish
peroxidase-conjugated immunoglobulin G secondary antibody (1:4,000;
J010203; Jackson ImmunoResearch Laboratories, West Grove, PA, USA)
for 1 h at 37°C. β-actin (1:3000, #4301; Abmart, Shanghai, China)
was used for normalization of protein levels. The reactive bands
were visualized using the Enhanced Chemiluminescence-Plus reagent
(GE Healthcare, Piscataway, NJ, USA), according to the
manufacturer's instructions. The density of each reactive band was
quantified using the Labworks Image Acquisition platform, and its
related analytical software (UVP, LLC, Upland, CA, USA).
ELISA analysis
Levels of the inflammatory mediators, TNF-α, IL-6
and IL-1β, in the serum were quantified using an ELISA kit specific
to rats, according to the manufacturer's instructions (BioSource
International Inc., Camarillo, CA, USA).
Quantitative polymerase chain reaction
(PCR) analysis
Total RNA was isolated from the cardiac tissue using
TRIzol® reagent, according to the manufacturer's instructions
(Takara Bio, Inc., Otsu, Japan). In total, 4 µg RNA was reverse
transcribed into cDNA using the PrimeScript RT Master Mix (Takara
Bio, Inc.) as instructed. Quantitative PCR amplifications were
conducted using the ABI 7500 system (Applied Biosystems Life
Technologies, Foster City, CA, USA). PCR primers (Invitrogen Life
Technologies, Carlsbad, CA, USA) for all the analyzed genes are
shown in Table I. PCR was conducted
as follows: Initial denaturation at 95°C for 30 sec, followed by 40
cycles of denaturation at 95°C for 5 sec, annealing at 60°C for 34
sec and elongation at 95°C for 15 sec. The mRNA expression for each
gene was normalized against the values for β-actin, and the
relative expression levels were calculated using the
2−ΔΔCt method, as previously reported (14).
 | Table I.Primers used for quantitative
polymerase chain reaction analysis. |
Table I.
Primers used for quantitative
polymerase chain reaction analysis.
| Gene | Species | Sense strand
sequence | Anti-sense strand
sequence |
|---|
| TNF-α | Rat |
CTGAACTTCGGGGTGATCGG |
GGCTTGTCACTCGAATTTTGAGA |
| IL-1β | Rat |
AGCTTCCTTGTGCAAGTGTCT |
GACAGCCCAGGTCAAAGGTT |
| IL-6 | Rat |
CTGCAAGAGACTTCCATCCAG |
AGTGGTATAGACAGGTCTGTTGG |
| β-actin | Rat |
AGAGGGAAATCGTGCGTGAC |
CAATAGTGATGACCTGGCCGT |
Statistical analysis
SPSS software, version 12.0 (SPSS, Inc., Chicago,
IL, USA) was used to perform all statistical analyses. All data are
expressed as the mean ± standard error of the mean. All statistical
comparisons were performed using one-way analysis of variance
followed by Dunetts's test. In all cases, P<0.05 was considered
to indicate a statistically significant difference.
Results
EPO attenuates IRI-induced
pathological changes in the myocardial tissue
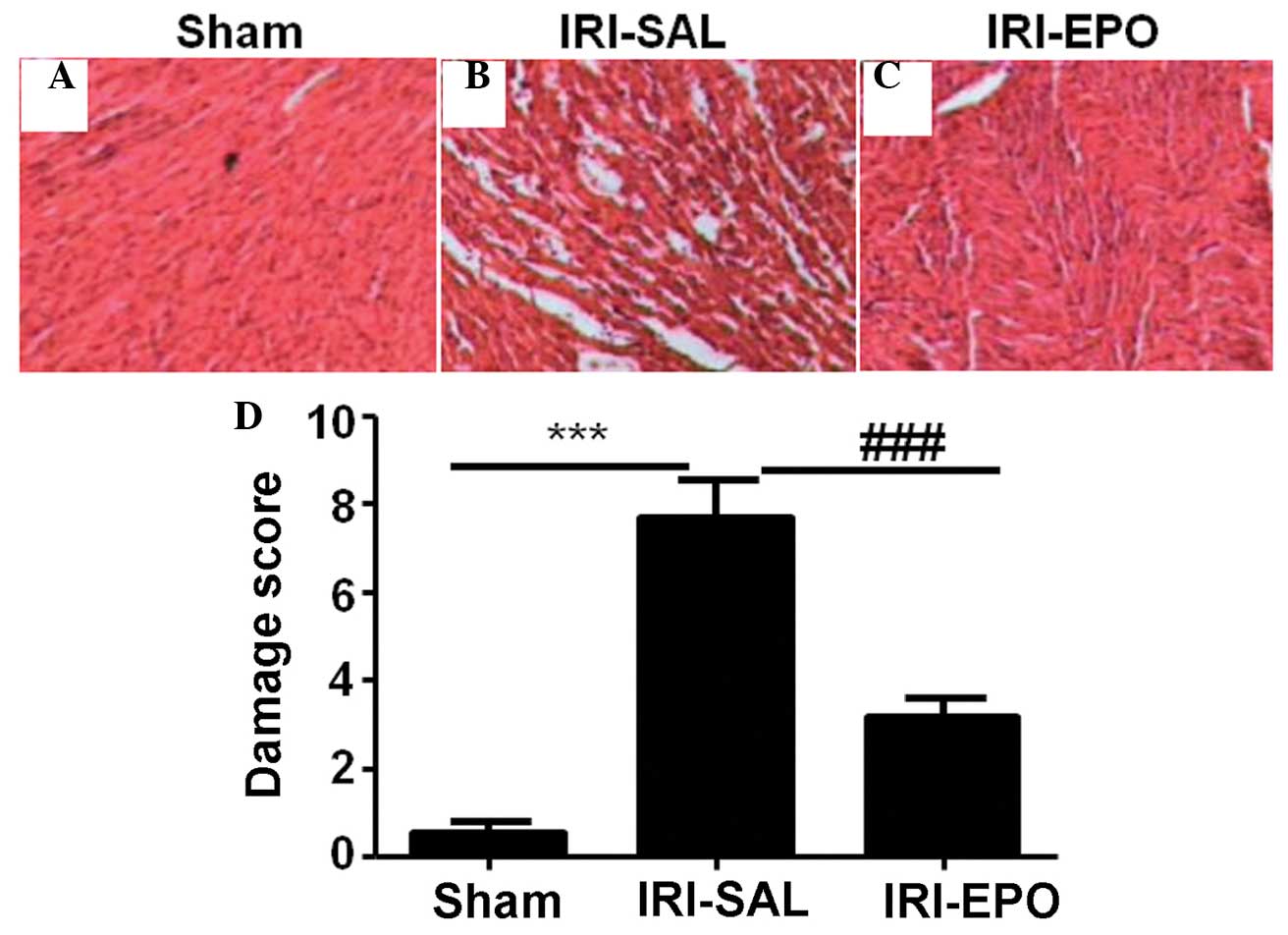
When compared with the rats in the Sham group, the
pathological changes in the myocardial tissues from the rats in the
IRI-SAL group were more marked (Fig. 1A
and B). The changes included atrophy of the myocardial fibers,
inflammatory cell infiltration, and coagulative and colliquative
necroses. By contrast, the myocardial tissues from the rats in the
IRI-EPO group (Fig. 1C) exhibited
fewer pathological changes compared with the IRI-SAL group.
Semi-quantitative assessment of the histological lesions revealed a
significantly higher score in the IRI-SAL-treated rats as compared
with the IRI-EPO-treated rats and Sham rats (Fig 1D).
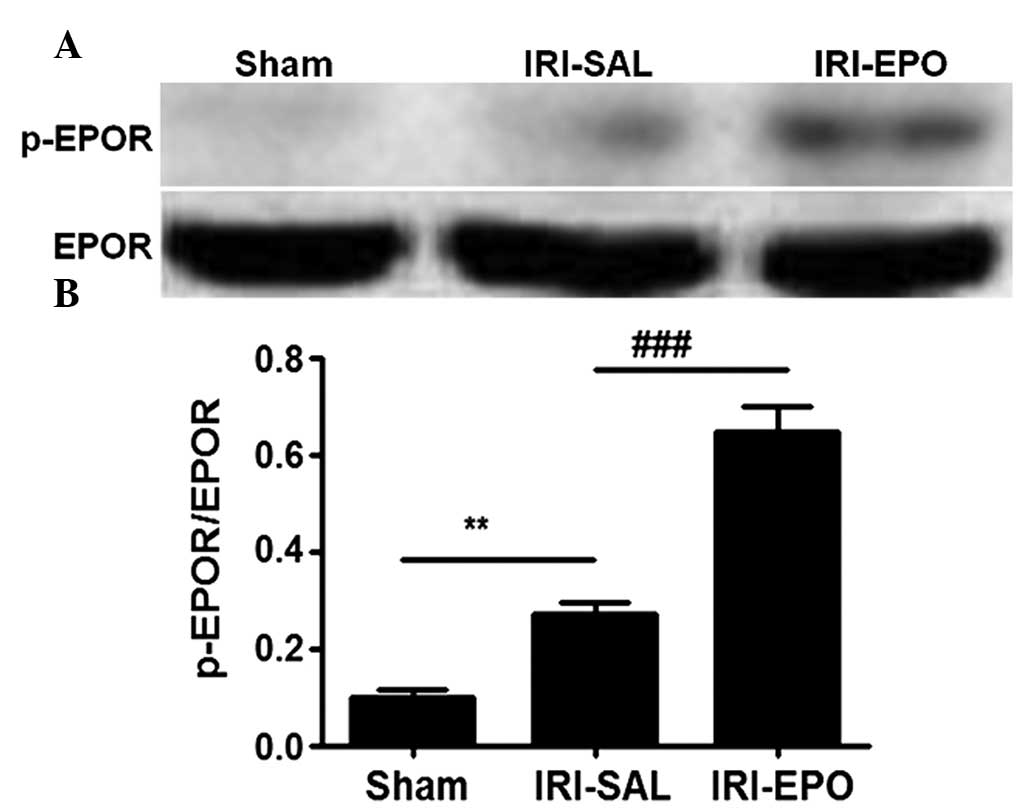
EPO promotes IRI-induced EPOR
activation
In order to address the mechanism underlying the
EPO-mediated protection of rat myocardial tissue from IRI, EPOR
expression and phosphorylation (activation) were examined. Neither
the IRI-SAL or IRI-EPO procedures revealed a marked impact on EPOR
expression (Fig. 2A, lower panel).
However, the amount of p-EPOR observed in the rat myocardial tissue
following IRI in the IRI-SAL group was significantly higher
compared with that in the Sham rats (Fig. 2A, upper panel and Fig. 2B). Administration of EPO slightly
promoted EPOR activation, as shown by the higher levels of p-EPOR
(40% higher) detected in the IRI-EPO rats when compared with the
IRI-SAL rats (Fig. 2B).
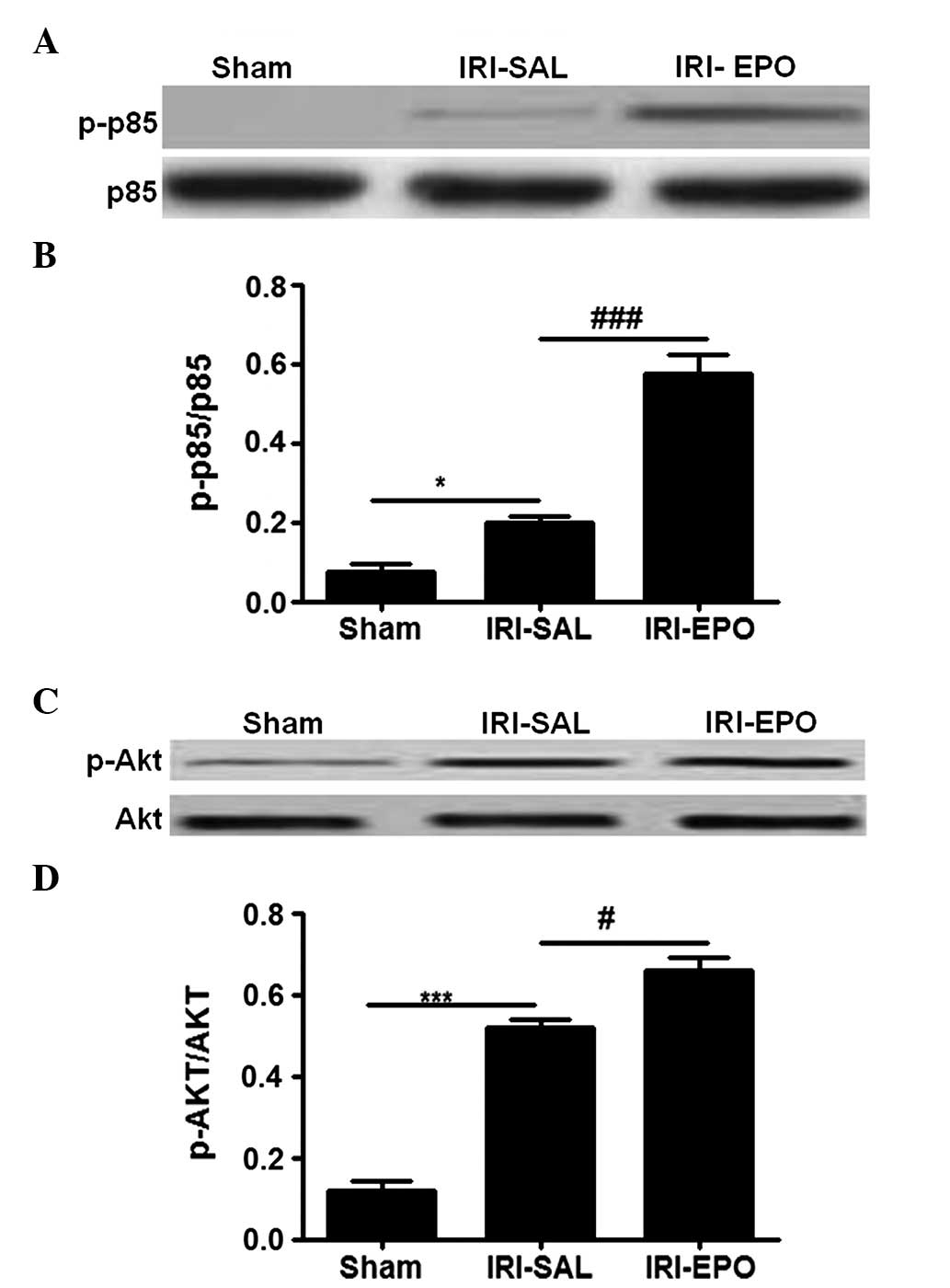
EPOR activation further activates
PI3K/Akt signaling
The PI3K/Akt pathway has been demonstrated to play
an important role in myocardial injury (15), while EPO is able to regulate PI3K/Akt
signaling (16). To investigate the
contribution of the PI3K/Akt pathway in the rat model, the activity
of the PI3K p85 regulatory subunit was examined. No statistically
significant difference in the total p85 level was detected among
the three groups of rats (Fig. 3A).
However, IRI was shown to induce PI3K p85 regulatory subunit
activation, as shown by the significantly higher levels of p-p85 in
the IRI-SAL rats when compared with the sham rats. In addition,
pretreatment with EPO promoted p85 activation, as manifested by the
significantly higher levels of p-p85 in the IRI-EPO rats when
compared with the IRI-SAL rats.
Since p85 activation provides signals to
phosphorylate Akt, Akt activity was examined. Similarly to p85,
there was no statistically significant difference in the total
levels of Akt among the three groups (Fig. 3C); however, the levels of p-Akt were
significantly higher in the IRI-SAL rats when compared with the
Sham rats. Furthermore, rats in the IRI-EPO group exhibited
significantly higher levels of p-Akt when compared with the IRI-SAL
rats (Fig. 3D). Thus, EPO
pretreatment increased PI3K p85 activity, which subsequently
increased Akt activation.
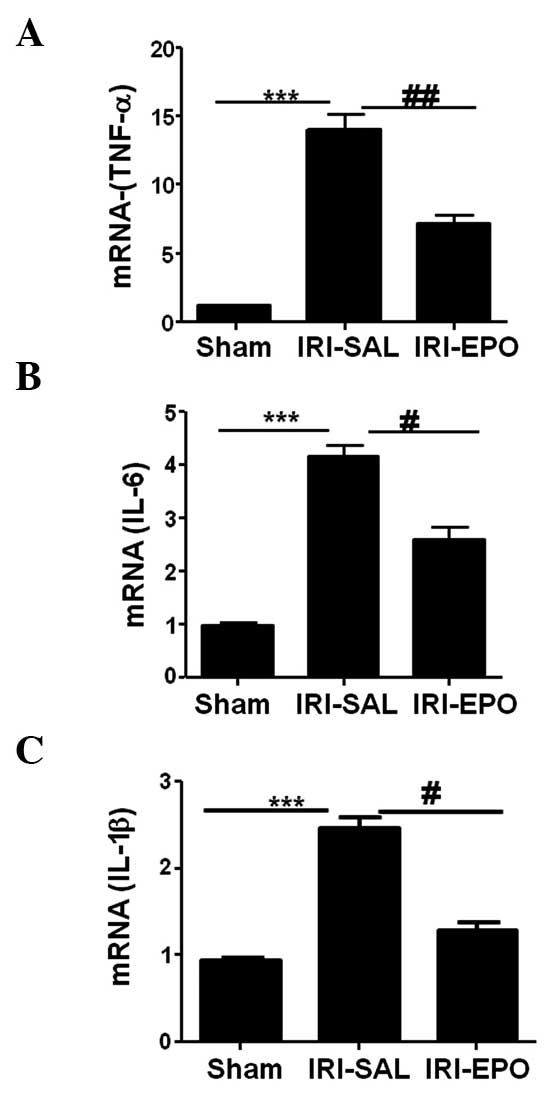
Activation of PI3K/Akt signaling
attenuates inflammatory cytokine expression
The present study also investigated whether the
activation of PI3K/Akt signaling attenuated the expression levels
of inflammatory cytokines in the heart. The expression levels of
TNF-α, IL-6 and IL-1β in the myocardial tissue following IRI were
selectively analyzed using quantitative PCR. IRI-SAL treatment was
shown to increase the expression of TNF-α by 13-fold (Fig. 4A), IL-6 by 3-fold (Fig. 4B) and IL-1β by 2.5-fold, as compared
with the expression levels in the Sham rats. Administration of EPO
inhibited IRI-induced TNF-α expression by 55% (Fig. 4A), IL-6 by 42% (Fig. 4B) and IL-1β by 30% (Fig. 4C) when compared with the expression
levels in the IRI-SAL group. Thus, the data showed that activation
of the PI3K/Akt signaling pathway by EPO significantly suppressed
IRI-induced inflammatory cytokine expression in the myocardial
tissue.
Activation of PI3K/Akt signaling
suppresses inflammatory cytokine secretion
The expression levels of TNF-α, IL-6 and IL-1β were
also analyzed in the serum following IRI. IRI-SAL treatment was
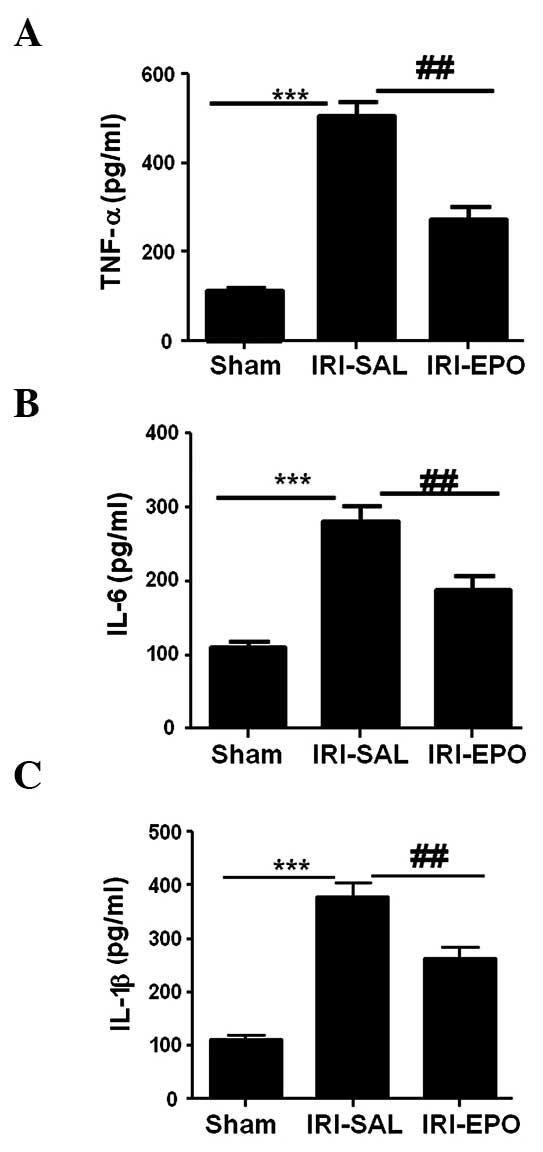
demonstrated to increase the secretion of TNF-α by 4-fold (Fig. 5A), IL-6 by 2-fold (Fig. 5B) and IL-1β by 3-fold, as compared
with the Sham rats. However, administration of EPO reduced the
IRI-induced TNF-α levels by 50% (Fig.
5A), IL-6 levels by 30% (Fig.
5B) and IL-1β levels by 25% (Fig.
5C) when compared with the expression levels in the IRI-SAL
group. Therefore, the data revealed that activation of the PI3K/Akt
signaling pathway by EPO significantly suppressed the levels of
IRI-induced inflammatory cytokines in the serum.
Discussion
EPO has been used for a number of years to promote
red blood cell production in various clinical settings (8,9). In the
present study, IRI was induced in rats by occluding the left
descending coronary artery for 30 min and subsequently subjecting
the heart to 3 h reperfusion. The effects of EPO on myocardial IRI
were subsequently investigated. Histological analyses demonstrated
significant pathological changes in the IRI group, as shown by
marked atrophy of the myocardial fibers, inflammatory cell
infiltration and coagulative and colliquative necroses.
Pretreatment of the rats with 5,000 U/kg EPO was
shown to significantly attenuate the pathological changes
associated with myocardial IRI. In agreement with the current
results, previous studies have suggested that treatment of rats
with EPO significantly decreases the pathological injury to the
heart tissue that is induced by myocardial IRI (12,17,18). The
present data indicate that EPO may be a novel agent for the
prevention of myocardial IRI in clinical practice.
In order to determine the molecular mechanism
through which EPO exerts protection against myocardial IRI in rats,
the present study investigated the effect of EPO on EPOR
activation. The results revealed that treatment with EPO did not
affect EPOR expression; however, IRI-SAL treatment led to increased
levels of p-EPOR. Pretreatment with EPO further increased EPOR
activation, as shown by the higher levels of p-EPOR in the IRI-EPO
rats when compared with the IRI-SAL rats. These observations may be
the result of negative feedback pathways, which function to limit
inflammatory responses and prevent a prolonged immune response
following severe tissue injury.
A number of studies have shown that EPO ameliorates
IRI in the kidneys and brain by suppressing inflammation (10,11).
Therefore, the present study investigated the downstream pathways
of EPOR activation that are associated with the regulation of
inflammation, namely the PI3K/Akt signaling pathway. IRI-SAL
treatment was shown to induce PI3K p85 regulatory subunit
activation, as manifested by the significantly higher levels of
p-p85 in the IRI-SAL rats when compared with the Sham rats. In
addition, pretreatment of the rats with EPO further promoted p85
activation following IRI.
These results prompted the examination of Akt
activation, since the phosphorylation of p85 can lead to Akt
activation (5,19,20).
Consistent with the aforementioned results, IRI-EPO treatment was
shown to significantly increase Akt activity, as demonstrated by
the higher levels of p-Akt expression in the IRI-EPO rats when
compared with the IRI-SAL rats. To demonstrate that activation of
the PI3K/Akt signaling pathway by EPO inhibited inflammatory
responses, the expression levels of TNF-α, IL-6 and IL-1β were
analyzed in the myocardium and serum following IRI. Rats pretreated
with EPO exhibited significantly lower levels of TNF-α, IL-6 and
IL-1β when compared with the IRI-SAL rats. Therefore, the current
data suggest that EPO enhances EPOR activation, which subsequently
leads to the activation of PI3K/Akt signaling and the suppression
of inflammatory responses. This suppression protected the
myocardium from IRI.
The duration of ischemia and the time point for
measurements following reperfusion are critical factors in
determining the severity of IRI in rats (21,22).
Published studies have applied numerous variations in experimental
conditions, including temperature and duration of ischemia.
Furthermore, study results may vary depending on the strain of rats
used (21–24). In the present study, Sprague-Dawley
rats were used, 30 min of ischemia was applied and 3 h after
reperfusion was selected as the time point for examining IRI. A
similar degree of myocardial injury severity was observed in the
current model, as compared with a previously published study
(12). As previously mentioned,
evidence for myocardial injury was strongly demonstrated by the
pathological changes and infiltration of inflammatory cells. These
data also support the hypothesis that IRI initiates a complex
cascade of events, which results in myocardial injury characterized
by histological changes and infiltration of inflammatory cells.
With regard to the ability of EPO pretreatment to
prevent myocardial IRI, it is worth considering that EPOR
activation by EPO may involve additional pathways other than
PI3K/Akt, such as the mitogen-activated protein kinase cascade
(24). Additional studies addressing
the other potential pathways involved in EPOR activation following
IRI are required to fully elucidate the mechanism of EPO.
In summary, the present study demonstrated that
pretreatment of rats with EPO can provide protection against
myocardial IRI. This protection is manifested through the
amelioration of pathological changes to the myocardium and the
suppression of inflammation. In the current study, EPO pretreatment
decreased myocardial IRI, which was associated with the activation
of EPOR, increased PI3K/Akt signaling activation and the inhibition
of inflammatory mediator production. Therefore, the results
indicated that EPO may be a potential therapy for the prevention of
myocardial IRI in clinical practice.
Acknowledgements
This study was supported by grants from the Natural
Sciences Fund of Hubei Province (no. 2012FFC115) and the Public
Health Fund of Hubei Province (no. QJX2012-49).
References
|
1
|
Chen K, Li G, Geng F, et al: Berberine
reduces ischemia/reperfusion-induced myocardial apoptosis via
activating AMPK and PI3K-Akt signaling in diabetic rats. Apoptosis.
19:946–957. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yan X, Qiu W, Jia B, et al: Myocardial
protection by interferon-γ late preconditioning during
cardiopulmonary bypass-associated myocardial ischemia-reperfusion
in pigs. Oncol Rep. 30:2145–2152. 2013.PubMed/NCBI
|
|
3
|
Lassaletta AD, Elmadhun NY, Zanetti AV, et
al: Rapamycin treatment of healthy pigs subjected to acute
myocardial ischemia-reperfusion injury attenuates cardiac functions
and increases myocardial necrosis. Ann Thorac Surg. 97:901–907.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Du X, Hu X and Wei J: Anti-inflammatory
effect of exendin-4 postconditioning during myocardial ischemia and
reperfusion. Mol Biol Rep. 41:3853–3857. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim HJ, Joe Y, Kong JS, et al: Carbon
monoxide protects against hepatic ischemia/reperfusion injury via
ROS-dependent Akt signaling and inhibition of glycogen synthase
kinase 3beta. Oxid Med Cell Longev. 2013:3064212013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hofmann BT and Jücker M: Activation of
PI3K/Akt signaling by n-terminal SH2 domain mutants of the p85α
regulatory subunit of PI3K is enhanced by deletion of its
c-terminal SH2 domain. Cell Signal. 24:1950–1954. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen L, Wei X, Hou Y, Liu X, et al:
Tetramethylpyrazine analogue CXC195 protects against cerebral
ischemia/reperfusion-induced apoptosis through PI3K/Akt/GSK3β
pathway in rats. Neurochem Int. 66:27–32. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Palis J: Primitive and definitive
erythropoiesis in mammals. Front Physiol. 5:32014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jelkmann W: Physiology and pharmacology of
erythropoietin. Transfus Med Hemother. 40:302–309. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang C, Zhao T, Lin M, et al: Helix B
surface peptide administered after insult of ischemia reperfusion
improved renal function, structure and apoptosis through beta
common receptor/erythropoietin receptor and PI3K/Akt pathway in a
murine model. Exp Biol Med (Maywood). 238:111–119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Q, Bian H, Li Y, Guo L, Tang Y and
Zhu H: Preconditioning with the traditional Chinese medicine
Huang-Lian-Jie-Du-Tang initiates HIF-1α-dependent neuroprotection
against cerebral ischemia in rats. J Ethnopharmacol. 154:443–452.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Xie W, Liu P, et al: Mechanism of
the cardioprotection of rhEPO pretreatment on suppressing the
inflammatory response in ischemia-reperfusion. Life Sci.
78:2255–2264. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meij JT, Sheikh F, Jimenez SK, et al:
Exacerbation of myocardial injury in transgenic mice overexpressing
FGF-2 is T cell dependent. Am J Physiol Heart Circ Physiol.
282:H547–H555. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang S, Lv JW, Yang P, et al: Loss of
dicer exacerbates cyclophosphamide-induced bladder overactivity by
enhancing purinergic signaling. Am J Pathol. 181:937–946. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu H, Guo X, Chu Y and Lu S: Heart
protective effects and mechanism of quercetin preconditioning on
anti-myocardial ischemia reperfusion (IR) injuries in rats. Genes.
545:149–155. 2014.
|
|
16
|
Jia Y, Mo SJ, Feng QQ, et al:
EPO-Dependent activation of PI3K/AKT/FOXO3a signalling mediates
neuroprotection in in vitro and in vivo models of Parkinson's
disease. J Mol Neurosci. 53:117–124. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jun JH, Jun NH, Shim JK, Shin EJ and Kwak
YL: Erythropoietin protects myocardium against ischemia-reperfusion
injury under moderate hyperglycemia. Eur J Pharmacol. 745:1–9.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu MJ, Chen YS, Huang HS and Ma MC:
Erythropoietin alleviates post-ischemic injury of rat hearts by
attenuating nitrosative stress. Life Sci. 90:776–784. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He W, Wang Z, Zhou Z, et al:
Lipopolysaccharide enhances Wnt5a expression through toll-like
receptor 4, myeloid differentiating factor 88, phosphatidylinositol
3-OH kinase/AKT and nuclear factor kappa B pathways in human dental
pulp stem cells. J Endod. 40:69–75. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J, Yao Y, Xiao F, et al:
Administration of dexamethasone protects mice against
ichemia/reperfusion induced renal injury by suppressing PI3K/AKT
signaling. Int J Clin Exp Pathol. 6:2366–2375. 2013.PubMed/NCBI
|
|
21
|
Wang J, Li L and Jiang H: c-Cbl inhbition:
A novel therapeutic approach for attenuating myocardial ischemia
and reperfusion injury. Int J Cardiol. 186:50–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lin Y, Chen L, Li W and Fang J: Role of
high-mobility group box-1 in myocardial ichemia/reperfusion injury
and the effect of ethyl pyruvate. Exp Ther Med. 9:1537–1541.
2015.PubMed/NCBI
|
|
23
|
Yao HC, Yang LJ, Han QF, et al:
Postconditioning with simvastatin decreases myocardial injury in
rats following acute myocardial ischemia. Exp Ther Med.
9:1166–1170. 2015.PubMed/NCBI
|
|
24
|
Rafiee P, Shi Y, Su J, et al:
Erythropoietin protects the infant heart against
ischemia-reperfusion injury by triggering multiple signaling
pathways. Basic Res Cardiol. 100:187–197. 2005. View Article : Google Scholar : PubMed/NCBI
|