Introduction
A positive correlation between tea consumption and
body bone mineral density among postmenopausal females has been
reported in numerous countries, such as Canada (1), the UK (2), Taiwan (3), the USA (4), Japan (5)
and Australia (6), which may evolve
the study of the treatment of osteoporosis. Further investigations
have verified that habitual tea consumption contributes to a lower
risk of hip fracture (7,8). As the most abundant catechin, which
constitutes almost 60% of the catechins in tea (9), (-)-epigallocatechin-3-gallate (EGCG)
has received widespread attention. EGCG is considered to be
pharmacologically active (10–12), and
play a role in preventing various diseases, including
osteoporosis.
EGCG has been reported to protect against bone loss
to prevent further deterioration of the bone microarchitecture in
ovariectomized or non-ovariectomized rats (13–16). In
addition, EGCG has been demonstrated to improve the bone
microstructure and quality in aged, orchidectomized rats (17). Rodriguez et al (18) found that a combination of
α-tricalcium phosphate particles and EGCG effectively stimulated
bone regeneration. Furthermore, in vitro tests revealed that
EGCG increased the formation of mineralized bone nodules by human
osteoblast-like cells (19). Chen
et al (20) indicated that
EGCG ranging between 1 and 10 µM exerted a stimulatory effect on
the osteogenesis of murine bone marrow mesenchymal stem cell (MSC)
line, D1, by detecting the mRNA expression levels of associated
osteogenic genes. By contrast, EGCG has also been shown to induce
mouse osteoclast-like cell apoptosis (21) and inhibit the formation of
osteoclasts (22). A previous study
(23) indicated that EGCG may be
used as a pro-osteogenic agent for stem cell-based therapy in the
treatment of osteoporosis, since EGCG promoted the osteogenic
differentiation of primary human bone marrow MSCs (hBMSCs). Thus,
EGCG was hypothesized to have the potential to promote
osteogenesis. However, the effect of EGCG alone on triggering the
onset of osteogenesis in MSCs has seldom been studied. In the
majority of cases, osteoinductive agents, such as osteoinductive
media or bioactive ceramics, have also been added to stimulate
osteogenesis. However, whether EGCG plays the role as an enhancer
or an inducer is yet to be answered.
In the present study, the individual effect of EGCG
on the osteogenic differentiation of primary hBMSCs, without other
additives, was investigated through examining cell proliferation,
alkaline phosphatase (ALP) activity and the expression of
associated osteogenic genes.
Materials and methods
Isolation and culture of primary
hBMSCs
hBMSCs were harvested from bone marrow extracts
obtained from a routine iliac bone graft procedure for the
reconstruction of bone defects following a traumatic tibial
fracture. The study was approved by the local Ethics Committee of
Guangxi Medical University (Nanning, China), and written informed
consent was obtained from the patient. The patient was healthy with
no specific metabolic or inherited diseases. The bone marrow tissue
was flushed out with culture medium of α-modified Eagle's medium
(α-MEM; Gibco Life Technologies, Carlsbad, CA, USA), supplemented
with 10% (v/v) fetal bovine serum (FBS; Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) and 1% (v/v) antibiotics (100 U/l
penicillin and 100 U/l streptomycin; Solarbio Science &
Technology Co. Ltd., Beijing, China). Following centrifugation (5
min at room temperature, 100 xg), the cells were suspended in
culture medium and maintained in a humidified atmosphere with 5%
CO2 at 37°C. Non-adherent cells were removed by changing
the culture medium every 3 days. When 80–90% confluence was
achieved following 7–10 days of culture, the primary cells were
ready for use in the subsequent experiments. The culture medium was
changed every 2–3 days.
EGCG treatment
EGCG, with a purity of ≥98% as determined by high
performance liquid chromatography, was purchased from Shanghai
Yuanye Biotechnology Co. Ltd. (Shanghai, China) and stored at 4°C.
Prior to the experiments, EGCG was dissolved in dimethyl sulfoxide
(DMSO; Solarbio Science & Technology Co. Ltd.) at a
concentration of 10 mM and stored at −20°C ready for use. The EGCG
stock was diluted with culture medium immediately prior to
treatment. Cells were treated with EGCG at various concentrations:
0 µM as a negative control and 2.5, 5 and 10 µM as experimental
groups. The concentration of DMSO was <0.1% in all the
experiments.
Cell proliferation assay
Effects of EGCG on cell proliferation were assessed
using the 3-(4,5-dimethylthiazol-2-y1)-3,5-diphenyltetrazolium
bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA) method. Cells were
digested with 0.25% trypsin/EDTA (Sigma-Aldrich), concentrated by
centrifugation at 100 × g for 5 min, resuspended in culture medium
and subsequently seeded into 24-well plates at a density of
5×103 cells/well. After 24 h of culture, the culture
medium was replaced with: i) Pure culture medium in the negative
control; ii) culture medium with EGCG at concentrations of 2.5, 5
and 10 µM in the experimental groups; and iii) osteogenesis-induced
culture medium (α-MEM containing 10% FBS, 50 µg/ml L-ascorbic acid,
10 mM β-glycerophosphate and 100 nM dexamethasone; Sigma-Aldrich)
in the positive control. At days 3, 7, 14 and 21, a solution of MTT
in phosphate-buffered saline (PBS; Sigma-Aldrich) was added to each
well to a final concentration of 5 mg/ml. Following incubation in a
5% CO2 incubator (Forma™ Series II 3110 Water-Jacketed;
Thermo Fisher Scientific, Waltham, MA, USA) at 37°C for 4 h, the
supernatant was discarded and 1 ml DMSO was added to dissolve the
formazan crystals. Following thorough and even mixing, samples of
200 µl were randomly extracted from each of three parallel wells
with the same culture medium three times and transferred to 96-well
plates; thus, all samples were performed in nonuplicate. The
absorbance value was measured at 570 nm with a microplate reader
(Multiskan™ GO Microplate Spectrophotometer; Thermo Fisher
Scientific, Vantaa, Finland), and the results are shown as units of
optical density absorbance values.
ALP activity assay
To examine the ALP activity of the hBMSCs, cells
were seeded into 24-well plates at a density of 5×103
cells/well with the different culture media. Following 3, 7, 14 and
21 days of culture, the cells were washed with PBS and lysed with
200 µl radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Shanghai, China), which was added to
phenylmethanesulfonyl fluoride to form a final concentration of 1
mM prior to the analysis of ALP activity. The total protein
concentration (mg/ml) and ALP activity levels (U/ml) were measured
with an enhanced bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology) and an ALP reagent kit (Nanjing
Jiancheng Bioengineering Research Institute, Nanjing, China),
respectively, according to the manufacturer's instructions. ALP
levels were normalized against the total protein content. All
samples were examined in triplicate.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) assay
RT-qPCR analysis was performed for the detection of
osteogenic gene expression levels in the cells cultured in six-well
plates from the three groups. Total RNA was extracted with an
additional purification step employing an RNA isolation kit
(Tiangen Biotech Co. Ltd., Beijing, China) according to the
manufacturer's instructions, on days 3, 7, 14 and 21. Subsequently,
1 mg total RNA was reverse transcribed into cDNA. Finally, an ABI
7300 Sequence Detection System (Applied Biosystems Life
Technologies, Foster City, CA, USA) was used to conduct RT-qPCR
with TaqMan Universal PCR Master Mix and gene-specific TaqMan PCR
primers (Applied Biosystems Life Technologies), which included
those for bone morphogenetic protein 2 (BMP2), runt-related
transcription factor 2 (RUNX2), ALP, bone sialoprotein (BSP),
osteocalcin (OCN), α-1 type I collagen (COL1A1) and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Gene expression
levels were normalized against those of GAPDH using the comparative
2−ΔΔCt method. The primers used in this experiment are
shown in Table I. All PCR assays
were conducted in triplicate.
 | Table I.Primers used for reverse transcription
quantitative polymerase chain reaction. |
Table I.
Primers used for reverse transcription
quantitative polymerase chain reaction.
| Gene name | Forward primer | Reverse primer |
|---|
| GAPDH |
5′-CTATAAATTGAGCCCGCAGC-3′ |
5′-GACCAAATCCGTTGACTCCG-3′ |
| BMP2 |
5′-TCCATGTGGACGCTCTTTCA-3′ |
5′-AGCAGCAACGCTAGAAGACA-3′ |
| RUNX2 |
5′-TGTCATGGCGGGTAACGATG-3′ |
5′-CCCTAAATCACTGAGGCGGT-3′ |
| ALP |
5′-CCAGGGCTGTAAGGACATCG-3′ |
5′-GCTCTTCCAGGTGTCAACGA-3′ |
| BSP |
5′-CAATCTGTGCCACTCACTGC-3′ |
5′-TGCCCTGAACTGGAAATCGTT-3′ |
| OCN |
5′-ACACTCCTCGCCCTATTGGC-3′ |
5′-CCATTGATACAGGTAGCGCCT-3′ |
| COL1A1 |
5′-GTTCAGCTTTGTGGACCTCCG-3′ |
5′-GCAGTTCTTGGTCTCGTCAC-3′ |
Immunohistochemical assay
After 7, 14 and 21 days of culture,
immunohistochemical staining for BMP2 was performed. Cells on the
coverslips were washed with PBS, rinsed with 0.01% Triton X-100
(Beyotime Institute of Biotechnology), washed thoroughly with PBS,
treated with 3% hydrogen peroxide (Wuhan Boster Biological
Technology Co. Ltd., Wuhan, China), washed again in PBS and blocked
with 3% bovine serum albumin. Following incubation with a primary
antibody against BMP2 (rabbit, anti-human; cat. no. BA0585-1; Wuhan
Boster Biological Technology, Ltd.) at a dilution of 1:200, a
secondary antibody (goat, anti-rabbit IgG; cat. no. BA1055) and
biotin-labeled horseradish peroxidase were added to the cells.
Following incubation with 3,3′-diaminobenzidine and counterstaining
with hematoxylin, the cells were air-dried and sealed with neutral
resin. The cells were subsequently examined and images were
captured using inverted phase contrast microscopy (IX71-F22PH;
Olympus Corporation, Tokyo, Japan).
Statistical analysis
Data are presented as the mean ± 2 standard
deviation. All data were evaluated by one-way analysis of variance,
whilst the least significance difference multiple comparisons test
was performed for further evaluation of the data. The software used
was SPSS (version 16.0; SPSS Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Cell proliferation assay
The effect of EGCG on the proliferation of hBMSCs
was examined by an MTT assay. hBMSCs were treated with different
concentrations of EGCG (0 µM in the negative control group, and
2.5, 5 and 10 µM in the experimental groups) or osteogenic-induced
culture medium in the positive control group. As shown in Fig. 1, the rate of cell proliferation at
day 3 following treatment with EGCG in the range of 0–10 µM was
comparable with that in the positive control group; therefore, the
drug was demonstrated to be non-toxic to the cells, which was in
agreement with the previous (pilot) study (23). Although no statistically significant
difference was observed between the groups on day 3, cell
proliferation was evidently promoted by 5 µM EGCG between days 7
and 14. On day 21, the number of cells in the positive control
group was significantly higher compared with the other groups,
which may be attributed to the limit of the cell culture area
(Fig. 1). Therefore, the results
indicated that EGCG facilitated the growth of hBMSCs, particularly
at a concentration of 5 µM.
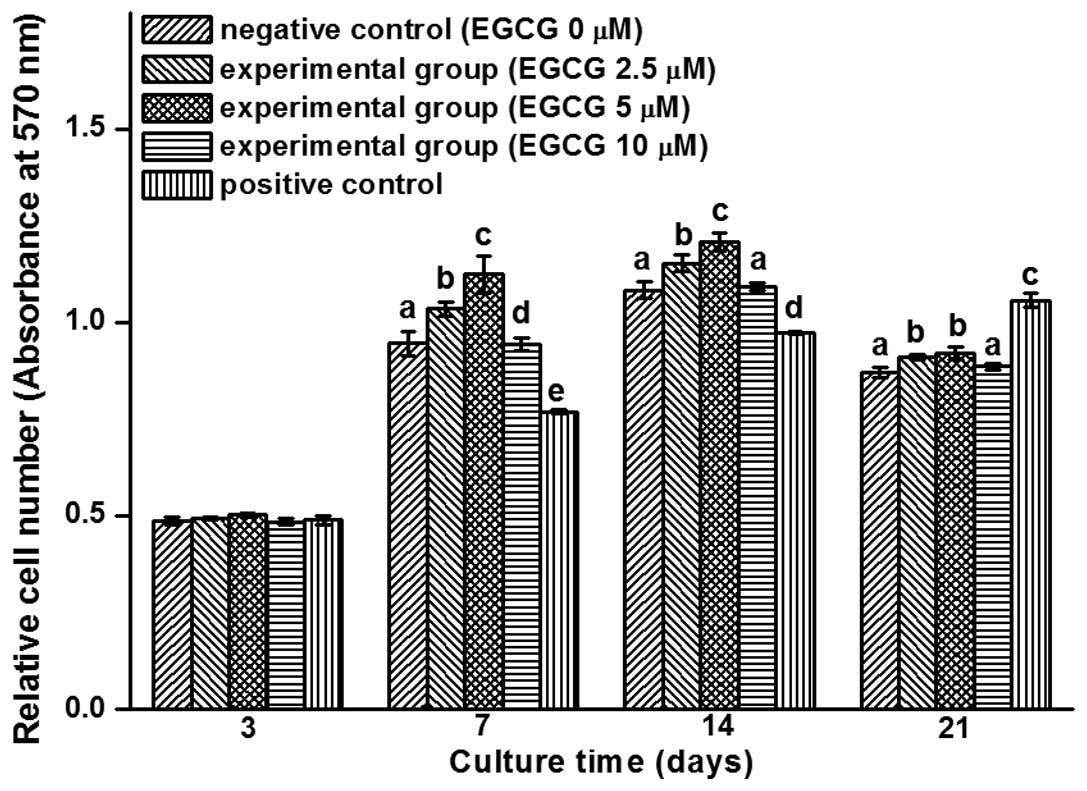 | Figure 1.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
analysis of cell proliferation in the human bone marrow mesenchymal
stem cells (hBMSCs) of the three groups (negative control, hBMSCs
were cultured with pure culture medium; experimental groups, cells
were treated with culture medium containing 2.5, 5 and 10 µM EGCG;
positive control, cells were cultured with osteogenesis-induced
culture medium). Cell proliferation in the experimental groups was
higher compared with the negative control group, while cell
proliferation in the positive control group increased over time. In
the experimental groups, cell proliferation with 5 µM EGCG was
higher compared with that of 2.5 and 10 µM. The bars with different
letters at the same time point are significantly different from
each other (P<0.05; n=9), and those with the same letter exhibit
no statistically significant difference. EGCG,
(-)-epigallocatechin-3-gallate. |
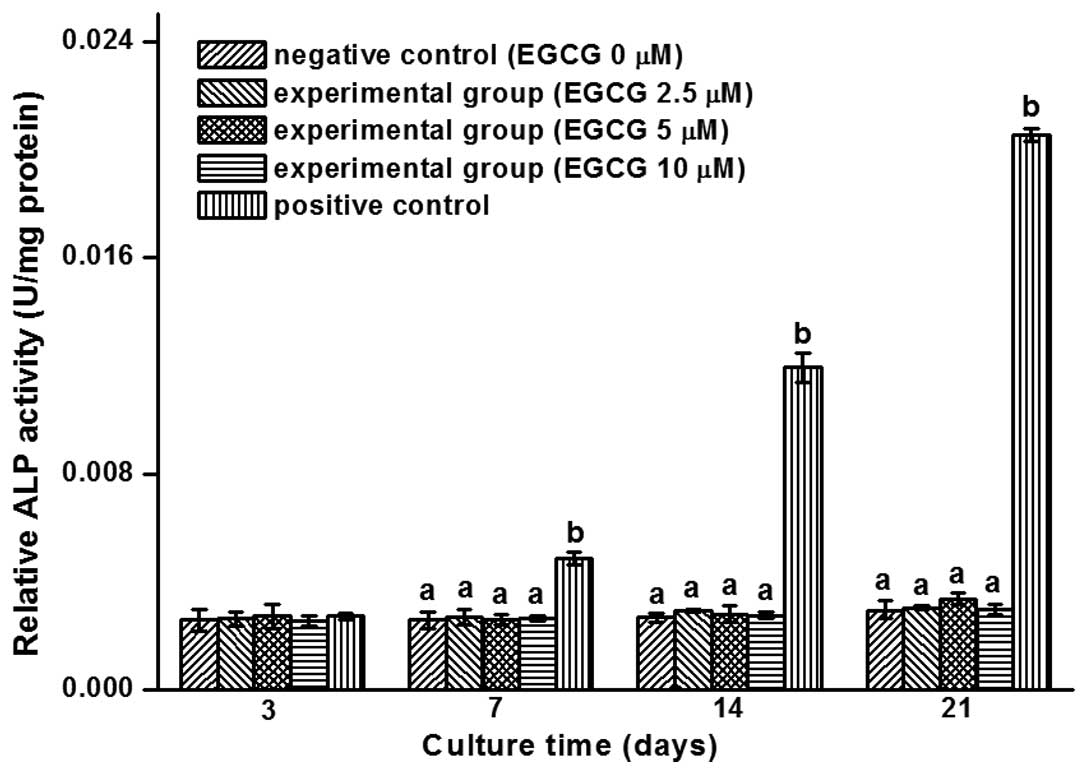
ALP activity assay
ALP is produced by osteoblasts and is hypothesized
to be involved in the degradation of inorganic pyrophosphates to
provide sufficient local phosphate or inorganic pyrophosphate for
the occurrence of mineralization (24). Commonly used as a marker of
osteogenesis, ALP activity is considered to reflect the degree of
osteogenic differentiation.
An ALP activity assay was used for the quantitative
assessment of the units of activity per mg protein in the three
groups, namely the negative control (hBMSCs cultured with pure
culture medium), experimental groups (cells treated with culture
medium containing 2.5, 5 and 10 µM EGCG respectively) and positive
control (cells cultured with osteogenesis-induced culture medium).
The histogram in Fig. 2 shows that
the ALP activity levels primarily remained unchanged with
increasing concentrations of EGCG; however, the activity level
increased significantly in the positive control over time.
RT-qPCR assay
The effect of EGCG on hBMSCs in the three groups
(negative control, experimental, positive control) was further
examined through gene expression analysis of BMP2, RUNX2, ALP, BSP,
OCN and COL1A1 following 3, 7, 14 and 21 days of culture.
Expression of the BMP2 gene was dose-dependently upregulated;
however, the expression levels of the other genes exhibited no
response to the different doses of EGCG, and the expression of the
COL1A1 gene following treatment with EGCG had a tendency to be
inferior to the controls. As a positive control, the expression of
all the detected genes in the positive group were superior to the
other groups (Fig. 3).
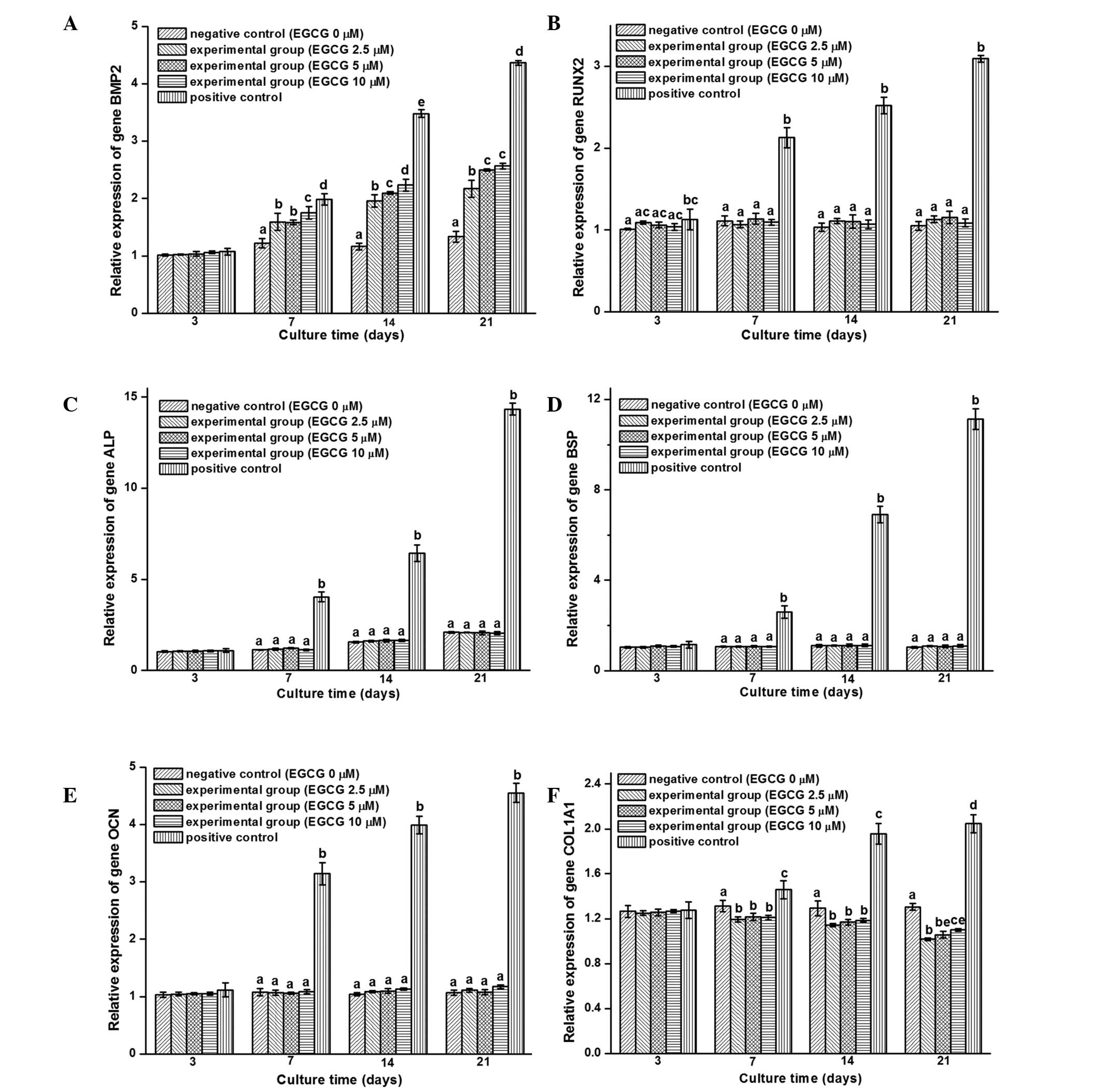 | Figure 3.Reverse transcription quantitative
polymerase chain reaction was used to analyze the expression of (A)
BMP2, (B) RUNX2, (C) ALP, (D) BSP, (E) OCN and (F) COL1A1
osteogenic genes in human bone marrow mesenchymal stem cells
(hBMSCs) cultured in the different groups for 3, 7, 14 and 21 days
(negative control, hBMSCs cultured with pure culture medium;
experimental, cells treated with culture medium containing 2.5, 5
and 10 µM EGCG; positive control, cells cultured with
osteogenesis-induced culture medium). In the positive control
group, all the detected genes were upregulated over time. When
compared with the negative control group, the expression of the
genes showed no change, with the exception of the BMP2 gene in the
experimental group that was upregulated, and the expression of the
COL1A1 gene, which exhibited a downward trend with time. The
minimum value was set to 1, and values are expressed as the mean ±
2 standard deviations. The bars with different letters at the same
time point are significantly different from each other (P<0.05;
n=3), and those with the same letter exhibit no statistically
significant difference. BMP2, bone morphogenetic protein 2; RUNX2,
runt-related transcription factor 2; ALP, alkaline phosphatase;
BSP, bone sialoprotein; OCN, osteocalcin; COL1A1, α-1 type I
collagen; EGCG, (-)-epigallocatechin-3-gallate. |
Immunohistochemical staining
Further investigation into the expression of
osteogenesis-associated proteins was performed using
immunocytochemistry with a specific antibody for BMP2 (Fig. 4). BMP2 is one of the members of the
transforming growth factor-β superfamily. The primary function of
BMP2 is the promotion of osteogenic differentiation (25–30). As
shown in Fig. 4, EGCG clearly
upregulated the protein expression levels of BMP2, with stronger
positive staining observed when compared with the control, which is
consistent with the RT-qPCR results. The staining results indicated
that the protein expression of BMP2 increased over time.
 | Figure 4.Immunohistochemical staining of bone
morphogenetic protein 2 expression in human bone marrow mesenchymal
stem cells (hBMSCs) of the three groups [negative control, hBMSCs
cultured with pure culture medium; experimental, cells treated with
culture medium containing 2.5, 5 and 10 µM
(-)-epigallocatechin-3-gallate (EGCG); positive control, cells
cultured with osteogenesis-induced culture medium]. (A–C) Staining
of hBMSCs in the negative control group on days 7, 14 and 21,
respectively. (D–F) Staining of hBMSCs in the 2.5 µM EGCG
experimental group on days 7, 14 and 21, respectively. (G–I)
Staining of hBMSCs in the 5 µM EGCG experimental group on days 7,
14 and 21, respectively. (J–L) Staining of hBMSCs in the 10 µM EGCG
experimental group on days 7, 14 and 21, respectively. (M–O)
Staining of hBMSCs in the positive control group on days 7, 14 and
21, respectively. The staining in the positive control group was
evidently greater, while that in the other groups was
dose-dependently upregulated. Magnification, x100. |
Discussion
EGCG, as the most abundant catechin and a major
bioactive component of green tea (31), has been reported by a number of
studies (19–23) to have an association with
osteogenesis. However, the association is dependent on an
osteogenic inducer, such as inductive media.
In the present study, the individual effect of EGCG
on osteogenesis was analyzed to clarify the role of EGCG in the
function of MSC differentiation. The results revealed that EGCG had
little effect on the osteogenic differentiation of MSCs, as
evidenced by only a marginal influence on ALP activity (Fig. 2) and the expression of osteogenic
genes, including RUNX2, ALP, BSP, OCN and COL1A1 (Fig. 3B–F). In accordance with the present
study, EGCG has a positive effect on proliferation and little
effect on the osteogenic differentiation without additional
osteoinductive agents. Kamon et al (32) also demonstrated that EGCG exerted a
suppressive effect on the differentiation of precursor cells of
osteoblasts, although EGCG had little effect on differentiated
osteoblasts. However, certain studies (20) have shown that EGCG can promote the
osteogenic differentiation of osteoblast precursors. These
observations indicate that EGCG functions more as an enhancer
rather than an inducer in osteogenic differentiation.
Notably, EGCG treatment was shown to upregulate BMP2
expression in a dose-dependent manner (Figs. 3A and 4). EGCG has been hypothesized to induce
osteogenesis at a relatively high concentration, but within the
recommended amount, which is more than the physiological level and
less than the detrimental level (33,34).
However, EGCG treatment alone is unable to produce sufficient
levels of BMP2 to trigger osteogenesis. By contrast, EGCG can
improve the facilitation of osteogenesis in the presence of
osteoinductive agents through coregulation of the BMP/Smad
signaling pathways (35–37). Thus, the effect of EGCG on
osteogenesis relies on the presence of an osteogenic inducer.
EGCG alone has a proliferative effect on the
proliferation of hMSCs. In particular, EGCG was shown to evidently
promote cell proliferation at a concentration of 5 µM. Furthermore,
a previous (pilot) study (23)
demonstrated that EGCG supports cell proliferation in the presence
of osteoinductive media. The study by Yagi et al (34) lent potent support for the present
study by elucidating that EGCG is capable of suppressing inducible
oxidative stress that may comprise cellular processes, including
cell proliferation. Therefore, EGCG is biocompatible and the
results support the possibility of clinical use.
In conclusion, EGCG alone has little effect on the
osteogenic differentiation of MSCs, and the catechin only plays a
role when combined with osteoinductive agents through the
upregulation of BMP2 expression. In addition, EGCG is able to
sustain cell proliferation, indicating the safety of the compound
for clinical application. Therefore, the present study indicated
that further development of EGCG as a pro-osteogenic agent should
be coupled with additional osteoinductive agents.
Acknowledgements
This study was financially supported by the National
Science and Technology Pillar Program of China (grant no.
2012BAI42G00), Guangxi Scientific Research and Technological
Development Foundation (grant no. Guikehe 14125008-2-14) and
Guangxi Science Fund for Distinguished Young Scholars (grant no.
2014GXNSFGA118006). This study has also been supported by the
Research Center for Regenerative Medicine of Guangxi Medical
University and Collaborative Innovation Center of Guangxi
Biological Medicine.
References
|
1
|
Hoover PA, Webber CE, Beaumont LF and
Blake JM: Postmenopausal bone mineral density: Relationship to
calcium intake, calcium absorption, residual estrogen, body
composition and physical activity. Can J Physiol Pharmacol.
74:911–917. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hegarty VM, May HM and Khaw KT: Tea
drinking and bone mineral density in older women. Am J Clin Nutr.
71:1003–1007. 2000.PubMed/NCBI
|
|
3
|
Wu CH, Yang YC, Yao WJ, et al:
Epidemiological evidence of increased bone mineral density in
habitual tea drinkers. Arch Intern Med. 162:1001–1006. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen Z, Pettinger MB, Ritenbaugh C, et al:
Habitual tea consumption and risk of osteoporosis: A prospective
study in the women's health initiative observational cohort. Am J
Epidemiol. 158:772–781. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Muraki S, Yamamoto S, Ishibashi H, et al:
Diet and lifestyle associated with increased bone mineral density:
Cross-sectional study of Japanese elderly women at an osteoporosis
outpatient clinic. J Orthop Sci. 12:317–320. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Devine A, Hodgson JM, Dick IM and Prince
RL: Tea drinking is associated with benefits on bone density in
older women. Am J Clin Nutr. 86:1243–1247. 2007.PubMed/NCBI
|
|
7
|
Johnell O, Gullberg B, Kanis JA, et al:
Risk factors for hip fracture in European women: The MEDOS study.
Mediterranean Osteoporosis Study. J Bone Miner Res. 10:1802–1815.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kanis J, Johnell O, Gullberg B, et al:
Risk factors for hip fracture in men from southern Europe: The
MEDOS study. Mediterranean Osteoporosis Study. Osteoporos Int.
9:45–54. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harbowy ME and Balentine DA: Tea
chemistry. Crit Rev Plant Sci. 16:415–480. 1997. View Article : Google Scholar
|
|
10
|
Chen CH, Ho ML, Chang JK, et al: Green tea
catechins enhance the expression of osteoprotegerin (OPG) in
pluripotent stem cells. J Orthop Surg Taiwan. 20:178–183. 2003.
|
|
11
|
Dulloo AG, Duret C, Rohrer D, et al:
Efficacy of a green tea extract rich in catechin polyphenols and
caffeine in increasing 24-h energy expenditure and fat oxidation in
humans. Am J Clin Nutr. 70:1040–1045. 1999.PubMed/NCBI
|
|
12
|
Stagg GV and Millin DJ: The nutritional
and therapeutic value of tea - a review. J Sci Food Agric.
26:1439–1459. 1975. View Article : Google Scholar
|
|
13
|
Shen CL, Wang P, Guerrieri J, et al:
Protective effect of green tea polyphenols on bone loss in
middle-aged female rats. Osteoporos Int. 19:979–990. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shen CL, Yeh JK, Stoecker BJ, et al: Green
tea polyphenols mitigate deterioration of bone microarchitecture in
middle-aged female rats. Bone. 44:684–690. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen CL, Yeh JK, Cao JJ, et al: Green tea
polyphenols mitigate bone loss of female rats in a chronic
inflammation-induced bone loss model. J Nutr Biochem. 21:968–974.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen CL, Yeh JK, Samathanam C, et al:
Green tea polyphenols attenuate deterioration of bone
microarchitecture in female rats with systemic chronic
inflammation. Osteoporos Int. 22:327–337. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen CL, Cao JJ, Dagda RY, et al:
Supplementation with green tea polyphenols improves bone
microstructure and quality in aged, orchidectomized rats. Calcif
Tissue Int. 88:455–463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rodriguez R, Kondo H, Nyan M, et al:
Implantation of green tea catechin α-tricalcium phosphate
combination enhances bone repair in rat skull defects. J Biomed
Mater Res B Appl Biomater. 98:263–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vali B, Rao LG and El-Sohemy A:
Epigallocatechin-3-gallate increases the formation of mineralized
bone nodules by human osteoblast-like cells. J Nutr Biochem.
18:341–347. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen CH, Ho ML, Chang JK, et al: Green tea
catechin enhances osteogenesis in a bone marrow mesenchymal stem
cell line. Osteoporos Int. 16:2039–2045. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakagawa H, Wachi M, Woo JT, et al: Fenton
reaction is primarily involved in a mechanism of
(-)-epigallocatechin-3-gallate to induce osteoclastic cell death.
Biochem Bioph Res Commun. 292:94–101. 2002. View Article : Google Scholar
|
|
22
|
Yun JH, Pang EK, Kim CS, et al: Inhibitory
effects of green tea polyphenol (-)-epigallocatechin gallate on the
expression of matrix metalloproteinase-9 and on the formation of
osteoclasts. J Periodontal Res. 39:300–307. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin P, Wu H, Xu G, et al:
Epigallocatechin-3-gallate (EGCG) as a pro-osteogenic agent to
enhance osteogenic differentiation of mesenchymal stem cells from
human bone marrow: An in vitro study. Cell Tissue Res. 356:381–390.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Na K, Sun BK, Woo DG, et al: Osteogenic
differentiation of rabbit mesenchymal stem cells in
thermo-reversible hydrogel constructs containing hydroxyapatite and
bone morphogenic protein-2 (BMP-2). Biomaterials. 28:2631–2637.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harris SE, Bonewald LF, Harris MA, et al:
Effects of transforming growth factor beta on bone nodule formation
and expression of bone morphogenetic protein 2, osteocalcin,
osteopontin, alkaline phosphatase and type I collagen mRNA in
long-term cultures of fetal rat calvarial osteoblasts. J Bone Miner
Res. 9:855–863. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hogan BL: Bone morphogenetic proteins:
Multifunctional regulators of vertebrate development. Genes Dev.
10:1580–1594. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Reddi AH: Bone and cartilage
differentiation. Curr Opin Genet Dev. 4:737–744. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wozney JM, Rosen V, Celeste AJ, et al:
Novel regulators of bone formation: Molecular clones and
activities. Science. 242:1528–1534. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wozney JM: The bone morphogenetic protein
family and osteogenesis. Mol Reprod Dev. 32:160–167. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yamaguchi A, Katagiri T, Ikeda T, et al:
Recombinant human bone morphogenetic protein-2 stimulates
osteoblastic maturation and inhibits myogenic differentiation in
vitro. J Cell Biol. 113:681–687. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Harborne JB and Williams CA: Advances in
flavonoid research since 1992. Phytochemistry. 55:481–504. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kamon M, Zhao R and Sakamoto K: Green tea
polyphenol (-)-epigallocatechin gallate suppressed the
differentiation of murine osteoblastic MC3T3-E1 cells. Cell Biol
Int. 34:109–116. 2009.PubMed/NCBI
|
|
33
|
Yang CH, Lin CY, Yang JH, et al:
Supplementary catechins attenuate cooking-oil-fumes-induced
oxidative stress in rat lung. Chin J Physiol. 52:151–159. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yagi H, Tan J and Tuan RS: Polyphenols
suppress hydrogen peroxide-induced oxidative stress in human
bone-marrow derived mesenchymal stem cells. J Cell Biochem.
114:1163–1173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hanai J, Chen LF, Kanno T, et al:
Interaction and functional cooperation of PEBP2/CBF with Smads.
Synergistic induction of the immunoglobulin germline C-alpha
promoter. J Biol Chem. 274:31577–31582. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Javed A, Barnes GL, Jasanya BO, et al:
Runt homology domain transcription factors (Runx, Cbfa and AML)
mediate repression of the bone sialoprotein promoter: Evidence for
promoter context-dependent activity of Cbfa proteins. Mol Cell
Biol. 21:2891–2905. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nishimura R, Hata K, Harris SE, et al:
Core-binding factor alpha 1 (Cbfa1) induces osteoblastic
differentiation of C2C12 cells without interactions with Smad1 and
Smad5. Bone. 31:303–312. 2002. View Article : Google Scholar : PubMed/NCBI
|