Introduction
Acute mountain sickness (AMS) is a medical
condition, resulting from acute exposure to high-altitude
environments (generally ≥2,500 m) and is characterized by headache,
dizziness, vomiting, fatigue and sleep disturbance (1,2). The
incidence of AMS ranges from 9 to 84% with variable altitudes,
ascent rates and latitudes (3). AMS
can progress to life-threatening high-altitude cerebral or
pulmonary edema (4), which seriously
impacts the health of the individual and reduces physical
capability. With increasing numbers of lowland individuals
travelling to high plateaus for a variety of reasons, AMS has
become a public health concern (5).
To date, the pathophysiology of AMS has yet to be fully elucidated,
although such a condition is believed to manifest neurological
complications linked to hypoxia (2).
It appears that, under the same acute hypoxic conditions, certain
lowland individuals are more susceptible to AMS than others
(6); therefore, it is possible that
genetic factors, such as the expression and variants of genes or
single nucleotide polymorphisms (SNPs), play a role in AMS
susceptibility.
Numerous genome-wide studies (7–11) and
other reports (12–14) of genetic variations in native Tibetan
highland populations have revealed that SNPs of endothelial PAS
domain protein 1 (EPAS1), which is also known as
hypoxia-inducible factor-2α (HIF-2α), have a strong and positive
selection in high-altitude hypoxia adaptation and are associated
with lower hemoglobin (Hb) concentrations (8). EPAS1 encodes the
oxygen-sensitive subunit of the HIF-2 transcription factor and
plays a role in regulating erythropoietin (15) and vascular endothelial growth factor
expression (16) under hypoxic
conditions. Genetic variants of EPAS1 that are shown to be
more common in highlanders may be worth investigating in lowlanders
with AMS, as the advantage conferred by the variants may be
beneficial under both acute and chronic hypoxia and represent the
migrational selection for AMS resistance in the population at high
altitude. Of the AMS genetic association studies that have been
performed, each has been limited to a few individuals or other gene
variants for AMS association (17,18).
Furthermore, little is known about the association between the
EPAS1 gene polymorphisms and the risk of AMS in lowlanders
following acute exposure to high-altitude hypoxia (19). Since the Han Chinese populate a
lowland region, they are considered to represent an appropriate
population for investigations into the association between genetic
factors and susceptibility or resistance to AMS.
In the present study, three tag SNPs [rs6756667,
rs13419896 and rs4953354; minor allele frequency (MAF), ≥5%] were
selected and genotyped in 603 unrelated Han Chinese men, who had
traveled to Lhasa by plane, using a matrix-assisted laser
desorption/ionization time-of-flight mass spectrometry (MALDI-TOF)
method. The physiological relevance of the three SNPs in the
development of AMS was then assessed, in order to enhance the
understanding of the molecular mechanisms associated with AMS risk
and facilitate the development of a novel strategy to control
AMS.
Materials and methods
Study population and AMS
evaluation
A total of 603 unrelated male Chinese Han subjects
were recruited from Chongqing, China. These individuals were
physical labors who traveled from Chengdu (500 m above sea level)
to Lhasa, Tibet (3,700 m above sea level) by plane. One week before
the trip, all the participants underwent health examinations in
Xinqiao Hospital (Chongqing, China); the participants were enrolled
in this study if they met the following inclusion criteria: i) Aged
between 18 and 45 years; ii) of Han Chinese ethnicity and with Han
Chinese parents; and iii) no history of high-altitude residency.
The exclusion criteria were as follows: i) Individuals with
diseases with similar clinical manifestations to AMS, such as
migraine and insomnia; and ii) individuals with respiratory or
cardiovascular system diseases, neuropsychosis, cerebrovascular
disease, cancer or dysfunction of the liver or kidneys. The
individuals were assessed using the current consensus of the Lake
Louise scoring system (LLss) (20)
18–24 h after their flight to Lhasa, which took place between June
21 and 25, 2012. The individuals with an LLss score of ≥3 points
and a headache were assigned to the AMS group (n=369, 61.2% of the
participants), while individuals with a score of <3 points or a
score of ≥3 points but no headache were assigned as a non-AMS group
(n=234, 38.8% of the participants).
Ethics statement
All subjects gave their informed consent for
inclusion prior to participation in the study. The study was
conducted in accordance with the Declaration of Helsinki, and the
protocol was approved by the Ethics Committee of Xinqiao Hospital,
Third Military Medical University (identification code, 2012014;
approved May 9, 2012; Chinese Clinical Trial Register no.
ChiCTR-OCS-11002213).
Data and blood sample collection
The demographic data were collected in Chongqing,
China, and physiological variables were measured within 18–24 h of
acute exposure to hypoxia (Lhasa, Tibet, China). Demographic data
[age, body mass index (BMI), tobacco smoking and alcohol
consumption] and AMS symptoms (dizziness, headache,
gastrointestinal disturbance, fatigue and difficulty sleeping) were
recorded using structured case report form questionnaires. The
physiological variables, including blood pressure (BP), arterial
oxygen saturation (SaO2) and heart rate (HR), were
measured once the participants had rested in the supine position
for 10 min. BP was recorded using a mercury sphygmomanometer, and
SaO2 and HR were monitored using a Finger-Pulse Oximeter
503 (Criticare Systems, Inc., Waukesha, WI, USA). BMI was
calculated following the measurement of the weight and height of
the participants.
For blood sampling, ~5 ml venous blood was collected
from each subject into EDTA tubes. Following collection, the sample
was centrifuged at 492 × g for 10 min at 4°C in order to separate
the blood cells and plasma. The plasma samples were stored at −80°C
until required for serological analysis, and the isolated blood
cells were immediately processed in order to extract DNA for
genotyping.
Selection of EPAS1 SNPs
The human EPAS1 gene (accession no.
NC_000002.12) is localized on chromosome 2 with nucleic acid
positions between 46,297,402 and 46,386,703 (National Center for
Biotechnology Information GenBank data; http://www.ncbi.nlm.nih.gov/genbank/). Genetic
variation data for the EPAS1 gene were obtained from the
HapMap project (21) for 82 healthy
Chinese Han Beijing (CHB) individuals (www.hapmap.org). In this study, three SNPs (rs6756667,
rs13419896 and rs4953354) with an MAF of ≥0.05 were selected.
EPAS1 rs13419896 and rs6756667 were selected based on two
genome-wide studies (8,14), while rs4953354 was selected according
to data from the Kunming Primate Research Center (Kunming, China)
(9). A maximally informative tag of
each SNP was selected using Tagger software (http://www.broad.mit.edu/mpg/haploview), which
captures all known common genetic variations in a gene based on a
linkage disequilibrium (LD) threshold of r2 ≥0.8 (22). The inverse of r2 represents
the ratio of sample size required to detect an indirect association
with a non-assayed SNP to direct an association at the same
statistical power.
DNA extraction and genotyping
Genomic DNA from each individual was extracted using
the Ezup Column Blood Genomic DNA Extraction kit (Shanghai Sangon
Biotechnology Co., Ltd., Shanghai, China) according to the
manufacturer's instructions and stored at −80°C until use. To
genotype these three SNPs, polymerase chain reaction primers were
first designed using Assay Design software (version 3.1; Sequenom,
Inc., San Diego, CA, USA) (Table I)
and then synthesized by Shanghai Sangon Biotechnology Co., Ltd.
MALDI-TOF (Sequenom Inc., San Diego, CA, USA) was subsequently
performed for genotyping. All genotyping was performed in a blinded
fashion without any knowledge of the identity of the individual,
and 10% of the DNA samples were repeatedly genotyped to monitor the
genotyping quality.
 | Table I.Primers used in this study to detect
EPAS1 SNPs. |
Table I.
Primers used in this study to detect
EPAS1 SNPs.
| SNP | Forward primer | Reverse primer | Sequencing
primer |
|---|
| rs6756667 |
5′-ACGTTGGATGTCGGAATCGACAGACTGGTG-3′ |
5′-ACGTTGGATGCTGACCAAGAGTTGATGCTG-3′ |
5′-AAACTGCTGTAAGGTGA-3′ |
| rs13419896 |
5′-ACGTTGGATGCTGAACCAGAGTCAGTAACC-3′ |
5′-ACGTTGGATGAACCCTTCCTGGTTGAGTAG-3′ |
5′-GAAAAGAGTCAGTAACCAATCCTAG-3′ |
| rs4953354 |
5′-ACGTTGGATGTCTGCCATTGTCTTGCATTG-3′ |
5′-ACGTTGGATGAAGAGGCGAAATGTGCAGAC-3′ |
5′-TGAAAGTATTTATGGAGCATATCT-3′ |
Statistical analysis
The statistical analyses were performed with SPSS
18.0 software (SPSS, Inc., Chicago, IL, USA). P≤0.05 was considered
to indicate a statistically significant difference, or a P-value of
<0.017 following Bonferroni correction for multiple testing. All
continuous variables are expressed as the mean ± standard
deviation. Differences between AMS and non-AMS subjects were
evaluated using the Student's t-test for continuous
variables and the χ2 test for categorical variables. The
Student's t-test was therefore performed to compare the
number of red blood cells (RBCs), hematocrit (HCT), Hb,
SaO2, HR and BP between different genotypes of AMS
individuals (a two-tailed P≤0.05 indicated statistical
significance). The allele frequencies of each SNP were calculated
by gene counting, and the genotype distribution of each SNP was
tested for Hardy-Weinberg equilibrium (HWE) using χ2
analyses. Three genetic models (allele-dose, dominant and
recessive) were subsequently used to assess association of these
SNPs with AMS risk. The association between the genotypes and the
AMS morbidity rate was calculated using χ2 analysis.
Odds ratios (ORs) with 95% confidence intervals (CIs) were adjusted
for age and BMI and calculated using multiple logistic regression
analyses to estimate the relative risk of AMS.
Results
Subject characteristics
The characteristics of this study cohort are
summarized in Table II. In brief,
there were 369 AMS cases (61.2%) and 234 non-AMS controls (38.8%)
in this unmatched, nested case-control study. All participants were
young Han Chinese men with a median age of 22.96±3.74 years (range,
18–45 years). No significant differences were found between the AMS
and non-AMS individuals in terms of age, BMI, tobacco smoking or
alcohol consumption (all P>0.05). The baseline characteristics
were thus considered comparable between the two groups. The
comparison of clinical characteristics between the two groups
revealed that the AMS individuals had a significantly higher HR and
lower SaO2 than the non-AMS individuals (both
P<0.01); however, the two groups showed similar levels of Hb,
RBCs and HCT, as well as systolic and diastolic BPs (all
P>0.05).
| Table II.Characteristics of the study subjects
(n=603). |
Table II.
Characteristics of the study subjects
(n=603).
|
Characteristics | AMS | Non-AMS | P-value |
|---|
| No. of patients, n
(%) | 369 (61.19) | 234 (38.81) |
|
| Baseline
demographic data |
|
|
|
|
Agea, yearsa | 23.18±3.85 | 22.64±3.56 | 0.088 |
|
BMIa, kg/m2 | 21.83±2.31 | 21.63±2.16 | 0.289 |
| Tobacco
smoking status, % |
|
| 0.964 |
|
Never | 36.05 | 33.18 |
|
|
Former | 59.01 | 61.14 |
|
|
Current |
4.94 |
5.68 |
|
| Alcohol
consumption, % |
|
| 0.102 |
|
Never | 31.43 | 25.58 |
|
|
Former | 22.00 | 18.60 |
|
|
Current | 46.57 | 55.82 |
|
| Clinical
indexesa |
|
|
|
| SBP,
mm/Hg | 118.95±12.14 | 117.61±10.66 | 0.107 |
| DBP,
mm/Hg | 78.47±10.01 | 78.70±10.14 | 0.781 |
| HR,
bpm | 86.29±12.61 | 83.49±12.60 | 0.008 |
|
SaO2, % | 87.93±3.38 | 88.91±2.79 |
<0.001 |
| Hb,
g/1 | 145.62±0.89 | 145.14±1.65 | 0.206 |
| RBCs,
x1012/1 | 4.56±0.03 | 4.61±0.05 | 0.893 |
| HCT,
% | 41.27±0.23 | 41.22±0.41 | 0.263 |
| LLss-related
symptoms, n (%) |
|
|
|
|
Headache | 369 (100.00) | 74 (31.62) |
|
|
Dizziness | 340 (92.14) | 102 (43.59) |
<0.001 |
|
Fatigue | 333 (90.24) | 84 (36.32) |
<0.001 |
|
Gastrointestinal symptoms | 123 (33.33) | 14 (5.60) |
<0.001 |
|
Difficulty sleeping | 303 (82.11) | 80 (34.19) |
<0.001 |
| LLssa | 5.05±1.89 | 1.56±1.17 |
<0.001 |
| Degree of AMS, n
(%) |
|
|
|
| Mild:
LLss score between 3 and 5 | 175 (47.43) |
|
|
| Severe:
LLss score ≥5 | 194 (52.57) |
|
|
Using the LLss, which included measurements of
headache, dizziness, gastrointestinal symptoms, difficulty sleeping
and fatigue (23), it was found that
the incidence of these five symptoms was significantly different
between the two groups, with the AMS group exhibiting significantly
higher incidences than the controls (all P<0.001), particularly
with regard to the incidence of headache. The total LLss score of
the AMS group was 5.05±1.89, which was significantly (P<0.001)
higher than that of the non-AMS group (1.56±1.17). Specifically,
there were 175 individuals (47.43%) with mild AMS (LLss, 3–5) and
194 individuals (52.57%) with severe AMS (LLss, ≥5).
Association of EPAS1 SNPs with AMS
risk
In this study, three EPAS1 SNPs were
genotyped in the individuals, and it was found that the overall MAF
of the three SNPs (including AMS and non-AMS subjects) was similar
to that reported in the HapMap III database (Tables III and IV). The distribution of these three SNP
genotypes was within HWE (P>0.05) between the AMS and non-AMS
individuals.
| Table III.Distribution of the three endothelial
PAS domain protein 1 tag SNPs among the subjects. |
Table III.
Distribution of the three endothelial
PAS domain protein 1 tag SNPs among the subjects.
|
|
|
| MAF, n (%) | Genotype, n
(%) | HWE P-value |
|---|
|
|
|
|
|
|
|---|
| SNPs | N | Minor allele | Subjects | Database | Wild | Heterozygous | Variant |
|---|
| rs6756667 | 603 | A | 13.3 |
5.6 | 467 (77.45) | 130 (21.56) | 6 (0.99) | 0.35 |
| rs13419896 | 602 | A | 31.5 | 27.8 | 281 (47.47) | 251 (43.40) | 60 (10.13) | 0.72 |
| rs4953354 | 602 | G | 10.0 | 13.3 | 451 (74.92) | 141 (23.42) | 10 (1.66) | 0.79 |
| Table IV.Association between endothelial PAS
domain protein 1 tag single nucleotide polymorphisms and the risk
of AMS. |
Table IV.
Association between endothelial PAS
domain protein 1 tag single nucleotide polymorphisms and the risk
of AMS.
| A, rs6756667 |
|
|
|
|
|
|---|
|
|---|
| Factor |
Genotype/allele | AMS, n (%) | Non-AMS, n (%) | OR (95% CI) | P-value |
|---|
| Genotype | GG | 301 (81.6) | 166 (70.9) |
| 0.002 |
|
| AG | 67 (18.2) | 63 (26.9) |
|
|
|
| AA | 1 (0.2) | 5 (2.2) |
|
|
| Allele | G | 669 (90.7) | 395 (84.4) | 1.701
(1.261–2.545) | 0.001 |
|
| A | 69 (9.3) | 73 (15.6) |
|
|
| Dominant model | GG | 301 (81.6) | 166 (70.9) | 1.815
(1.233–2.666) | 0.002 |
|
| AG+AA | 68 (18.4) | 68 (29.1) |
|
| Recessive
model | AG+GG | 368 (99.7) | 229 (97.9) | 8.035
(1.007–69.214) | 0.025 |
|
| AA | 1 (0.3) | 5 (2.1) |
|
|
|
| B, rs13419896 |
|
|
|
|
|
|
| Factor |
Genotype/allele | AMS, n (%) | Non-AMS, n (%) | OR (95% CI) | P-value |
|
| Genotype | AA | 34 (9.2) | 26 (11.1) |
| 0.452 |
|
| AG | 154 (42.0) | 105 (45.1) |
|
|
|
| GG | 179 (48.8) | 102 (43.8) |
|
|
| Allele | G | 512 (69.8) | 309 (66.3) | 1.172
(0.914–1.502) | 0.211 |
|
| A | 222 (30.2) | 157 (33.7) |
|
|
| Dominant model | GG | 179 (48.8) | 102 (43.8) | 1.222
(0.879–1.701) | 0.232 |
|
| AG+AA | 188 (51.2) | 131 (56.2) |
|
| Recessive
model | AG+GG | 333 (90.7) | 207 (88.8) | 1.230
(0.717–2.110) | 0.451 |
|
| AA | 34 (9.3) | 26 (11.2) |
|
|
| C, rs4953354 |
|
|
|
|
|
|
| Factor |
Genotype/allele | AMS, n (%) | Non-AMS, n (%) | OR (95% CI) | P-value |
|
| Genotype | AA | 276 (75.0) | 175 (74.8) |
| 0.372 |
|
| AG | 88 (23.9) | 53 (22.6) |
|
|
|
| GG | 4 (1.1) | 6 (2.6) |
|
|
| Allele | A | 640 (87.0) | 403 (86.1) | 1.075
(0.766–1.509) | 0.674 |
|
| G | 96 (13.0) | 65 (13.9) |
|
|
| Dominant model | AA | 301 (81.6) | 166 (70.9) | 1.011
(0.693–1.476) | 0.953 |
|
| AG+GG | 68 (18.4) | 68 (29.1) |
|
| Recessive
model | AG+AA | 364 (98.9) | 228 (97.4) | 2.395
(0.669–8.578) | 0.167 |
|
| GG | 4 (1.1) | 6 (2.6) |
|
|
Under an additive model and Bonferroni correction,
and following adjustments for age and BMI, it was found that the
SNP rs6756667 was associated with the risk of developing AMS
(Table IV). By contrast, no
statistically significant association was found between rs13419896
or rs4953354 and the risk of AMS in the allele-dose, dominant and
recessive models (all P>0.1).
The genotype frequencies of EPAS1 rs6756667
GG, AG and AA were 81.60% (n=301), 18.2% (n=67) and 0.3% (n=1),
respectively, in the AMS group and 70.9% (n=166), 26.9% (n=63) and
2.1% (n=5), respectively, in the non-AMS group. The distribution of
the overall genotype frequency was significantly different between
the AMS and non-AMS groups (P=0.002). The allele frequency of G
(AMS, 90.7% vs. non-AMS, 84.4%) and A (AMS, 9.3% vs. non-AMS,
15.6%) was also significantly different between these two groups.
The individuals carrying the rs6756667 G allele had a 1.701-fold
greater risk of developing AMS (OR, 1.701; 95% CI, 1.261–2.545;
P=0.001) compared with those carrying the rs6756667 A allele. The
rs6756667 GG homozygous genotype was also significantly associated
with elevated AMS risk compared with the rs6756667 homozygous AA
and heterozygous AG genotypes (OR, 1.815; 95% CI, 1.233–2.666;
P=0.002) following the dominant model analysis; however, the
recessive model analysis did not show a statistically significant
association between rs6756667 and the AMS risk (OR, 8.035; 95% CI,
1.233–2.666; P=0.025).
Association between EPAS1 rs6756667
and AMS severity
The individuals carrying the variant G allele had a
significantly higher rate of mild AMS when compared with those
carrying the A allele (P=0.004 and 0.018 for mild AMS using the
allele-dose and dominant models, respectively). For severe AMS, the
subjects carrying the variant G allele also exhibited a
significantly higher rate when compared with those carrying the A
allele (P=0.014, 0.008 and 0.041 for severe AMS using the
allele-dose, dominant and recessive models, respectively) (Table V).
| Table V.Association between the endothelial
PAS domain protein 1 rs6756667 single nucleotide polymorphism and
AMS severity. |
Table V.
Association between the endothelial
PAS domain protein 1 rs6756667 single nucleotide polymorphism and
AMS severity.
|
|
|
|
| P-value |
|---|
|
|
|
|
|
|
|---|
|
Allele/genotype | Mild AMS, n
(%) | Severe AMS, n
(%) | Non-AMS, n (%) | Pa | Pa |
|---|
| A | 34 (9.7) | 35 (9.0) | 73 (15.6) | – | – |
| G | 316 (90.3) | 353 (91.0) | 395 (84.4) | 0.004 | 0.014 |
| GG | 142 (81.1) | 159 (82.0) | 166 (70.9) | 0.018 | 0.008 |
| AG+AA | 33 (18.9) | 35 (18.0 | 68 (29.1) |
|
|
| AG+GG | 174 (99.4) | 194 (100.0) | 229 (97.9) | – | 0.041 |
| AA | 1 (0.6) | 0 (0.000) | 5 (2.1) |
|
|
Association between EPAS1 rs6756667
and clinicophysiological characteristics
The SNP rs6756667 was next assessed for associations
with certain clinicophysiological characteristics (RBCs, HCT, Hb,
SaO2, HR and BP) in the AMS group. A significant
association was found between the SNP rs6756667 and Hb (P=0.007),
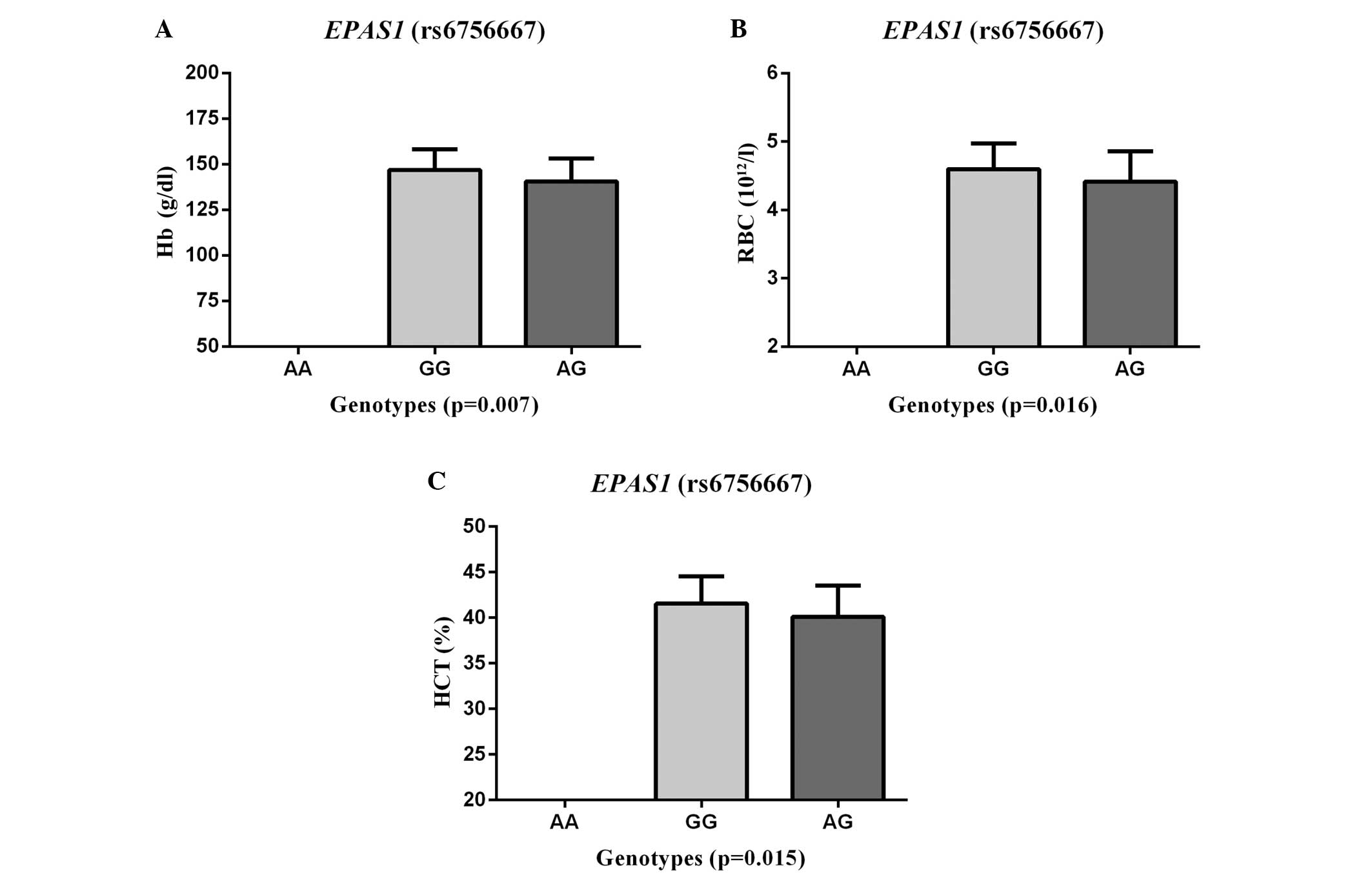
RBCs (P=0.016) and HCT (P=0.015) (Fig.
1), but there was no evident association with SaO2,
HR or BP (all P>0.05). The AMS patients with GG genotypes had
significantly higher levels of Hb, RBCs and HCT than those with the
rs6756667 AG heterozygous genotype.
Discussion
In this study, the association between three
EPAS1 SNPs and the risk of AMS in a male Chinese Han
population was assessed. It was found that the EPAS1
rs6756667 G allele was associated with a risk of developing AMS and
that the EPAS1 rs6756667 SNP was also associated with Hb,
RBC and HCT levels; AMS patients carrying the rs6756667 GG genotype
exhibited significantly higher Hb, RBC and HCT levels than those
with the AG genotype. These data indicate that EPAS1 SNPs
play a role in the physiological effects of AMS development in this
Chinese population; however, further study is required to evaluate
and confirm whether targeting EPAS1 could be a therapeutic
strategy to control acute hypoxia-related human diseases.
With regard to the association between AMS and
genetic variants in humans, this study was the first, to the best
of our knowledge, to investigate the potential clinical relevance
of EPAS1 rs6756667, rs13419896 and rs4953354 polymorphisms
in a large Chinese population. The major attraction of the use of
the LD tag SNPs was that it enabled a reduction in the genotyping
cost while maintaining a powerful detection system of non-assayed
causal SNPs. These tag SNPs are a subset of all variants within a
chromosome region for the study of an association with a disease.
In the current study, the approach to select SNPs was different
from that used in other studies, such as in the study by Buroker
et al (19). In their study,
one SNP (rs46441523), which contained five base pairs of nucleic
acids from the start of exon 6 that had a 78% frequency difference
between the highland Tibetans and the lowland Han Chinese, was
assessed.
EPAS1 has 16 exons extending over 90 kb on
chromosome 2, with a large first intron (50 kb) (24). EPAS1 rs6756667, rs13419896 and
rs4953354 are localized in the EPAS1 intron region and can
affect the binding to various transcription factors, as indicated
by bioinformatics analysis (http://www.gene-regulation.com/pub/programs.html;
http://motif.genome.ad.jp). In the present study,
the exonic SNPs were not selected, as a result of the lack of
information on SNPs in the exons of EPAS1 in Tibetans
(8–10). Furthermore, previous studies have
shown that the majority of the EPAS1 SNPs that are highly
differentiated between Tibetans on the Tibetan Plateau and
non-Tibetan CHB individuals only occur within the introns of
EPAS1 (9,25). At the protein level, exposure to
acute hypoxia induces rapid but short-lived responses, which often
occur as a result of the modification of existing proteins; by
contrast, chronic hypoxia induces responses that are delayed but
durable and that depend on changes in mRNA and protein expression
(26).
A major limitation of a traditional case-control
study is the potential for population stratification when
inappropriate patient-control matching occurs, such as the use of
healthy blood donors as the control group. To avoid a confounding
association, an unmatched, nested case-control study design was
selected (27); in the study, male
lowland Han individuals were enrolled and prospectively followed,
in order to determine whether those who had genetic variants had a
lesser or higher risk of AMS. It was found that the rs6756667 GG
genotype significantly increased the risk of developing AMS
compared with the AA and heterozygous AG genotypes. The present
results are consistent with those of a previous study involving
individuals of a Tibetan ethnicity, including Sherpas, that was
performed by Hanaoka et al (28). Previous genome-wide studies have
revealed that the rs6756667 polymorphism is significantly
associated with high-altitude adaptation in Tibetans (8,14).
Furthermore, Beall et al (8)
investigated 31 SNPs, including rs6756667, and the data showed a
significant association with the blood levels of Hb in Tibetans.
Results from Hanaoka et al (28) indicated that EPAS1 was under
selection for adaptation to the high-altitude life of the Sherpa
population. Bioinformatics analysis (http://www.gene-regulation.com/pub/programs.html;
http://motif.genome.ad.jp) has revealed that the
rs6756667 site contains a non-functional domain, but
cis-element activity regulates EPAS1 expression by
binding certain transcription factors (CREB, CRB-BP or v-ErbA);
however, additional functional studies are necessary to clarify
this theory. These findings underscore a potentially important role
of EPAS1 in influencing the development of life-threatening
high-altitude cerebral or pulmonary edema from AMS. The analysis of
patients with cerebral and pulmonary edema is likely to provide
further validation.
The present study additionally assessed
physiological data (Hb, HCT and RBC levels), in order to
investigate their association with EPAS1 rs6756667 SNP. At
sea level, Hb, HCT and RBCs are important physiological
requirements for the Hb-oxygen transport system to efficiently
carry and deliver oxygen to the tissues, cells and molecules of the
organism (29). The ascent to high
altitude is associated with a reduction in the atmospheric
O2 level; therefore, individuals ascending to high
altitude undergo the physiological stimulus of systemic hypoxia
(27), which evokes an adaptive
(homeostatic) response. EPAS1 plays a central role in the
response to hypoxia (30), and
hypoxia-sensing genes, including the EPAS1 pathways, are the
key regulators of RBC production (erythropoiesis), development,
energy metabolism, vasculogenesis, iron metabolism, cardiopulmonary
regulation and tumor promotion in mammals (13,31). In
the present study, it was found that the EPAS1 rs6756667 SNP
was significantly associated with levels of Hb, RBCs and HCT in the
AMS group, indicating that EPAS1 SNPs are associated with
AMS development and that GG homozygotes may have an advantage over
heterozygotes during this process. A previous study showed that
EPAS1 was associated with certain clinical phenotypes in
high-altitude illness (19). In the
study, the EPAS1 rs46441523 (C/G) SNP was assessed for an
association with age, RBCs, HCT, Hb, SaO2, HR and
systolic/diastolic BP among the chronic mountain sickness (CMS) and
AMS groups. An association was found between age and CMS, but no
association was found among the difference parameters and AMS
(19). The results may thus be
complicated by different sites and subjects (including males and
females).
The present study does have certain limitations.
First, only three, and not all, of the EPAS1 tag SNPs were
selected. Furthermore, no verification study was performed using
another set of samples, and the effect of the SNP that was
positively associated with AMS on the expression or function of
EPAS1 protein was not assessed. In addition, the complexity
and difficulties in obtaining additional blood samples did not
enable the plasma index to be determined in all subjects at high
altitude, and the subjects were randomly selected for the routine
blood examination (such as levels of Hb, RBCs and HCT). Another
limitation was that the proportion of individuals with the AA
genotype in the database was very low (5.6%); therefore, patients
with the rs6756667 AA homozygous genotype were not included for the
analysis of physiological features (Hb, RBCs and HCT) in the AMS
population in the present study. Despite the limited power of the
clinical association study, however, the results have shown for the
first time, to the best of our knowledge, the presence of an
association between the rs6756667 polymorphism and the risk of
developing AMS in this male Chinese Han population.
Acknowledgements
This study was supported in part by a grant from the
Special Health Research Project, Chinese Ministry of Health (grant
no. 201002012). The authors would like to thank all the individuals
that participated in the study and are grateful to Dr En-Hao Zhang,
Dr Xiao-Han Ding, Dr Ming-Yue Rao and Mr. Can Chen of the research
team for their continued support of this study. The authors would
additionally like to thank Medjaden Bioscience Ltd. for
assisting in the preparation of this manuscript.
References
|
1
|
Basnyat B and Murdoch DR: High-altitude
illness. Lancet. 361:1967–1974. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
MacInnis MJ, Wang P, Koehle MS and Rupert
JL: The genetics of altitude tolerance: The evidence for inherited
susceptibility to acute mountain sickness. J Occup Environ Med.
53:159–168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barry PW and Pollard AJ: Altitude illness.
BMJ. 326:915–919. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartsch P and Swenson ER: Clinical
practice: Acute high-altitude illnesses. N Engl J Med.
368:2294–2302. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schommer K, Bartsch P and Maeder MB: High
Altitude Illness. Notarzt. 29:175–183. 2013.(In German).
|
|
6
|
Schneider M, Bernasch D, Weymann J, Holle
R and Bartsch P: Acute mountain sickness: Influence of
susceptibility, preexposure, and ascent rate. Med Sci Sports Exerc.
34:1886–1891. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bigham A, Bauchet M, Pinto D, Mao X, Akey
JM, Mei R, Scherer SW, Julian CG, Wilson MJ, López Herráez D, et
al: Identifying signatures of natural selection in Tibetan and
Andean populations using dense genome scan data. PLoS Genet.
6:e10011162010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Beall CM, Cavalleri GL, Deng L, Elston RC,
Gao Y, Knight J, Li C, Li JC, Liang Y, McCormack M, et al: Natural
selection on EPAS1 (HIF2alpha) associated with low hemoglobin
concentration in Tibetan highlanders. Proc Natl Acad Sci USA.
107:11459–11464. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng Y, Yang ZH, Zhang H, Cui C, Qi X, Luo
X, Tao X, Wu T, Ouzhuluobu Basang, et al: Genetic variations in
Tibetan populations and high-altitude adaptation at the Himalayas.
Mol Biol Evol. 28:1075–1081. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simonson TS, Yang YZ, Huff CD, Yun H, Qin
G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, Jorde LB, et al:
Genetic evidence for high-altitude adaptation in Tibet. Science.
329:72–75. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yi X, Liang Y, Huerta-Sanchez E, Jin X,
Cuo ZX, Pool JE, Xu X, Jiang H, Vinckenbosch N, Korneliussen TS, et
al: Sequencing of 50 human exomes reveals adaptation to high
altitude. Science. 329:75–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ji LD, Qiu YQ, Xu J, Irwin DM, Tam SC,
Tang NL and Zhang YP: Genetic adaptation of the hypoxia-inducible
factor pathway to oxygen pressure among eurasian human populations.
Mol Biol Evol. 29:3359–3370. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Majmundar AJ, Wong WJ and Simon MC:
Hypoxia-inducible factors and the response to hypoxic stress. Mol
Cell. 40:294–309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu S, Li S, Yang Y, Tan J, Lou H, Jin W,
Yang L, Pan X, Wang J, Shen Y, et al: A genome-wide search for
signals of high-altitude adaptation in Tibetans. Mol Biol Evol.
28:1003–1011. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Percy MJ, Furlow PW, Lucas GS, Li X,
Lappin TR, McMullin MF and Lee FS: A gain-of-function mutation in
the HIF2A gene in familial erythrocytosis. N Engl J Med.
358:162–168. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bougatef F, Quemener C, Kellouche S, Naïmi
B, Podgorniak MP, Millot G, Gabison EE, Calvo F, Dosquet C, Lebbé
C, et al: EMMPRIN promotes angiogenesis through hypoxia-inducible
factor-2alpha-mediated regulation of soluble VEGF isoforms and
their receptor VEGFR-2. Blood. 114:5547–5556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Buroker NE, Ning XH, Zhou ZN, Li K, Cen
WJ, Wu XF, Zhu WZ, Scott CR and Chen SH: AKT3, ANGPTL4, eNOS3 and
VEGFA associations with high altitude sickness in Han and Tibetan
Chinese at the Qinghai-Tibetan Plateau. Int J Hematol. 96:200–213.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding H, Liu Q, Hua M, Ding M, Du H, Zhang
W, Li Z and Zhang J: Polymorphisms of hypoxia-related genes in
subjects susceptible to acute mountain sickness. Respiration.
81:236–241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Buroker NE, Ning XH, Zhou ZN, Li K, Cen
WJ, Wu XF, Zhu WZ, Scott CR and Chen SH: EPAS1 and EGLN1
associations with high altitude sickness in Han and Tibetan Chinese
at the Qinghai-Tibetan Plateau. Blood Cells Mol Dis. 49:67–73.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roach RC, Bärtsch P, Oelz O and Hachett
PLake Louise AMS Scoring Consensus Committee: The Lake Louise acute
mountain sickness scoring systemSutton JR, Houston CS and Coates G:
Hypoxia and Molecular Medicine. Queen City Printers; Burlington,
VT: pp. 272–274. 1993
|
|
21
|
International HapMap Consortium, . The
International HapMap Project. Nature. 426:789–796. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barrett JC, Fry B, Maller J and Daly MJ:
Haploview: Analysis and visualization of LD and haplotype maps.
Bioinformatics. 21:263–265. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Imray C, Booth A, Wright A and Bradwell A:
Acute altitude illnesses. Brit Med J. 343:d49432011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian H, McKnight SL and Russell DW:
Endothelial PAS domain protein 1 (EPAS1), a transcription factor
selectively expressed in endothelial cells. Genes Dev. 11:72–82.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang BB, Zhang YB, Zhang F, Lin H, Wang X,
Wan N, Ye Z, Weng H, Zhang L, Li X, et al: On the origin of
Tibetans and their genetic basis in adapting high-altitude
environments. PLoS One. 6:e1700222011.
|
|
26
|
Prabhakar NR and Semenza GL: Adaptive and
maladaptive cardiorespiratory responses to continuous and
intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2.
Physiol Rev. 92:967–1003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Desai RJ, Glynn RJ and Gagne JJ:
Performance of disease risk score matching in nested case-control
studies. Pharmacoepidemiol Drug Saf. 23 (Suppl):1482014.
|
|
28
|
Hanaoka M, Droma Y, Basnyat B, Ito M,
Kobayashi N, Katsuyama Y, Kubo K and Ota M: Genetic variants in
EPAS1 contribute to adaptation to high-altitude hypoxia in Sherpas.
PLoS One. 7:e505662012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jensen FB: The dual roles of red blood
cells in tissue oxygen delivery: Oxygen carriers and regulators of
local blood flow. J Exp Biol. 212:3387–3393. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hirota K and Semenza GL: Regulation of
angiogenesis by hypoxia-inducible factor 1. Crit Rev Oncol Hematol.
59:15–26. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Semenza GL: Hypoxia-inducible factors:
Mediators of cancer progression and targets for cancer therapy.
Trends Pharmacol Sci. 33:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|