Introduction
Pancreatic cancer has an extremely poor survival
rate; it ranks as the fourth most common cause of cancer-related
mortality and has a five-year survival rate of ~5% (1,2). Surgery
is effectively the only potential treatment for its eradication and
is suitable for only 10–20% of surgical candidates. In the majority
of patients, the opportunity to carry out surgery is lost for
reasons such as metastatic potential and delayed diagnosis
(1,3). The lack of a novel prognostic biomarker
is the most important reason for the poor outcome of patients with
pancreatic cancer. Therefore, the identification of efficient
prognostic biomarkers is urgently required.
The pathogenesis of pancreatic cancer has not been
completely elucidated. It has been found to be associated with
factors including the activation of oncogenes, dysfunction of tumor
suppressor genes, and aberrant activation of signal transduction
pathways (4). β-catenin, which is an
important effector in the Wnt signaling pathway, has an important
role in cell-cell adhesion (5).
Membrane-associated proteins that are formed by the binding of
E-cadherin to β-catenin are involved in the regulation and
provision of cellular adhesion (6).
In addition, cell motility, including metastasis and invasion, is
also regulated by membrane-associated adhesion proteins. In
association with the extracellular matrix and actin cytoskeleton,
membrane-associated adhesion proteins play crucial roles in
numerous biological signal transduction pathways (7). Dysfunction of β-catenin and E-cadherin
has been observed in various types of tumor, including breast
cancer, colorectal cancer and pancreatic cancer (7–9).
Ezrin-radixin-moesin-binding phosphoprotein-50
(EBP50, also known as NHERF1) is a 55-kDa phosphoprotein, which is
a member of the family of PDZ scaffolding proteins (10). The main site of localization of EBP50
is at the apical plasma membrane in human epithelial tissues
(11). The loss of normal apical
membrane expression of EBP50 and/or its distribution to the
cytoplasm and nuclear overexpression have been observed in several
types of human cancer (10). EBP50
has been indicated to be a tumor suppressor in several types of
tumor (10). Previous studies
conducted by the authors of the present study have demonstrated
that the downregulation of EBP50 expression promotes the growth of
gastric and pancreatic cancer cells (12,13), and
that the overexpression of EBP50 regulates the apoptosis of
pancreatic cancer cells by decreasing the expression levels of
Bcl-2 (11).
In the present study, the aim was to explore the
expression of EBP50 in pancreatic cancer tissue, the effect the
overexpression of EBP50 played in pancreatic cancer cell growth and
invasion and the underlying mechanism. The pBK-CMV-HA-EBP50 plasmid
was used to upregulate EBP50 expression in pancreatic cancer cells
and identify whether β-catenin/E-cadherin is directly targeted by
its effects on the growth and invasion of pancreatic cancer
cells.
Materials and methods
Human tissue specimens
Pancreatic cancer specimens were obtained from 120
patients of Renmin Hospital of Wuhan University (Wuhan, China) from
2003 to 2013 who were diagnosed with pancreatic cancer and had not
received any preoperative chemotherapy or radiotherapy. The
patients comprised 54 females and 66 males, aged from 25–82 years.
A total of 60 surgical samples of pancreatic cancer and matched
non-tumor tissues were collected for analysis by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
World Health Organization criteria were used for the diagnosis of
pancreatic cancer and the pathological tumor-node-metastasis (pTNM)
staging system was applied to define the tumor stage and
clinicopathological classification. Permission for the study was
granted by the Ethics Committee of Wuhan University, acquired
consent was obtained from every patient and the study was performed
according to the principles of the Declaration of Helsinki.
Immunohistochemistry
Followed the manufacturer's instructions,
immunohistochemical (IHC) staining of paraffinized sections was
performed with anti-EBP50 polyclonal rabbit antibody (1:800;
NB-300-536; Novus, Saint Charles, MO, USA). First, the pancreatic
cancer tissues were fixed in 10% buffered formalin, then embedded
in paraffin. After cutting into 3-µm sections and deparaffinizing
in xylene, the tissues were rehydrated in several descending
ethanol concentrations, incubated in 0.03% hydrogen peroxide for 10
min and then incubated in 10 mM sodium citrate buffer for 15 min
for antigen retrieval. Initial blocking was followed by incubation
with the anti-EBP50 antibody overnight at 4°C, three-washes with
phosphate-buffered saline (PBS)-Tween-20 (T) and a second blocking
step followed by incubation for 1 h at 37°C with goat anti-rabbit
IgG-PerCP-Cy5.5 biotinylated secondary antibody (1:400; sc-45101;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Finally, the
sections were washed with PBS-T, blocked again, then incubated with
diaminobenzidine (DAB) chromogen (DAB kit; Fujian Maixin Biological
Technology Ltd., Fujian, China) and counterstained with hematoxylin
prior to mounting. Isotype-matched irrelevant antibody was
substituted for the primary antibody to act as a negative
control.
Scoring of IHC
All stained tissue specimens were scored separately
by two pathologists, who were blinded to the clinical or
clinicopathological features of the specimens. The slides were
scanned at low magnification (x100 objective) and confirmed under
high magnification (x200 objective). The percentage of positive
stained cells in 10 representative microscopic fields was evaluated
as: 0, <5%; 1, 5–25%; 2, 25–50%; and 3, >50%. The intensity
of staining was scored as: 0 (none), 1+ (mild), 2+ (moderate), or
3+ (intense) according to a previous study (13,14).
RT-qPCR
Total RNA was extracted from fresh pancreatic cancer
tissues and paired adjacent non-tumorous samples using TRIzol
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA).
According to the manufacturer's instructions, the Reverse
Transcription system kit (Invitrogen Life Technologies) was used to
reverse-transcribe the RNA to first-strand complementary DNA
(cDNA). Corresponding levels of β-actin and EBP50 mRNA were
detected by RT-qPCR using the 7500 Real-Time PCR System (Applied
Biosystems Life Technologies, Foster City, CA, USA). The PCR was
run for 95°C for 3 min, followed by 40 cycles of 95°C for 3 sec and
60°C for 30 sec. β-actin was used as a normalization control for
EBP50 mRNA. The 2−ΔΔCt method was used to quantify the
relative levels of gene expression and each sample was analyzed in
triplicate. The qPCR primers sequences were as follows: EBP50,
forward: 5′-AGG AGT GCC TGA GTA GTC GCC AGT CAC CTG GGT CTG AGG GGC
CGA CGTC-3′ and reverse: 3′-TCA GGC ACT CCT GCT TTC TTG ACC GGA CCG
AAC CTG ATC A-5′; β-actin, forward: 5′-GTG ACG TTG ACA TCC G-3′ and
reverse: 5′-GAG CGT TTG TTG TAC CT-3′.
Cell lines and transient
transfection
The human pancreatic cancer cell lines PANC-1 and
SW1990 were obtained from the Cell Bank of the Shanghai Institutes
for Biological Sciences (Shanghai, China). Cells were maintained in
HyClone™RPMI-1640 medium with 10% fetal calf serum (Gibco-BRL,
Grand Island, NY, USA) and were cultured at 37°C with 5%
CO2. The pBK-CMV-HA empty vector was obtained from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA) and Dr Randy Hall from
Emory University (Atlanta, GA, USA) provided the pBK-CMV-HA-EBP50
plasmid. Cells were seeded in 6-well plates at a concentration of
2×105 cells/well and cultured in medium without
antibiotics for 24 h prior to transfection. Lipofectamine® 2000
(Invitrogen Life Technologies) was used to conduct the transfection
in accordance with the manufacturer's instructions. Cells were
transiently transfected with pBK-CMV-HA-EBP50, pBK-CMV-HA or
negative control (NC). After 24 h incubation at 37°C and 5%
CO2, the cells were trypsinized and reseeded into a
12-well plate and the medium was replaced with fresh normal culture
medium with G418 solution (Gibco-BRL) to select the stable
transfected cell clones. The transfection efficiency of each cell
clone was examined by western blot analysis (9,11).
Cell proliferation assay
The analysis of cell proliferation and viability was
performed using a cell-counting colorimetric assay (CCK-8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan). As recommended by
the manufacturer, three independent experiments were conducted for
each set of conditions. After washing with ice-cold PBS twice, the
cells were collected by trypsinization, and then seeded on a
96-well plate at a final density of 5×103 cells/well for
counting. A CCK-8 kit was used to assess the cell viability at 24,
48 and 72 h. The absorbance at 450 nm was measured using a plate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Colony formation assay
The colony formation assay was used to detect the
anchorage-independent growth of the transfected cells. The cells
were plated in a 6-well plate at a final density of
1×104 cells/well with Dulbecco's modified Eagle's medium
(DMEM) agarose medium with fetal bovine serum (FBS) and SeaPlaque™
agarose (all from Gibco-BRL). After 30 min at room temperature, the
plate was hardened and then incubated at 37°C with 5%
CO2. When the cell colonies were formed with a size
>0.1 mm, phase contrast microscopy was used to calculate and
photograph the result.
Cell invasion assay
Cell invasion assays were performed in Transwell
chambers (24-well insert, pore size 8 µm; BD Biosciences, Bedford,
MA, USA). The Transwell with a filter membrane having 8.0-mm pores
was inserted into a 24-well plate (bottom chamber). After being
suspended in serum-free medium, cells were seeded onto the top
chamber in 400 µl serum-free medium. Simultaneously, 800 µl DMEM
containing 10% FBS was added to the lower chambers of the Transwell
plate to act as a chemoattractant. A wet cotton-tipped swab was
used to remove the non-invaded cells from the upper surface. After
fixation with formalin, the chambers were stained with crystal
violet for 30 min. The number of cells that penetrated to the lower
surface of the filter was counted and images were captured
(magnification, x100) in five random fields. The data were
presented as a percentage of the invaded cells in the control and
each experiment was repeated three times.
Western blot analysis
Cells were washed with PBS then collected and lysed
on ice in RIPA buffer (Thermo Fisher Scientific, Inc., Rockford,
IL, USA). The BCA Protein assay kit (Pierce Biotechnology, Inc.,
Rockford, IL, USA) was used to measure the protein concentration.
The protein fractions were denatured at 100°C by suspension in
loading buffer. Total proteins were separated equally on 10%
SDS-PAGE gels and electro-transferred to nitrocellulose membranes
for 2 h at 4°C. Then the membranes were blocked in 5% fat-free milk
in TBS-T buffer at room temperature for 2 h. Rabbit anti-EBP50
antibody (1:1,000; Abcam, Cambridge, MA, USA), rabbit
anti-β-catenin (1:1,000; Abcam), mouse anti-E-cadherin (1:1,000;
Abcam) and mouse anti-β-actin (1:1,000; Abcam) primary antibodies
were added and the membranes were incubated overnight at 4°C. The
secondary horseradish peroxidase-conjugated anti-rabbit antibodies
(Sigma-Aldrich, St. Louis, MO, USA) were then incubated with the
membranes for 1 h at 4°C. The bands were detected by enhanced
chemiluminescence (ECL; Amersham Pharmacia Biotech, Piscataway, NJ,
USA) and quantified by densitometry using UN-SCAN-IT software (Silk
Scientific Corp., Orem, UT, USA). β-actin was used as an internal
control.
Results
Expression of EBP50 is decreased in
pancreatic cancer tissues
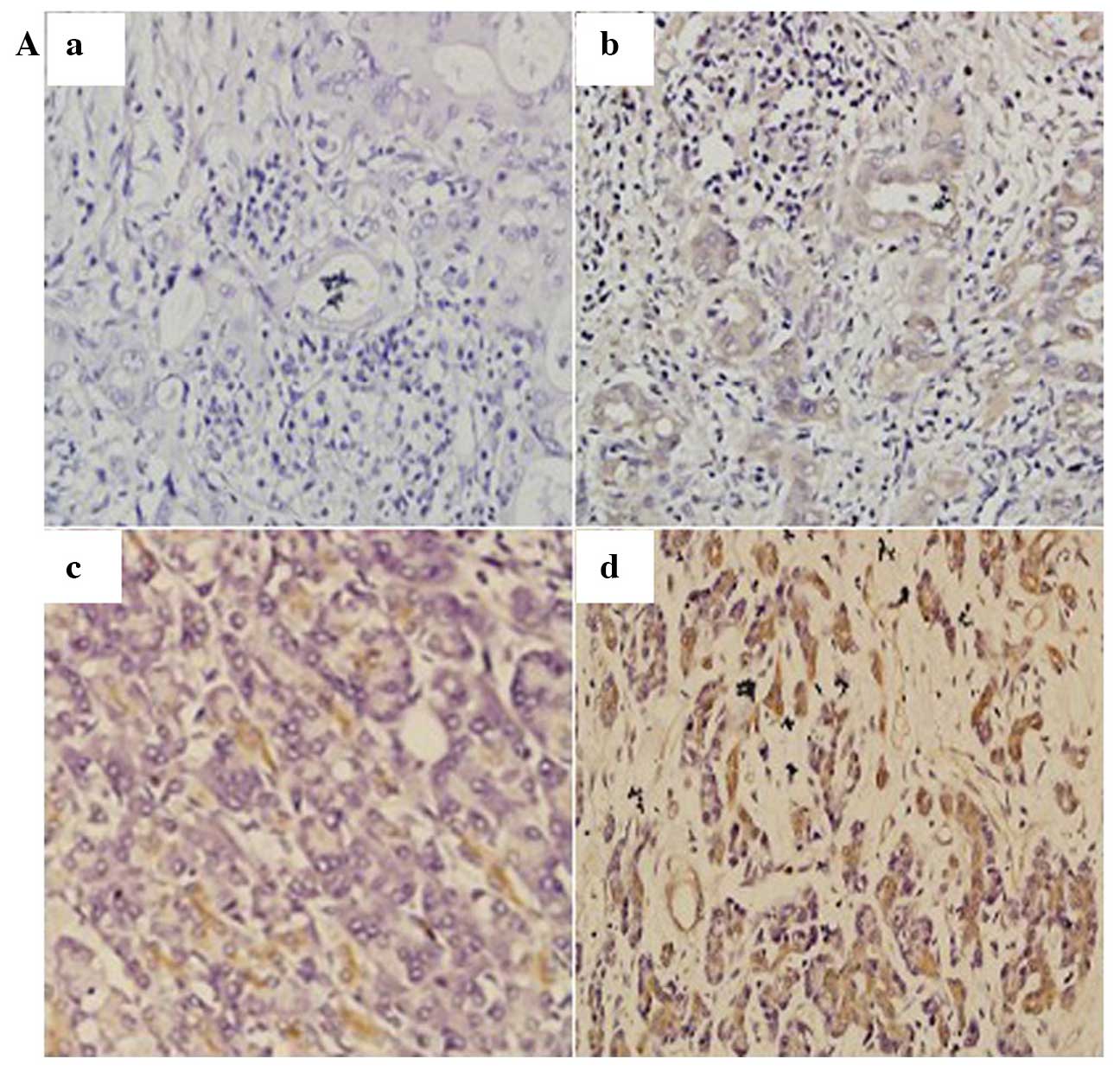
The expression of EBP50 was detected in pancreatic
cancer tissues and corresponding non-tumor tissues. IHC results
demonstrated that EBP50 was expressed in the majority of the
pancreatic cancer tissues (75/120, 62.5%); 38 tissue samples
(31.67%) were scored as 1+, 22 (18.33%) cases were scored as 2+ and
15 (12.5%) cases were scored as 3+ (Fig.
1A). In addition, the RT-qPCR data indicated that EBP50 mRNA
expression was significantly decreased in pancreatic cancer tissues
compared with the levels in corresponding normal tissues (Fig. 1B).
Overexpression of EBP50 represses
pancreatic cancer cell growth
PANC-1 and SW1990 cells were transfected with
pBK-CMV-HA-EBP50 or pBK-CMV-HA vector to establish transfected
cells. G418 solution was then used to screen the stably transfected
cells. To evaluate the transfection efficiency, western blotting
was used to determine the protein expression of EBP50 in the
pancreatic cancer cells [this data has been published previously
(11)]. It was observed that EBP50
protein was upregulated in pBK-CMV-HAEBP50-transfected group
compared with the levels in the pBK-CMV-HA vector-transfected cells
and untreated groups.
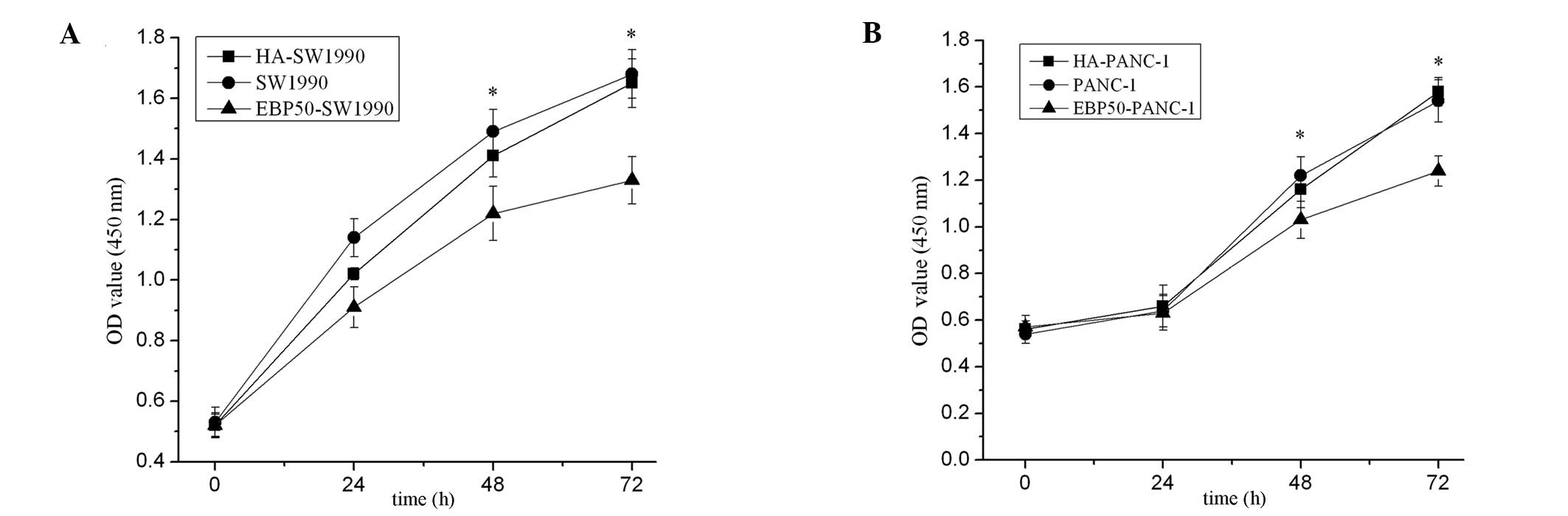
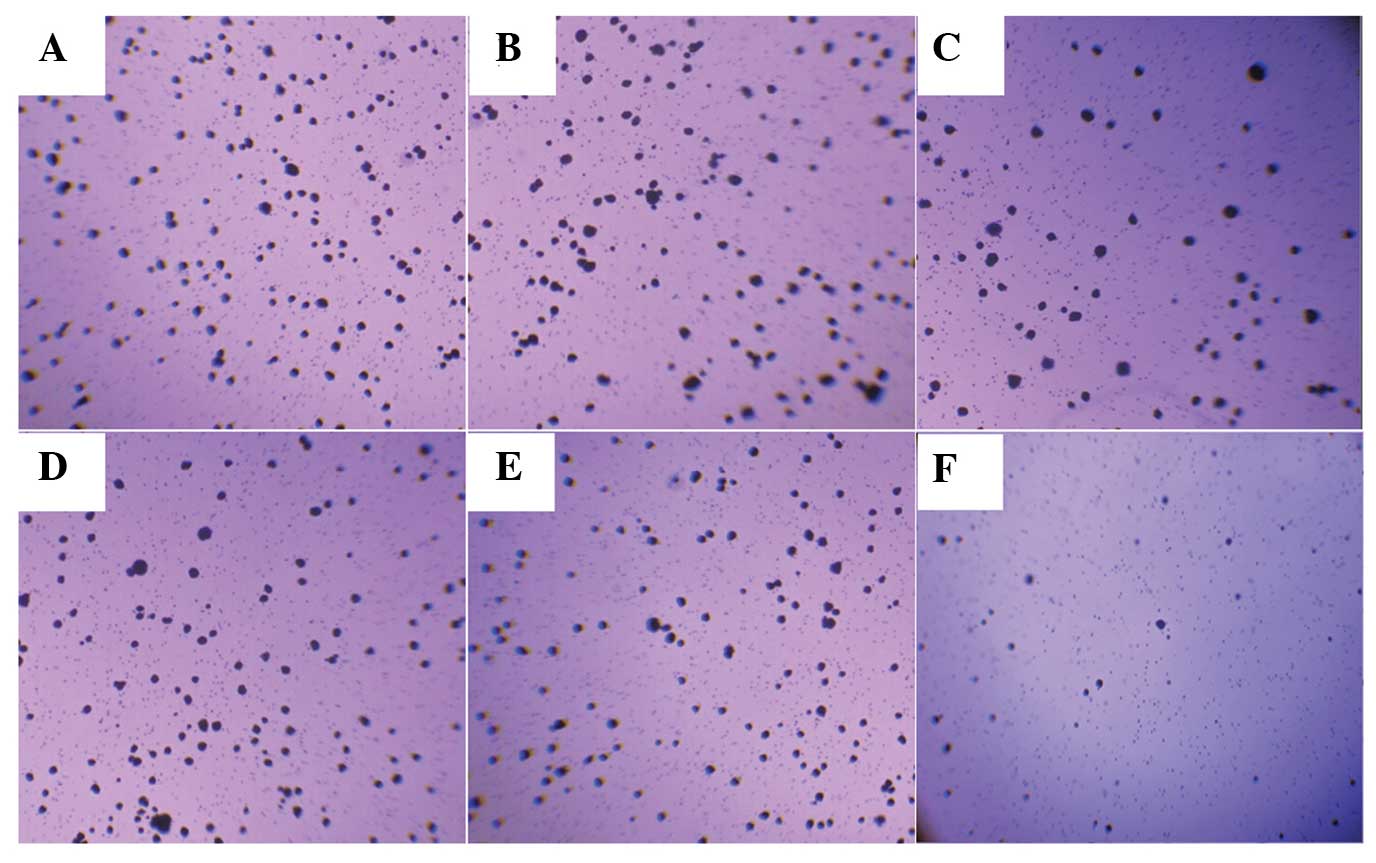
CCK-8 assays and a colony formation assay were used
to assess the potential effects of EBP50 overexpression on cell
growth and anchorage-independent growth. The number of viable cells
at different times were measured in vitro. The results
showed that the pBK-CMV-HA-EBP50-transfected cells had markedly
reduced cell growth (Fig. 2) and
anchorage-independent growth (Fig.
3) compared with the untransfected cells, whereas the empty
vector had no effect on pancreatic cancer cell growth.
Overexpression of EBP50 inhibits
cancer cell migration and invasion
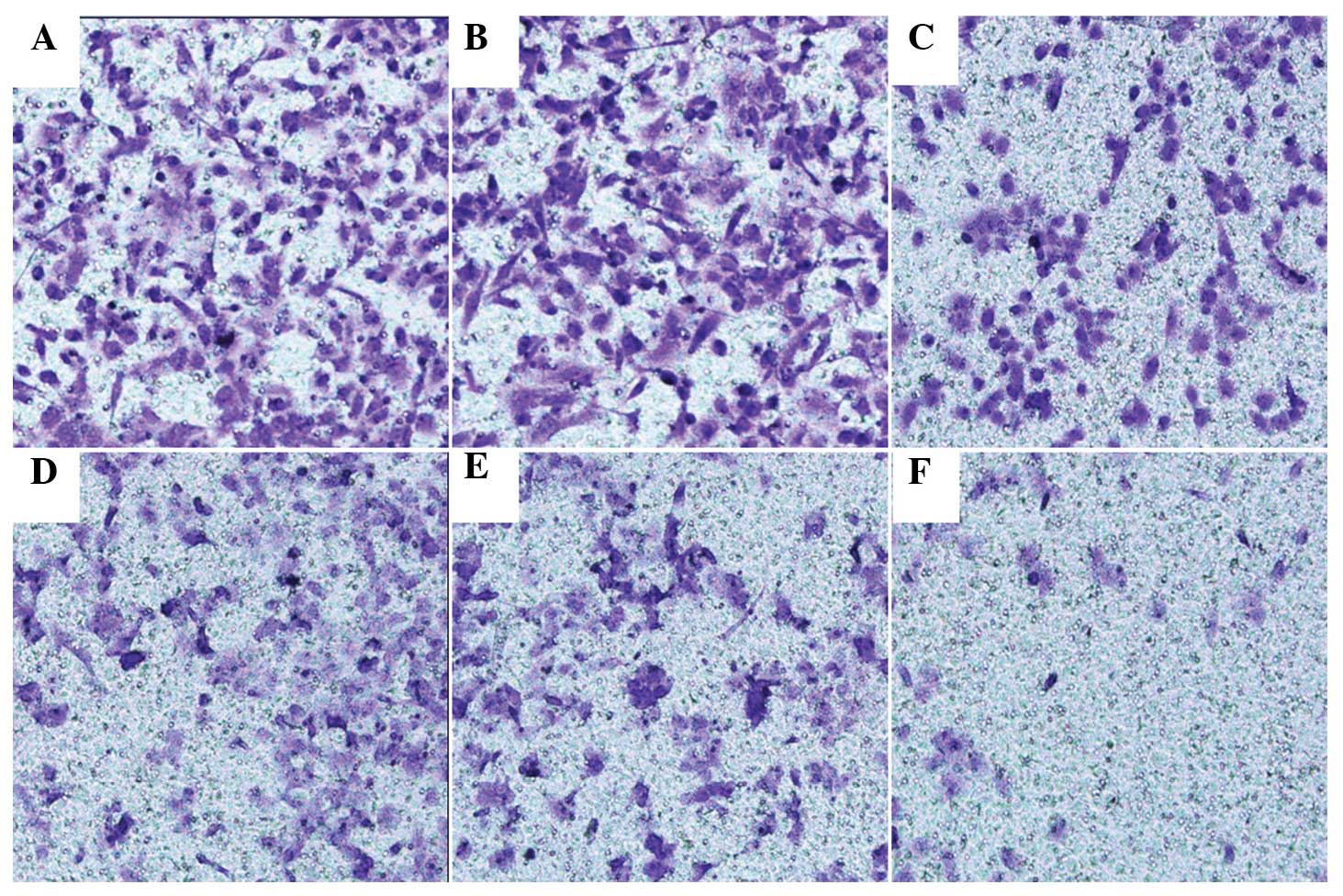
Transwell assay was used to explore whether the
overexpression of EBP50 restrained the invasion and migration of
pancreatic cancer cells. In SW1990 cells, the number of cells
transfected with pBK-CMV-HA-EBP50 that migrated was less than that
of the empty vector-transfected and untreated cells. In PANC-1
cells, the number of cells transfected with pBK-CMV-HA-EBP50 that
migrated was less than that of the empty vector-transfected and
untreated cells. No significant difference was observed between the
migration and invasion of untreated and empty vector-transfected
PANC-1 and SW1990 cells (Fig. 4).
The data indicate that the overexpression of EBP50 significantly
decreased the invasive ability of pancreatic cancer cells.
Expression of β-catenin and E-cadherin
in pancreatic cancer cells
To study the potential mechanisms underlying the
biological effect of EBP50 in pancreatic cancer cells, the
expression of β-catenin and E-cadherin was analyzed in
pBK-CMV-HA-EBP50- and pBK-CMV-HA-transfected cells and untreated
cells. The data showed that protein levels of β-catenin were
reduced and E-cadherin were increased in EBP50-overexpressing cells
compared with those in the untreated cells (Fig. 5).
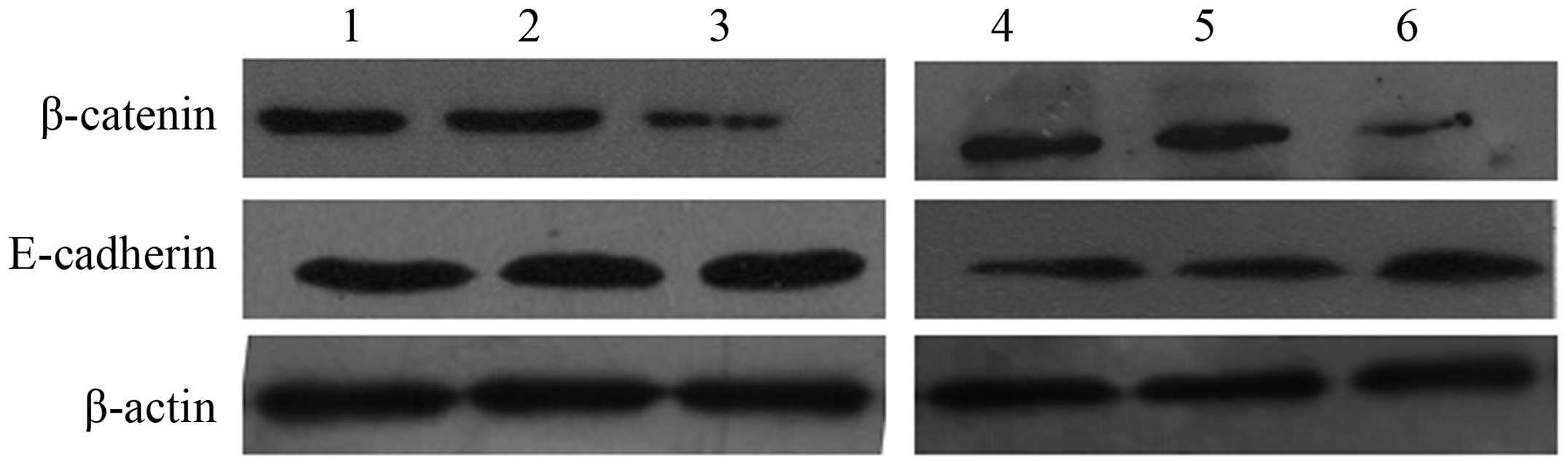 | Figure 5.β-catenin, and E-cadherin detected by
western blotting. Representative data of: Lane 1, SW1990 cells;
lane 2, HA-SW1990 cells; lane 3, EBP50-SW1990 cells; lane 4, PANC-1
cells; lane 5, HA- PANC-1 cells; and lane 6, EBP50- PANC-1 cells.
EBP50, ezrin-radixin-moesin-binding phosphoprotein 50; HA-SW1990,
SW1990 cells transfected with pBK-CMV-HA empty vector;
EBP50-SW1990, SW1990 cells transfected with pBK-CMV-HV-EBP50. |
Discussion
In the present study, it was found that the
expression of EBP50 was significantly decreased in pancreatic
cancer tissues compared with that in noncancerous tissues. To
detect the effect of EBP50 overexpression on pancreatic cancer cell
growth and invasion, the pBK-CMV-HA-EBP50 plasmid was used to
increase the expression of EBP50 in PANC-1 and SW1990 cells. The
CCK-8 and colony formation assays showed that the growth rate of
cells overexpressing EBP50 was lower than that of the two control
groups. In a Transwell assay, the number of migrated cells in the
EBP50-overexpressing group was significantly lower than that in the
control group. These results indicate that the overexpression of
EBP50 suppressed the growth and invasion of pancreatic cancer
cells. This is consistent with our previous studies (10,11,13,15).
There have been many studies in which the molecular
mechanisms by which EBP50 exerts its tumor suppressor functions are
reported (8–12). EBP50 links with the majority of
protein receptors, including the parathyroid hormone type 1,
β2-adrenergic, κ-opioid, parathyroid hormone type 1 and G
protein-coupled receptors, through the first PDZ domain (8,16–19). The
second PDZ domain binds with β-catenin, sodium-hydrogen exchanger 3
(NHE3) and Yes-associated protein 65 (Yap 65) (10). Both β-catenin and E-cadherin have an
important role in cell-cell junctions in the Wnt signaling pathway
(20). Previous studies have shown
that the downregulation of EBP50 can promote pancreatic cancer cell
growth through increasing β-catenin expression (13,21,22).
However, there have been no further studies concerning the effect
of EBP50 on the β-catenin/Wnt signaling pathway and its target
protein. These data demonstrate that the overexpression of EBP50
suppresses pancreatic cancer cell growth and invasion through
decreasing β-catenin and increasing E-cadherin levels. Lazar et
al have reported the observation of disorganization of
E-cadherin-mediated adherens junctions and an increase in β-catenin
transcriptional activity in EBP50-deficient MEFs (23). It may be hypothesized that the
balance of β-catenin/E-cadherin, which is influenced by EBP50
expression, leads to pancreatic cancer development or
progression.
These data indicate that EBP50 may be a promising
target for therapeutic intervention in pancreatic cancer. In
addition, the tumor suppressor properties of EBP50 in pancreatic
cancer may be optimized by interaction with β-catenin and
E-cadherin.
References
|
1
|
Bosetti C, Bertuccio P, Negri E, La
Vecchia C, Zeegers MP and Boffetta P: Pancreatic cancer: Overview
of descriptive epidemiology. Mol Carcinog. 51:3–13. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ashour AA, Gurbuz N, Alpay SN, AbdelAziz
AA, Mansour AM, Huo L and Ozpolat B: Elongation factor-2 kinase
regulates TG2/β1 integrin/Src/uPAR pathway and
epithelial-mesenchymal transition mediating pancreatic cancer cells
invasion. J Cell Mol Med. 18:2235–2251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dores GM, Curtis RE, van Leeuwen FE,
Stovall M, Hall P, Lynch CF, Smith SA, Weathers RE, Storm HH,
Hodgson DC, et al: Pancreatic cancer risk after treatment for
Hodgkin lymphoma. Ann Oncol. 25:2073–2079. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cui J, Jiang W, Wang S, Wang L and Xie K:
Role of Wnt/β-catenin signaling in drug resistance of pancreatic
cancer. Curr Pharm Des. 18:2464–2471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kyuno D, Yamaguchi H, Ito T, Kono T,
Kimura Y, Imamura M, Konno T, Hirata K, Sawada N and Kojima T:
Targeting tight junctions during epithelial to mesenchymal
transition in human pancreatic cancer. World J Gastroenterol.
20:10813–10824. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bao J, Wang S, Gunther LK, Kitajiri SI, Li
C and Sakamoto T: The actin-bundling protein TRIOBP-4 and -5
promotes the motility of pancreatic cancer cells. Cancer Lett.
356:367–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Z and Ma Q: Beta-catenin is a
promising key factor in the SDF-1/CXCR4 axis on metastasis of
pancreatic cancer. Med Hypotheses. 69:816–820. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Voltz JW, Brush M, Sikes S, Steplock D,
Weinman EJ and Shenolikar S: Phosphorylation of PDZ1 domain
attenuates NHERF-1 binding to cellular targets. J Biol Chem.
282:33879–33887. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fuchs SY, Ougolkov AV, Spiegelman VS and
Minamoto T: Oncogenic beta-catenin signaling networks in colorectal
cancer. Cell Cycle. 4:1522–1539. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Georgescu MM, Morales FC, Molina JR and
Hayashi Y: Roles of NHERF1/EBP50 in cancer. Curr Mol Med.
8:459–468. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji M, Yuan L, Lv X, Dong W and Peng X:
EBP50 regulates the apoptosis of pancreatic cancer cells by
decreasing the expression levels of Bcl-2. Exp Ther Med. 8:919–924.
2014.PubMed/NCBI
|
|
12
|
Lv XG, Ji MY, Dong WG, Lei XF, Liu M, Guo
XF, Wang J and Fang C: EBP50 gene transfection promotes
5-fluorouracil-induced apoptosis in gastric cancer cells through
Bax- and Bcl-2-triggered mitochondrial pathways. Mol Med Rep.
5:1220–1226. 2012.PubMed/NCBI
|
|
13
|
Ji MY, Fan DK, Lv XG, Peng XL, Lei XF and
Dong WG: The detection of EBP50 expression using quantum dot
immunohistochemistry in pancreatic cancer tissue and down-regulated
EBP50 effect on PC-2 cells. J Mol Histol. 43:517–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma C, Nong K, Wu B, Dong B, Bai Y, Zhu H,
Wang W, Huang X, Yuan Z and Ai K: miR-212 promotes pancreatic
cancer cell growth and invasion by targeting the hedgehog signaling
pathway receptor patched-1. J Exp Clin Cancer Res. 33:542014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun C, Zheng J, Cheng S, Feng D and He J:
EBP50 phosphorylation by Cdc2/Cyclin B kinase affects actin
cytoskeleton reorganization and regulates functions of human breast
cancer cell line MDA-MB-231. Mol Cells. 36:47–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cardone RA, Greco MR, Capulli M, Weinman
EJ, Busco G, Bellizzi A, Casavola V, Antelmi E, Ambruosi B,
Dell'Aquila ME, et al: NHERF1 acts as a molecular switch to program
metastatic behavior and organotropism via its PDZ domains. Mol Biol
Cell. 23:2028–2040. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Georgescu MM, Cote G, Agarwal NK and White
CL III: NHERF1/EBP50 controls morphogenesis of 3D colonic glands by
stabilizing PTEN and ezrin-radixin-moesin proteins at the apical
membrane. Neoplasia. 16:365.e1-e2–374.e1-e2. 2014. View Article : Google Scholar
|
|
18
|
Guo G, Yao G, Zhan G, Hu Y, Yue M, Cheng
L, Liu Y, Ye Q, Qing G, Zhang Y and Liu H: N-methylhemeanthidine
chloride, a novel Amaryllidaceae alkaloid, inhibits pancreatic
cancer cell proliferation via down-regulating AKT activation.
Toxicol Appl Pharmacol. 280:475–483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bellizzi A, Malfettone A, Cardone RA and
Mangia A: NHERF1/EBP50 in breast cancer: Clinical perspectives.
Breast Care (Basel). 5:86–90. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun GY, Wu JX, Wu JS, Pan YT and Jin R:
Caveolin-1, E-cadherin and β-catenin in gastric carcinoma,
precancerous tissues and chronic non-atrophic gastritis. Chin J
Cancer Res. 24:23–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kreimann EL, Morales FC, de Orbeta-Cruz J,
Takahashi Y, Adams H, Liu TJ, McCrea PD and Georgescu MM: Cortical
stabilization of beta-catenin contributes to NHERF1/EBP50 tumor
suppressor function. Oncogene. 26:5290–5299. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cornez I and Taskén K: Spatiotemporal
control of cyclic AMP immunomodulation through the PKA-Csk
inhibitory pathway is achieved by anchoring to an Ezrin-EBP50-PAG
scaffold in effector T cells. FEBS Lett. 584:2681–2688. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lazar CS, Cresson CM, Lauffenburger DA and
Gill GN: The Na+/H+ exchanger regulatory factor stabilizes
epidermal growth factor receptors at the cell surface. Mol Biol
Cell. 15:5470–5480. 2004. View Article : Google Scholar : PubMed/NCBI
|