Introduction
Cataract, the foremost cause of visual impairment,
is found notably in diabetic patients and causes health and
economic problems predominantly in developing countries (1). Genetic and environmental stresses
combined with age are considered as leading contributors to
aggregation, crystalline modification and pathogenesis in lens
oxidation. The ciliary body and blood vessels of the iris supply
glucose and oxygen (O2) to the lens, where it is present
in a hypoxic environment containing 0.5–2.3% O2
(2–4). The level of O2 is a very
important factor in cataract, and various cataractogenic stressors
such as hypoxia, hypoxic conditions along with a low glucose level
(5) or high glucose level (6), homocysteine (7), and galactose (6) are found to induce stress in the
endoplasmic reticulum (ER), thereby mediating the activation of the
unfolded protein response (UPR) along with the production of
reactive oxygen species (ROS), which is usually abnormally
increased, and lens epithelial cell (LEC) death (3,8,9).
Osmotic stress, primarily caused by the accumulation
of sugars, has been found to induce stress in the ER, which is a
major site of protein synthesis, thereby leading to free radical
generation. The stress is also found to result from fluctuations in
glucose levels initiating an UPR that further generates ROS, and
thus causes oxidative stress damage to lens fibers (10).
ER-induced UPR is reported to be activated by the
phosphorylation of inositol-requiring enzyme-1 (IRE-1), PKR-like
endoplasmic reticulum kinase (PERK) and eukaryotic translation
initiation factor 2α (eIF2α) and this mechanism is also reported to
be protective, although the cellular components are a stress
response, whereas in prolonged UPR, apoptosis is induced by
caspases (caspase-12) (11) and
C/EBP homologous protein (CHOP) (12), death factors that are activated by
activating transcription factor 4 (ATF4), which is a significant
controller of mammalian lens development (13). Free radicals such as ROS are
generated by the UPR in increased intensities (14) and in response, the upregulation of
NF-E2-related factor 2 (Nrf2) and activation of the PERK-dependent
antioxidant defense system occurs due to the UPR (15).
This study aimed to determine whether hypoxic
conditions or O2 fluctuation in the environment of LECs
induces UPR, leading to ROS production and failure of the
Nrf2-dependent antioxidant defense protection. To elucidate this,
UPR was studied along with the production of free radicals and the
levels of Nrf2 in human LECs (hLECs) that were treated with various
O2 environments.
Materials and methods
hLEC culture
hLECs (Lonza Clonetics™, Basel, Switzerland) were
cultured overnight in Dulbecco's modified Eagle's medium (DMEM;
Invitrogen Life Technologies, Carlsbad, CA, USA) containing 25 mM
glucose, along with 10% fetal calf serum (FCS) under 20%
atmospheric O2 at 37°C. Prior to each experiment, the
hLECs were precultured overnight in DMEM with 5 mM glucose and 4%
atmospheric O2. The hLECs were then cultured under
various conditions. Some were cultured in glucose-free (GF) DMEM
supplemented with 2% FCS and were maintained at 37°C in 20%
atmospheric O2. Some hLECs were maintained under
anaerobic conditions (0% atmospheric O2) in an AnaeroGen
vacuum bag (Sigma-Aldrich, St. Louis, MO, USA). Atmospheric
O2 environments of 1 and 4% were maintained in an
O2/CO2 incubator (SANYO MCO-19M
CO2 incubator; Sanyo, Tokyo, Japan) attached to a liquid
nitrogen gas tank. A normal tissue culture incubator (SANYO MCO-19M
CO2 incubator; Sanyo) was used to maintain the 20%
atmospheric O2 environment.
Cell viability/death and ROS
staining
Ethidium homodimer-1 (EthD) and calcein AM
(Viability/Cytotoxicity assay kits; Biotium Inc., Hayward, CA, USA)
mixtures were used to stain the cultured hLECs and used in
accordance with the manufacturer's recommendations.
2′,7′-Dichlorodihydrofluorescein diacetate (H2-DCFH-DA; Invitrogen
Life Technologies, Grand island, NY, USA) in phosphate-buffered
saline (PBS) at a concentration of 1 mg/ml was used to determine
the cytosolic ROS level. The mixture was allowed to stand for 40
min at 20°C, then washed with PBS twice and subjected to
microscopic imaging using a fluorescence microscope (Nikon
TE2000-U; Nikon Corporation, Tokyo, Japan).
Evaluation of protein levels in
hLECs
Protein levels were determined by western blotting.
RIPA buffer (Cell Signaling Technology, Danvers, MA, USA) was used
to lyse the cultured hLECs and the proteins were separated by
SDS-PAGE. The gels were then blotted onto nitrocellulose membranes,
which were blocked in PBS buffer (pH 8.0) containing 5% non-fat
milk for 1 h. The primary antibodies were to the following
proteins: Binding immunoglobulin protein (BiP; cat. no. sc-33757;
rabbit polyclonal IgG, 1:500; Santa Cruz Biotechnology Inc.,
Dallas, TX, USA), ATF4 (cat. no. ab23760; rabbit polyclonal IgG,
1:500; Abcam, Cambridge, MA, USA), ATF6 (cat. no. ab37149; rabbit
polyclonal IgG, 1:500; Abcam), CHOP (cat. no. MA1-250; mouse
monoclonal IgG, 1:500; Invitrogen Life Technologies, Carlsbad, CA,
USA), ER oxidoreductin 1-like (Ero1-L)α (cat. no. sc-100805; mouse
polyclonal IgG, 1:500; Santa Cruz Biotechnology, Inc.), Ero1-Lβ
(cat. no. sc-162776; goat polyclonal IgG, 1:500; Santa Cruz
Biotechnology, Inc.), hypoxia-inducible factor (HIF)-1α (cat. no.
ab16066; mouse monoclonal IgG, 1:500; Santa Cruz Biotechnology,
Inc.), kelch-like ECH-associated protein 1 (Keap1; cat. no.
sc-15246; goat polyclonal IgG, 1:500; Santa Cruz Biotechnology,
Inc.), Nrf2 (cat. no. sc-722; rabbit polyclonal IgG, 1:500; Santa
Cruz Biotechnology, Inc.), protein disulfide isomerase (PDI; cat.
no. sc-20132; rabbit polyclonal IgG, 1:500; Santa Cruz
Biotechnology, Inc.), phospho (p)-eIF2α (cat. no. sc-101670; rabbit
polyclonal IgG, 1:500; Santa Cruz Biotechnology, Inc.), p-IRE1α
(cat. no. PA1-16927; rabbit polyclonal IgG, 1:500; Invitrogen Life
Technologies), p-PERK (cat. no. sc-32577; rabbit polyclonal IgG,
1:500; Santa Cruz Biotechnology, Inc.) and GAPDH (cat. no.
sc-25778; rabbit polyclonal IgG, 1:500; Santa Cruz Biotechnology
Inc.). Anti-caspase-4 antibodies (cat. no. sc-56056; mouse
monoclonal IgG; 1:500; Santa Cruz Biotechnology Inc.) were used in
western blot analysis. The membranes were incubated with primary
antibody at 4°C overnight and then with the secondary antibodies
(cat. no. sc-56056; goat anti-rabbit IgG-horseradish peroxidase
goat polyclonal, 1:5000; Santa Cruz Biotechnology Inc.) for 1 h at
room temperature. The intensity of each band was normalized to that
of GAPDH, and the data are presented as relative intensities, which
were determined using ImageJ analysis software (National Institutes
of Health, Bethesda,. MD, USA).
Evaluation of mRNA levels in
hLECs
Total RNA was extracted from the hLECs exposed to
various O2 environments using TRIzol reagent (Invitrogen
Life Technologies) according to the manufacturer's instructions.
The purified total RNA was reverse transcribed using iScript™
Reverse Transcription Supermix for real-time PCR (Bio-Rad T100;
Bio-Rad, Hercules, CA, USA) following the manufacturer's
instructions. The reverse transcribed RNA was analyzed by
quantitative polymerase chain reaction using SsoFast™ EvaGreen®
Supermix (Bio-Rad). The primer sequences for Nrf2, Keap1, and
β-actin as described by Elanchezhian et al (5) were used for mRNA detection. The primer
sequences for Nrf2, Keap1, and β-actin were: Nrf2 sense,
5′-ACACGGTCCACAGCTCATC-3′ and antisense,
5′-TGCCTCCAAAGTATGTCAATCA-3′: Keap1 sense, 5′-GGGTCCCCTACAGCCAAG-3′
and antisense, 5′-TGGGGTTCCAGAAGATAAGC-3′; and β-actin sense
5′-CCAACCGCGAGAAGATGA-3′ and 5′-CCAGAGGCGTACAGGGATAG-3′ antisense.
Each reaction was carried out in triplicate and three independent
experiments were run. A standard curve was prepared, relative copy
numbers were obtained from the standard curve and the relative
expression levels were normalized to the values obtained for
β-actin.
Statistical analysis
Results are presented as the mean ± standard
deviation from three individual experiments. P-values were
determined by Student's t-tests and analyzed using SPSS software,
(version 16.0; SPSS Inc., Chicago, IL, USA).P<0.05 was
considered to indicate a statistically significant result.
Results
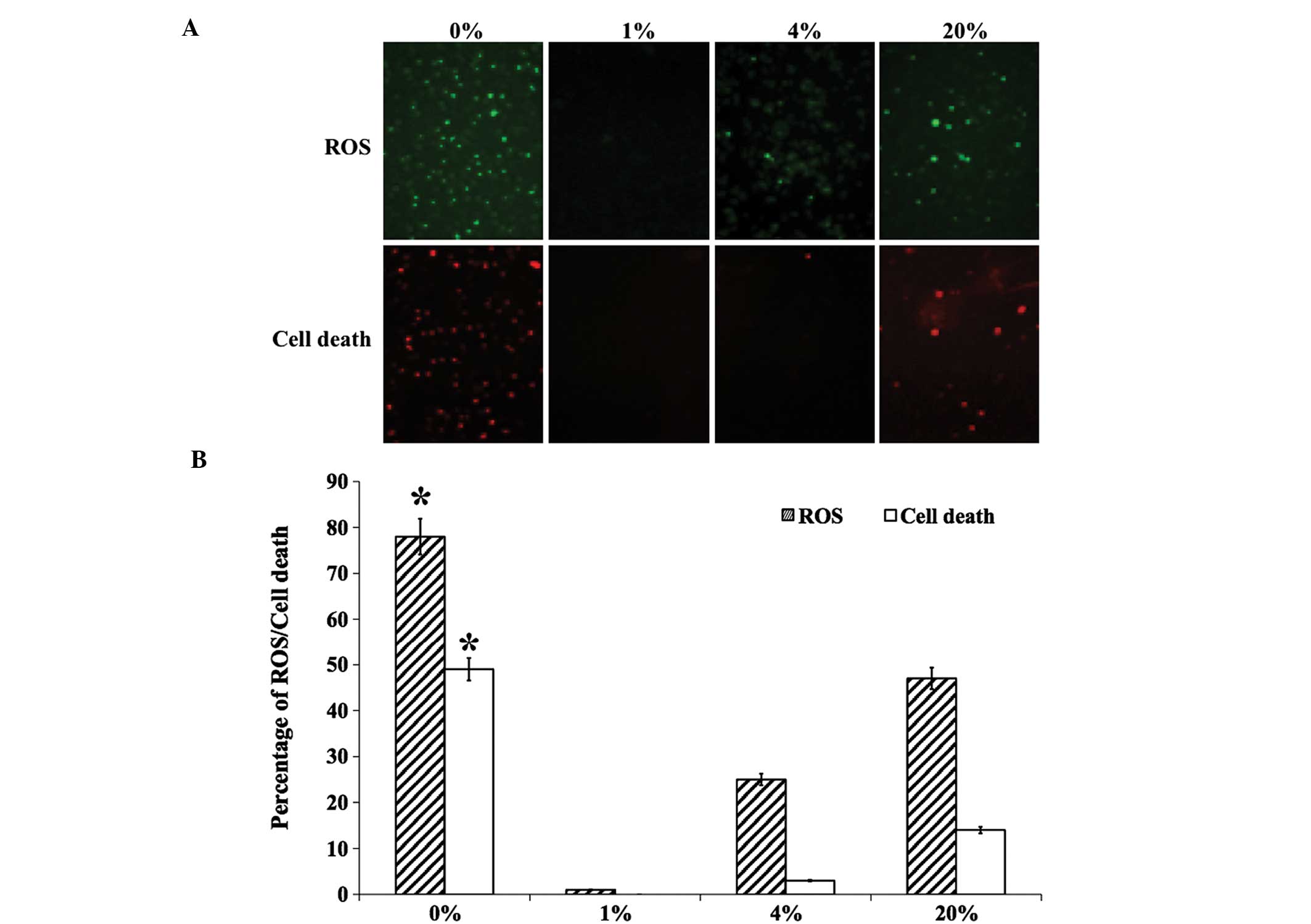
ROS production and cell death
hLECs that were cultured in different levels of
O2 were stained for the evaluation of ROS production and
cell death (Fig. 1A). The production
of ROS was found to be significantly increased in 0 and 20%
O2 than in 1 and 4% O2 (Fig. 1B). A similar pattern was observed for
dead cell staining, with no cell death observed in 1 and 4%
O2 (Fig. 1). The levels
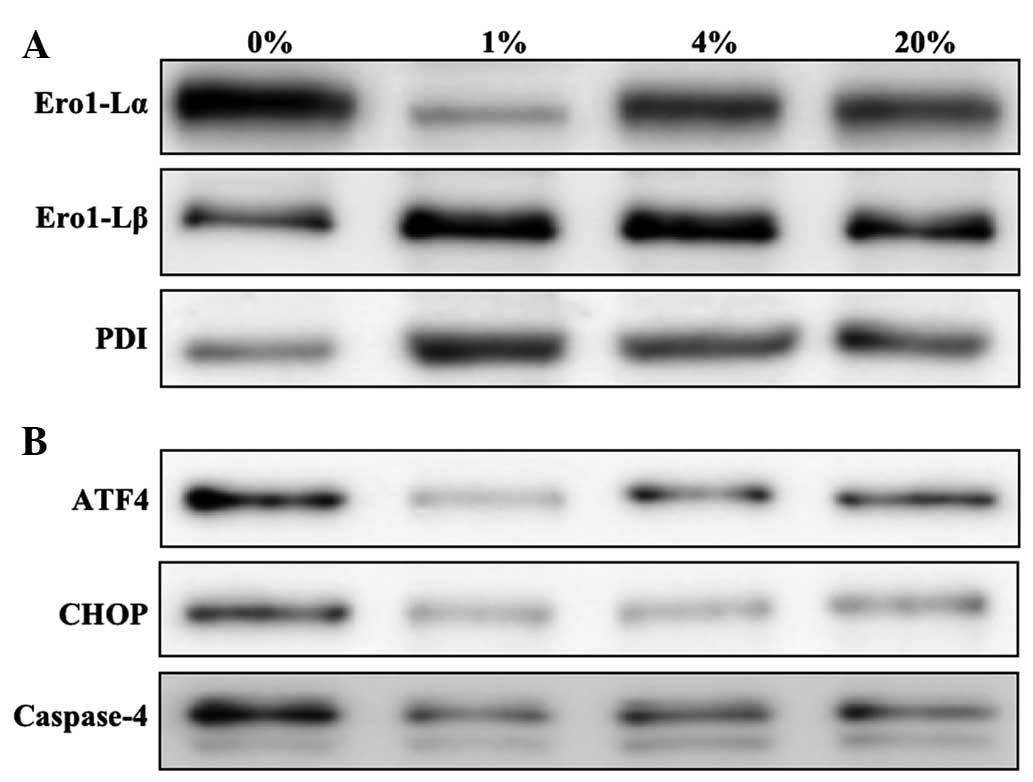
of ROS- (Fig. 2A) and
apoptosis-related ER stress proteins (Fig. 2B) were then investigated. The
ROS-related ER stress proteins that were investigated were Ero1-Lα,
Ero1-Lβ and PDI. An increased level of Ero1-Lα was detected in
cells cultured with 0, 4 and 20% O2. However, Ero1-Lβ
and PDI were detected at greater levels in hLECs cultured in 1%
O2 than in those cultured in other percentages of
O2 (Fig. 2A). The
apoptosis-related ER stress proteins that were investigated were
ATF4, CHOP and caspase-4. Similar to the ROS-related protein
Ero1-Lα, apoptotic proteins were detected in greater quantities in
hLECs cultured in 0% O2 than in those cultured in other
percentages of O2 (Fig.
2B). Notably, cells cultured in 1% O2 revealed a
protective effect when compared with the cells cultured in other
percentages of O2.
Detection of O2 deprivation
with different percentages of O2
hLECs were cultured in 0, 1, 4 and 20% O2
and cells were collected at various time intervals (1, 6, 12 and 24
h) for the investigation of HIF-1α levels. Increased levels of
HIF-1α were detected in the cells cultured in 0% O2 from
6 h, and the HIF-1α levels were significantly increased following
24 h of culture (Fig. 3). However,
the cells cultured in 1% O2 showed minimum levels of
HIF-1α when compared with those cultured in 0% O2. The
other O2 percentages, 4 and 20%, showed a minimal HIF-1α
protein levels (Fig. 3).
Evaluation of ER stress proteins
To elucidate the hypothesis concerning the induction
of ER stress by O2 fluctuation, the levels of ER stress
proteins in cells cultured for 24 h with 0, 1, 4 and 20%
O2 were investigated. The ER stress marker protein BiP
and the UPR-associated proteins p-PERK, p-IRE1α, p-eIF2α and ATF6
were investigated. Increased levels of ER stress proteins were
found in cells cultured with 0 and 20% O2 when compared
with those cultured with 1 and 4% O2 (Fig. 4). However, very minimal or negligible
amounts of ER stress proteins were detected in cells cultured in 1%
O2, which clearly indicates that this is a protective
environment for the growth of hLECs.
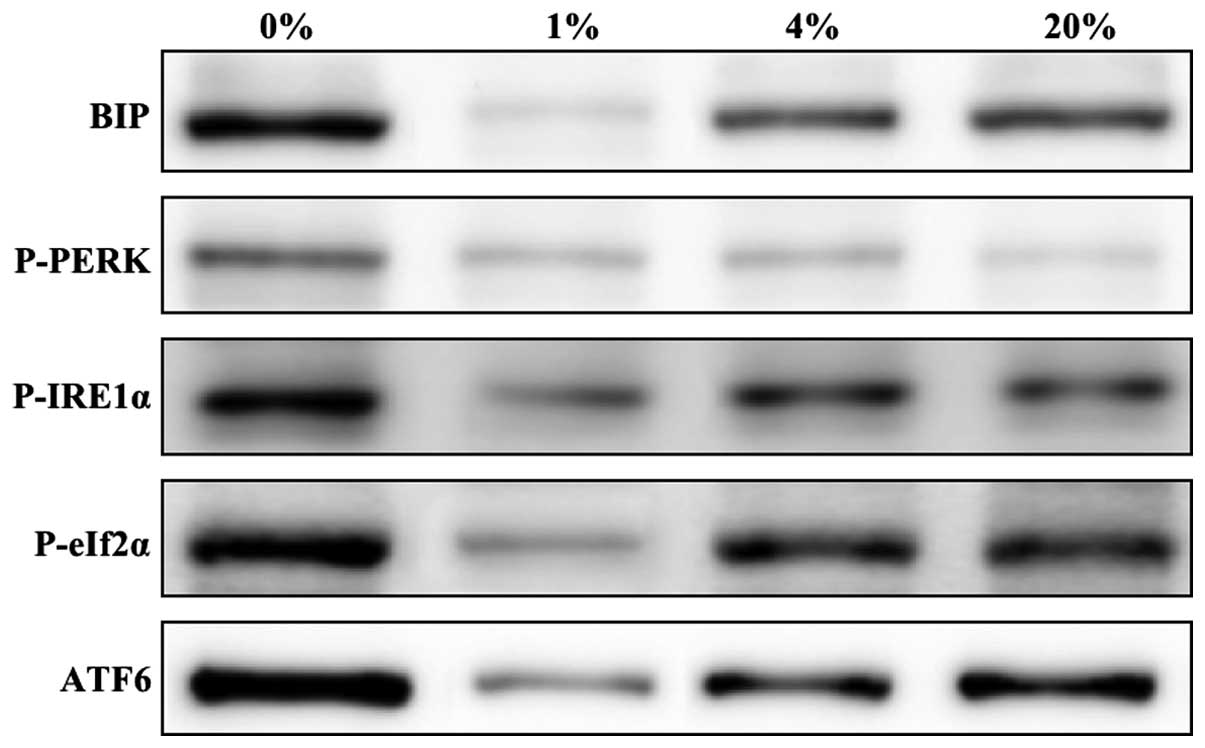 | Figure 4.Representative western blots of ER
stress-related UPR proteins in hLECs cultured for 24 h with 0, 1, 4
and 20% oxygen. ER, endoplasmic reticulum; UPR, unfolded protein
response; BiP, binding immunoglobulin protein; p, phospho; PERK,
PKR-like endoplasmic reticulum kinase; IRE1α, inositol-requiring
enzyme 1α; eIf2α, eukaryotic translation initiation factor 2α;
ATF6, activating transcription factor 6. |
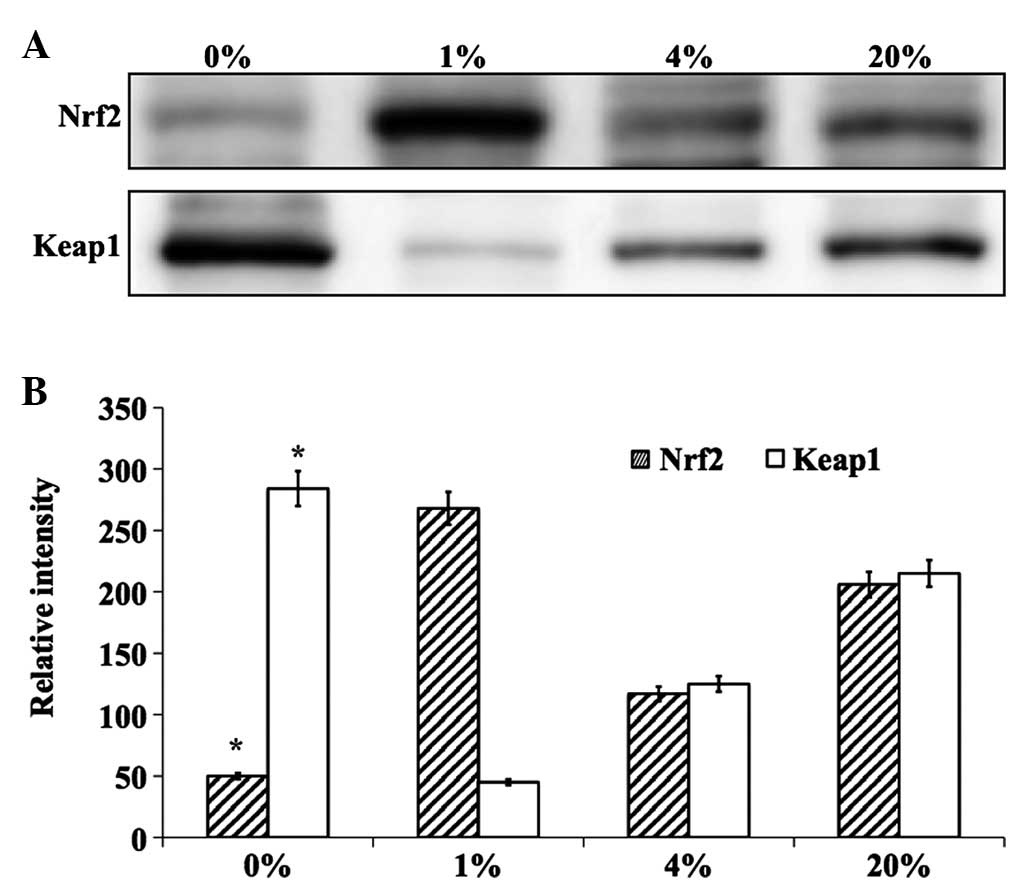
Evaluation of key antioxidant
proteins
The effect of different percentages of O2
on the levels of the lens antioxidant proteins Nrf2 and Keap1 were
investigated. The effects were evaluated in cells cultured for 24 h
in 0, 1, 4 and 20% O2 by quantifying Nrf2 and Keap1 at
the protein and mRNA levels. Notably, cells cultured in 1%
O2 showed a protective effect and the level of Nrf2
protein was increased to near normal levels (Fig. 5). However, cells cultured in 0, 4 and
20% O2 showed decreased levels of this protein. The
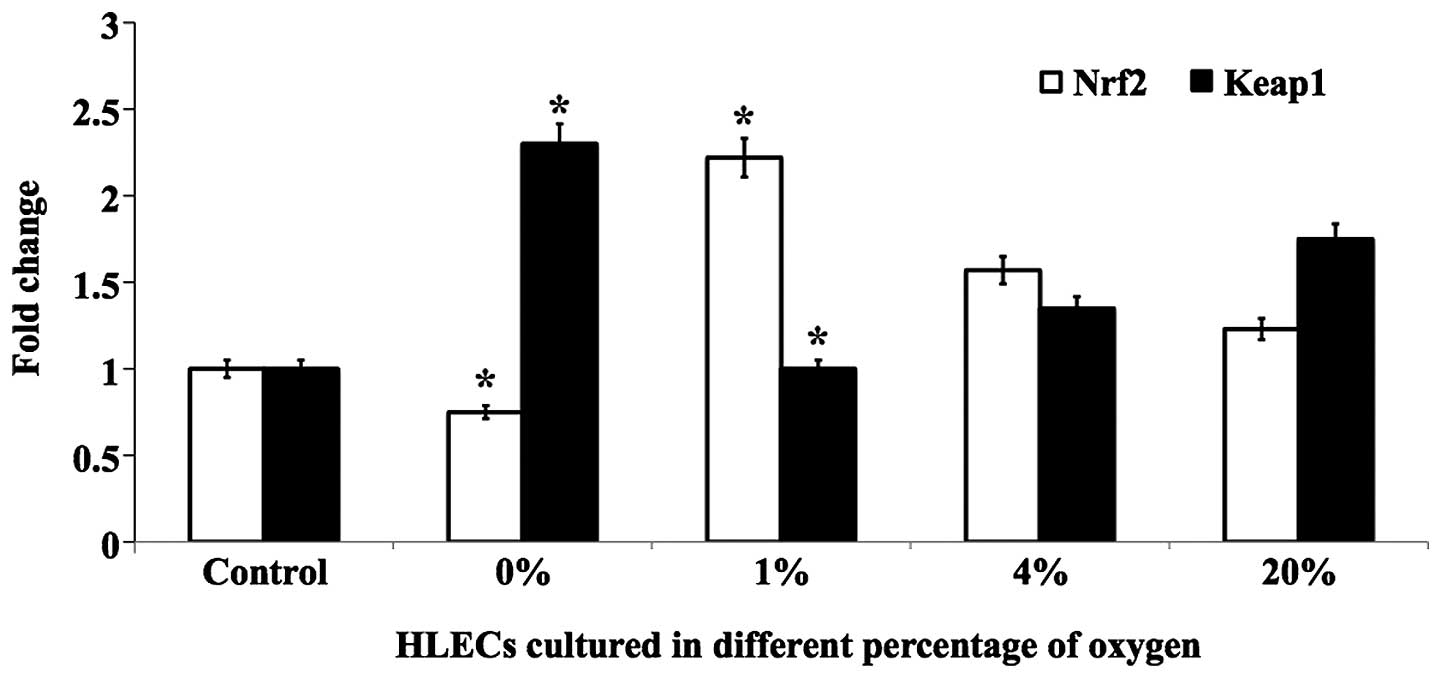
inverse effect was observed for Keap1 protein (Fig. 5). The mRNA levels of Nrf2 and Keap1
followed similar trends (Fig. 6).
Altogether, it is clear that 1% O2 was found to be
protective for cultured hLECs when compared with other percentages
of O2. In addition, a complete lack of O2
(0%) resulted in severe damage to lens proteins due to the
induction of ER stress, where chronic exposure led to the
production of ROS and cell death.
Discussion
Lens researchers generally consider that age-related
cataracts (ARCs) are strongly associated with lens oxidation and
aging (16–18). Although a direct association between
cataract formation and hypoxia has not been clearly demonstrated,
there are many adverse conditions that lead to hypoxic disorders in
the human lens. In the present study, exposure of hLECs to certain
O2 environments was found to activate a protective UPR,
whereas prolonged exposure to severe hypoxia induced ROS production
and apoptotic UPR. This response was observed as LEC death and lens
oxidation. The obtained results are in line with previous studies,
in which cataracts under diabetic conditions have been found to be
strongly associated with age-dependent circulatory disorders and
also dependent on diastolic blood pressure, the duration of
diabetes, elevated glycosylated hemoglobin levels and lower
intraocular pressure (19,20).
In addition, metabolic diseases and extreme fasting
conditions have been found to induce hypoglycemia; a recent study
showed that a 30-min exposure to very low glucose was adequate to
induce an UPR in the germinative zone of the lens in rodents
(5) and hyperoxia and hyperglycemia
are been widely reported to induce oxidation in the lens and
cataract formation (21,22). The present study has revealed that
physiological concentrations of O2 concentrations also
induce lens oxidation and activate the UPR, indicating that hypoxia
and hyperoxia are potentially contributing factors to oxidation in
lenses.
Kiviluoto et al (23) reported that the activation of ER
stress proteins deplete the level of Ca2+ in the ER-;
the ER-Ca2+ level along with ER localized oxidative
machinery is vital for appropriate protein folding. The results of
the present study suggest that the culture of hLECs under severe
hypoxia (0% O2) or hyperoxia (20% O2) induces
apoptotic UPR leading to ROS production; this may also lead to the
release of Ca2+ from the ER, consequently increasing
cytosolic Ca2+ and thus inducing apoptosis and causing
severe impairment in the hLECs. It may be hypothesized that aged
individuals develop cortical or nuclear cataracts due to the
induction of UPR and also due to ROS production in LECs.
A study by Elanchezhian et al (5) demonstrated that ROS increased in LECs
in the germinative zone that differentiate into cortical fiber
cells, where new fiber cells are generated over old lens fiber
cells. These lens fiber cells were suggested to contain less
Nrf2-dependent antioxidant protection, and the changes result in
crystallin aggregation and oxidation in the posterior and cortical
regions. Moreover, earlier findings have reported that diabetic
cataracts are intensely associated with diabetic exposure time
(19,20), where longer exposure results in an
increased density of the cortical lens fiber cell layer.
Thus, the present study has revealed that 1%
O2 is a protective environment for the healthy growth of
hLECs. However, 0 and 20% O2 may activate the UPR, and
prolonged exposure leads to the production of ROS, oxidation of the
lens and ultimately leads to cell death. Maintaining cells in 1%
O2 can attenuate O2-fluctuation induced ER
stress. The results of this study suggest that 1% O2
provides a protective environment for the healthy cell culture and
experimental use of hLECs.
Acknowledgements
The present study was supported by grants from the
National Science Foundation of China (no. 81202021), the National
Science Foundation of Zhejiang Province, China (no. LQ13H120001)
and the Key Laboratory of Diagnosis and Treatment of Neonatal
Diseases of Zhejiang Province.
References
|
1
|
Tabin G, Chen M and Espandar L: Cataract
surgery for the developing world. Curr Opin Ophthalmol. 19:55–59.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barbazetto IA, Liang J, Chang S, Zheng L,
Spector A and Dillon JP: Oxygen tension in the rabbit lens and
vitreous before and after vitrectomy. Exp Eye Res. 78:917–924.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McNulty R, Wang H, Mathias RT, Ortwerth
BJ, Truscott RJ and Bassnett S: Regulation of tissue oxygen levels
in the mammalian lens. J Physiol. 559:883–898. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shui YB, Fu JJ, Garcia C, Dattilo LK,
Rajagopal R, McMillan S, Mak G, Holekamp NM, Lewis A and Beebe DC:
Oxygen distribution in the rabbit eye and oxygen consumption by the
lens. Invest Ophthalmol Vis Sci. 47:1571–1580. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Elanchezhian R, Palsamy P, Madson CJ,
Mulhern ML, Lynch DW, Troia AM, Usukura J and Shinohara T: Low
glucose under hypoxic conditions induces unfolded protein response
and produces reactive oxygen species in lens epithelial cells. Cell
Death Dis. 3:e3012012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ikesugi K, Yamamoto R, Mulhern ML and
Shinohara T: Role of the unfolded protein response (UPR) in
cataract formation. Exp Eye Res. 83:508–516. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Elanchezhian R, Palsamy P, Madson CJ,
Lynch DW and Shinohara T: Age-related cataracts: Homocysteine
coupled endoplasmic reticulum stress and suppression of
Nrf2-dependent antioxidant protection. Chem Biol Interact.
200:1–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Helbig H, Hinz JP, Kellner U and Foerster
MH: Oxygen in the anterior chamber of the human eye. Ger J
Ophthalmol. 2:161–164. 1993.PubMed/NCBI
|
|
9
|
Bassnett S and McNulty R: The effect of
elevated intraocular oxygen on organelle degradation in the
embryonic chicken lens. J Exp Biol. 206:4353–4361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mulhern ML, Madson CJ, Danford A, Ikesugi
K, Kador PF and Shinohara T: The unfolded protein response in lens
epithelial cells from galactosemic rat lenses. Invest Ophthalmol
Vis Sci. 47:3951–3959. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xie Q, Khaoustov VI, Chung CC, Sohn J,
Krishnan B, Lewis DE and Yoffe B: Effect of tauroursodeoxycholic
acid on endoplasmic reticulum stress-induced caspase-12 activation.
Hepatology. 36:592–601. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tinhofer I, Anether G, Senfter M, Pfaller
K, Bernhard D, Hara M and Greil R: Stressful death of T-ALL tumor
cells after treatment with the anti-tumor agent Tetrocarcin-A.
FASEB J. 16:1295–1297. 2002.PubMed/NCBI
|
|
13
|
Hettmann T, Barton K and Leiden JM:
Microphthalmia due to p53-mediated apoptosis of anterior lens
epithelial cells in mice lacking the CREB-2 transcription factor.
Dev Biol. 222:110–123. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tu BP and Weissman JS: The FAD- and
O2-dependent reaction cycle of Ero1-mediated oxidative
protein folding in the endoplasmic reticulum. Mol Cell. 10:983–994.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pagani M, Fabbri M, Benedetti C, Fassio A,
Pilati S, Bulleid NJ, Cabibbo A and Sitia R: Endoplasmic reticulum
oxidoreductin 1-Lbeta (ERO1-Lbeta), a human gene induced in the
course of the unfolded protein response. J Biol Chem.
275:23685–23692. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baynes JW and Thorpe SR: Role of oxidative
stress in diabetic complications: A new perspective on an old
paradigm. Diabetes. 48:1–9. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brennan LA and Kantorow M: Mitochondrial
function and redox control in the aging eye: Role of MsrA and other
repair systems in cataract and macular degenerations. Exp Eye Res.
88:195–203. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lou MF: Redox regulation in the lens. Prog
Retin Eye Res. 22:657–682. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harding JJ: Recent studies of risk factors
and protective factors for cataract. Curr Opin Ophthalmol. 8:46–49.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Negahban K and Chern K: Cataracts
associated with systemic disorders and syndromes. Curr Opin
Ophthalmol. 13:419–422. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Simpanya MF, Ansari RR, Suh KI, Leverenz
VR and Giblin FJ: Aggregation of lens crystallins in an in
vivo hyperbaric oxygen guinea pig model of nuclear cataract:
Dynamic light-scattering and HPLC analysis. Invest Ophthalmol Vis
Sci. 46:4641–4651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Simpanya MF, Ansari RR, Leverenz V and
Giblin FJ: Measurement of lens protein aggregation in vivo
using dynamic light scattering in a guinea pig/UVA model for
nuclear cataract. Photochem Photobiol. 84:1589–1595. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kiviluoto S, Vervliet T, Ivanova H,
Decuypere JP, De Smedt H, Missiaen L, Bultynck G and Parys JB:
Regulation of inositol 1,4,5-trisphosphate receptors during
endoplasmic reticulum stress. Biochim Biophys Acta. 1833:1612–1624.
2013. View Article : Google Scholar : PubMed/NCBI
|