Introduction
A sudden increase in the morbidity of upper
respiratory tract infections (1) and
the attack and exacerbation of autoimmune diseases (2) have been observed in the few days
following sudden environmental temperature decreases. Cold stress
has been reported to suppress host innate immune defenses.
Macrophages from mice subjected to 10 min of cold stress (at −15°C)
exhibited a lower phagocytic capacity than cells from control
animals, and corticosterone affected phagocytosis by macrophages
(3). TLR4 (Toll-like receptor 4) is
a pattern recognition receptor of pathogen-associated molecular
patterns. It has been shown that TLR4 mutant mice are more
susceptible to pulmonary tuberculosis and enhanced mycobacterial
outgrowth (4). Levels of cluster of
differentiation (CD)3+ (CD4 and CD8) T cells have been
found to be reduced following acute cold restraint stress, and the
T-cell lymphopenia was mediated predominantly through a
β2-adrenergic receptor mechanism (5). Cold exposure (−5°C) has been shown to
increase the delayed-type hypersensitivity (DTH) reaction by 42% in
calves after one week (6), and serum
interleukin-2 (IL-2) levels have been found to be reduced in rats
immersed in cold water for 30 min (7). Cold stress also affects the host
adaptive immune system.
The serum adrenocorticotropin (ACTH), epinephrine
(EPI) and angiotensin II (ANG-II) levels have been demonstrated to
be significantly higher in mice and rats exposed to the cold
compared with those in normal-temperature control group animals
(8–11). Cold stress induces an increase in
ANG-II and supports a sympathetic-mediated stress response through
stimulation of angiotensin receptor 1 [AT(1)] in humans (12).
In order to confirm whether the sudden environmental
temperature drop decreases immunity in mammals, the aim of the
present study was to observe the effect of sudden cold stress on
the ACTH, EPI, ANG- II, interferon-γ (IFN-γ), IL-2, IL-4 and IL-10
levels in the plasma, the TLR4 expression of immunocompetent cells
in the spleen and the regulatory T (Treg) cell expression in the
peripheral blood in rats. Investigations of this type may provide
insight that would aid with disease prevention following sudden
environmental temperature decreases.
Materials and methods
Study groups
A total of 252 Sprague Dawley (SD) rats were
purchased from Shanghai Xi Pu Er-Bi Kai Experimental Animal Co.,
Ltd. (Shanghai, China) (certification no. 00800161082; license no.
SCXK (Shanghai) 2008-0016). The rats had a mean weight of
275.24±35.73 g (range, 198–378 g), were aged 9–15 weeks and were
housed under a 12-h light/dark cycle with free access to food and
water, were included in the present study. Following adaptation for
seven days at 20°C, the rats were divided into 20, 4 and −12°C
groups. In each temperature group, there were four observation
time-points (1, 12, 24 and 48 h), and each time group was further
subdivided into non-stimulation, lipopolysaccharide
(LPS)-stimulation and concanavalin A (Con-A)-stimulation groups.
The body's response to bacterial invasion was investigated by
LPS-stimulation, and the response to viral invasion was
investigated by Con-A-stimulation. The rats in the 20°C group were
kept in the natural environment while the rats in the 4 and −12°C
groups were maintained in low-temperature climate incubators (model
RXZ-0450; Ningbo Southeast Instrument Co., Ltd., Ningbo, China).
The rats were given ad libitum access to food in all of the
groups; however, with regards to water, the rats in the 12, 24 and
48 h groups were given constant ad libitum access, whereas
the −12°C group were given free access at three specific time
points per day. One rat was placed in each cage. The environmental
humidity was 50%. The experimental studies with the rats were
performed in accordance with the Animal Experiment Guidelines
established by The Ministry of Science and Technology of the
People's Republic of China. The animal experiments in this paper
were approved by Jiangxi Province People's Hospital Ethics
Committee (Nanchang, China).
LPS and Con-A stimulation
Following adaptation for seven days at 20°C, LPS
(1.0 mg/kg body weight; from Escherichia coli 055:B5; cat.
no. L2880; Sigma-Aldrich, St. Louis, MO, USA) or Con-A [5.0 mg/kg
body weight; from Canavalia ensiformis (Jack bean) Type IV;
cat. no. C2010-1G; Sigma-Aldrich] were administered to the SD rats
once via intraperitoneal injection. The rats were then placed in
the natural environment (20°C) or the 4 and −12°C environment
cabinets for 1, 12, 24 and 48 h.
Spleen mononuclear cell (MNC)
suspension preparation
The rats were anesthetized by pentobarbital sodium
via intraperitoneal injection, prior to the removal of the spleen.
The cell suspension was prepared by grinding the spleen with 1.0%
fetal calf serum (FCS) (Zhejiang Tianhang Biotechnology Co., Ltd.,
Hangzhou, China), 0.03% NaN3 (Binhai Hanhong Biochemical
Co., Ltd., Shanghai, China) and 0.01 M phosphate-buffered saline
(PBS) (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) (pH 7.4) and filtering through 200-mesh stainless
steel nets. The MNCs were separated by Ficoll-Hypaque (Tianjin Hao
Yang Biological Manufacture Co., Ltd., Tianjin, China) density
gradient centrifugation and washed two times in 1.0% FCS, 0.03%
NaN3 and 0.01 M PBS via centrifugation at 400 × g for 10
min at 4°C (Allegra™ 6R, Beckman Coulter, Miami, FL, USA) for 10
min. The cell concentration was then adjusted to
1×107/ml.
Fluorescence staining and
detection
TLR4 expression of MNCs
A total of 10 µl mouse monoclonal
anti-TLR4-phycoerythrin (PE) antibody (1.0 µg/10 µl; cat. no.
ab45104 Abcam, Hong Kong SAR, China) was added to the bottom of a
plastic tube; as a control, 5 µl isotype control antibody [mouse
monoclonal isotype control PE immunoglobulin G (IgG)2b κ; cat. no.
12–4732 eBioscience, Inc., San Diego, CA, USA] was added to another
tube. A total of 5×105 MNCs/50 µl was then added to each
tube, and the tubes were vortexed and placed at room temperature,
avoiding the light, for 40 min. PBS (2 ml) was added to each tube,
and the mixture was washed with PBS two times via centrifugation
(400 × g, 5 min, 4°C). Following centrifugation, 0.4 ml flow
cytometric sheath fluid was added to each tube, and
TLR4+ cells were detected using a flow cytometer
(Coulter Epics XL; Beckman Coulter). The voltage was adjusted with
the naked cells, and >30,000 cells were counted in each
tube.
Treg cell detection in the peripheral blood
A total of 5 µl mouse monoclonal anti-rat CD25-PE
(0.125 µg/5 µl; cat. no. 12-0390; eBioscience, Inc.) and 5.0 µl
mouse monoclonal PE-cyanine 5 (Cy5)-conjugated anti-rat CD4 (0.25
µg/5 µl; cat. no. 554839; BD Pharmingen, San Diego, CA, USA)
antibodies were added to the bottom of a plastic tube, and 5.0 µl
mouse monoclonal isotype control IgG1κ-PE (cat. no. 12–4714;
eBioscience, Inc.) and 5.0 µl mouse monoclonal isotype control
IgG2a κ-PE-Cy5 (cat. no. 15–4724; eBioscience, Inc.) antibodies
were added to another tube. In addition, 50 µl EDTA-anticoagulated
rat peripheral blood and 50 µl PBS were added to each tube; the
tubes were then vortexed and placed at room temperature, avoiding
the light, for 60 min. After 60 min, 0.2 ml red blood cell lysing
solution (OptiLyse® C, Beckman Coulter) was added to each tube for
incubation at room temperature for 13 min. Following incubation,
the tubes were washed two times with 2 ml PBS via centrifugation
(400 × g, 10 min, 5°C). The supernatant was then removed, and 1.0
ml 1X fixation/permeabilization solution [Forkhead box P3 (Foxp3)
Fixation/Permeabilization Concentrate and Diluent; eBioscience,
Inc.] was added to each tube, which was vortexed and incubated at
4°C, avoiding the light, for 60 min. Following incubation, 1X
permeabilization buffer solution (10X Permeabilization Buffer;
eBioscience, Inc.) was added to each tube, and the tubes were
vortexed and washed two times via centrifugation (400 × g, 10 min,
5°C). The supernatant was subsequently removed, and 5.0 µl
monoclonal fluorescein isothiocyanate (FITC)-anti-rat Foxp3
antibody (cat. no. 11-5773; eBioscience, Inc.) was added to the
CD4+CD25+ tube, while 5.0 µl monoclonal
FITC-rat IgG2a isotype control antibody (cat. no. 11–4321;
eBioscience, Inc.) was added to the isotype control tube. The tubes
were then vortexed and incubated at 4°C, avoiding the light, for 60
min. After 60 min, 1.0 ml 1X permeabilization buffer was added, and
the tubes were vortexed and washed two times via centrifugation
(400 × g, 10 min, 5°C). Following centrifugation, 0.4 ml flow
cytometric sheath fluid was added to each tube and the tubes were
vortexed, prior to the tube contents being analyzed using a flow
cytometer (Coulter Epics XL; Beckman Coulter). The voltage was
adjusted with the naked cells, and >30,000 cells were counted in
each tube. The CD4+CD25+ cells were initially
measured and counted, and then the Foxp3+ cells among
the CD4+CD25+ cells were measured and
counted.
Hormone and cytokine assay
The ACTH, EPI, ANG-II, IFN-γ, IL-2, IL-4 and IL-10
levels in the plasma were determined via sandwich ELISA using ELISA
kits from Shanghai Westang Bio-Tech Co., Ltd. (Shanghai, China) and
a Multiskan Ascent® instrument (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The procedure was performed according to the
ELISA kit specifications.
Statistical analysis
The experimental data are expressed as the mean ±
standard deviation. Data analysis was performed using SPSS 16.0
statistical software (SPSS, Inc., Chicago, IL, USA) with multi-way
univariate analysis of variance and Levene's test for equality of
error variances, and then post hoc tests were used for comparison
between two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of different temperatures and
stimulants on levels of stress hormones in the rat plasma
The mean levels of ACTH, EPI and ANG-II were
compared at different times, temperatures, and in response to
various stimulants (Figs. 1 and
2). Compared with the results at
20°C, the level of ACTH in the rat plasma was increased at the
1–48-h time-points in the 4 and −12°C non-stimulation groups; at
the 1–48-h time-points at 4°C and at the 1–24-h time-points at
−12°C in the LPS-stimulation group; and at the 1–48-h time-points
in the −12°C Con-A-stimulation group (Figs. 1A and B and 2A). With regard to EPI levels, comparisons
with the values at 20°C revealed increases in the 4°C and −12°C
non-stimulation groups at the 1–48-h time-points, in the 4°C
LPS-stimulation group at the 1–48-h time-points, in the −12°C
LPS-stimulation group at the 1–24-h time-points, in the 4°C
Con-A-stimulation group at the 1–12-h time-points and in the −12°C
Con-A-stimulation group at 1, 12 and 48 h (Figs. 1C and D and 2B). Compared with the results at 20°C, the
level of ANG-II in the rat plasma was increased in the 4°C and
−12°C non-stimulation and LPS-stimulation groups at the 1–48-h
time-points, in the 4°C Con-A-stimulation group at 1 h and in the
−12°C Con-A-stimulation group at the 1–48-h time-points (Figs. 1E and F and 2C).
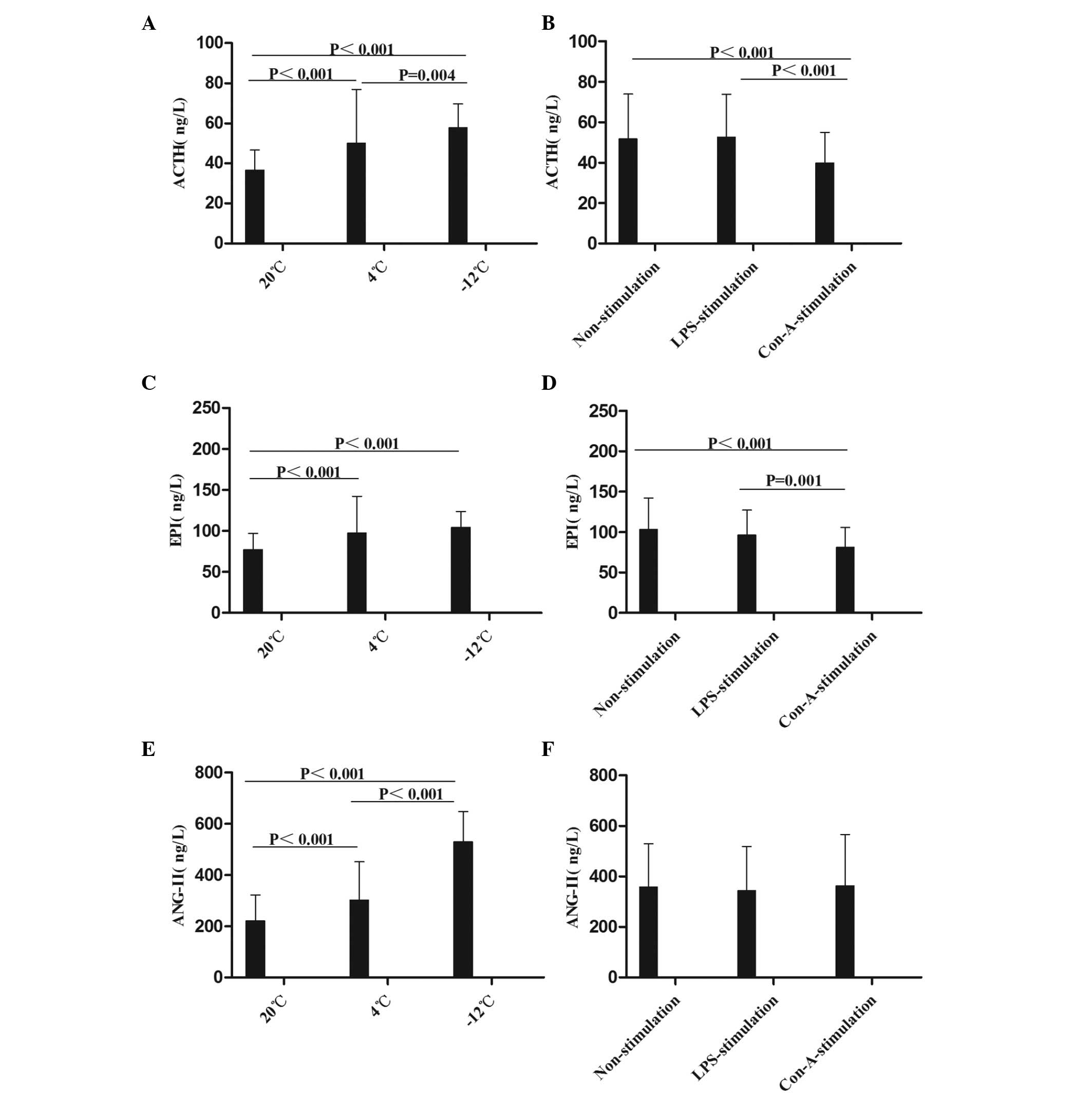 | Figure 1.Comparison of the mean levels of (A
and B) ACTH, (C and D) EPI and (E and F) ANG-II in the plasma at
different temperatures and with different stimulants. Data are
presented in ng/l as the mean ± standard deviation. (A and B)
Analysis of the ACTH data revealed the following: Comparison among
multiple temperatures, F (2,230)=28.714, P<0.001; comparison
among multiple stimulants, F (2,230)=10.864, P<0.001; comparison
between temperatures and stimulants, F (4,230)=6.652, P<0.001.
(C and D) Analysis of the EPI data revealed the following:
Comparison among multiple temperatures, F (2,215)=20.685,
P<0.001; comparison among multiple stimulants, F (2,215)=8.663,
P<0.001; comparison between temperatures and stimulants, F
(4,215)=6.195, P<0.001. (E and F) Analysis of the ANG-II data
revealed the following: Comparison among multiple temperatures, F
(2,213)=150.164, P<0.001; comparison between temperatures and
stimulants: F (4,213)=5.971, P<0.001. ACTH, adrenocorticotropin;
EPI, epinephrine; ANG-II, angiotensin-II; LPS, lipopolysaccharide;
Con-A, concanavalin A. |
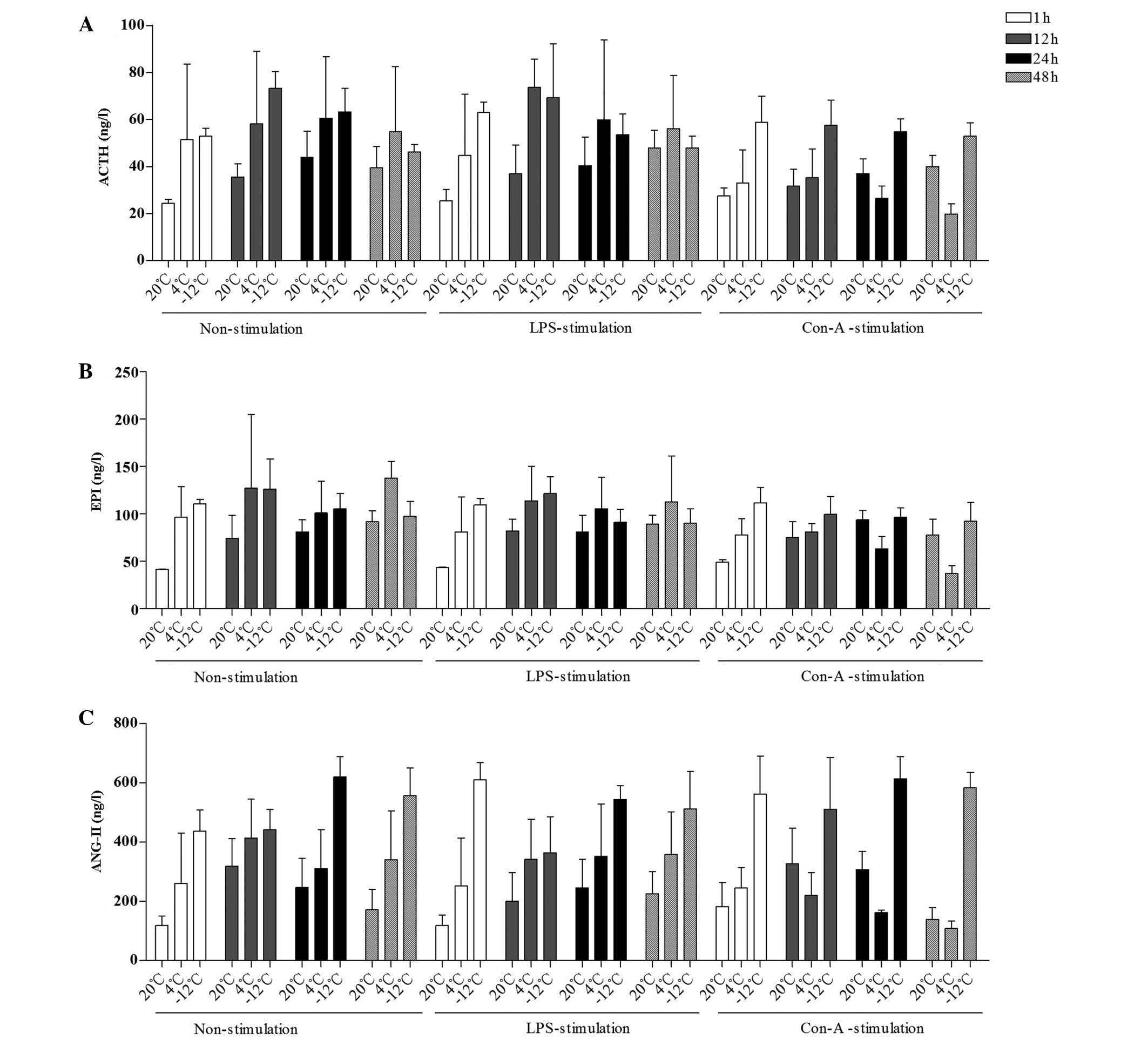 | Figure 2.Comparison of the mean levels of (A)
ACTH, (B) EPI and (C) ANG-II in the plasma at different times,
temperatures and with different stimulants. Data are presented in
ng/l as the mean ± standard deviation. (A) For the comparison of
the ACTH data among multiple times: F (3,230)=3.743, P=0.012; 1 h
vs. 12 h, P=0.002; 12 h vs. 48 h, P=0.014. For the comparison of
the ACTH data between temperatures and times: F (6,230)=2.612,
P=0.019. (B) For the comparison of the EPI data among multiple
times: F (3,215)=4.229, P=0.006; 1 h vs. 12 h, P=0.005; 12 h vs. 24
h, P=0.034. For the comparison of the EPI data between temperatures
and times: F (6,215)=3.759, P=0.002. (C) For the comparison of the
ANG-II data among multiple times: F (3,213)=3.181, P=0.025; 1 h vs.
24 h, P=0.004; 24 h vs. 48 h, P=0.046. For the comparison of the
ANG-II data between temperatures and times: F (6,213)=5.103,
P<0.001. For the comparison of the ANG-II data between times and
stimulants: F (6,213)=2.297, P=0.037. ACTH, adrenocorticotropin;
EPI, epinephrine; ANG-II, angiotensin-II; LPS, lipopolysaccharide;
Con-A, concanavalin A. |
In summary, it was found in the present study that
the levels of ACTH, EPI and ANG-II in the rat plasma were increased
following cold stress, which was consistent with the results
described by Ablimit et al (8), Belay and Woart (9), Vernikos et al (10), Yang et al (11) and Israel et al (12). The data suggest that cold stress
activates the hypothalamic-pituitary-adrenocortical (HPA) axis
(ACTH), the adrenomedullary hormonal system (AHS) (EPI) and the
renin-angiotensin-aldosterone system (RAAS) (ANG-II), which may
suppress the immunity of rats.
Effect of different temperatures and
stimulants on levels of T-lymphocyte cytokines in the rat
plasma
The mean levels of IL-2, IL-4, IFN-γ and IL-10 were
compared at different times, temperatures, and in response to
various stimulants (Figs. 3 and
4). Compared with the values at
20°C, the level of IL-2 in the rat plasma was decreased at 24 h in
the 4°C non-stimulation and LPS-stimulation groups, at the 24–48-h
time-points in the −12°C non-stimulation and LPS-stimulation
groups, at the 1–48-h time-points in the 4°C Con-A-stimulation
group and at the 12–48-h time-points in the −12°C Con-A-stimulation
group (Figs. 3A and B and 4A). With regard to the level of IL-4 in the
rat plasma, comparisons with the results at 20°C revealed decreases
at the 12–48-h time-points in the 4 and −12°C non-stimulation and
LPS-stimulation groups, at the 1–24-h time-points in the 4°C
Con-A-stimulation group and at 12 h in the −12°C Con-A-stimulation
group (Figs. 3C and D and 4B). The level of IFN-γ in the rat plasma
was decreased at the 12–24-h time-points in the 4°C non-stimulation
and LPS-stimulation groups, at 1, 24 and 48 h in the −12°C
non-stimulation and LPS-stimulation groups, at the 12–24-h
time-points in the 4°C Con-A-stimulation group and at the 1–12-h
time-points in the −12°C Con-A-stimulation group compared with the
results at 20°C (Figs. 3E and F and
4C). By contrast, the level of IL-10
in the rat plasma was increased at the 1–48-h time-points in the 4
and −12°C non-stimulation and LPS-stimulation groups and at the
1–48-h time-points in the −12°C Con-A-stimulation group compared
with the values at 20°C (Figs. 3G and
H and 4D).
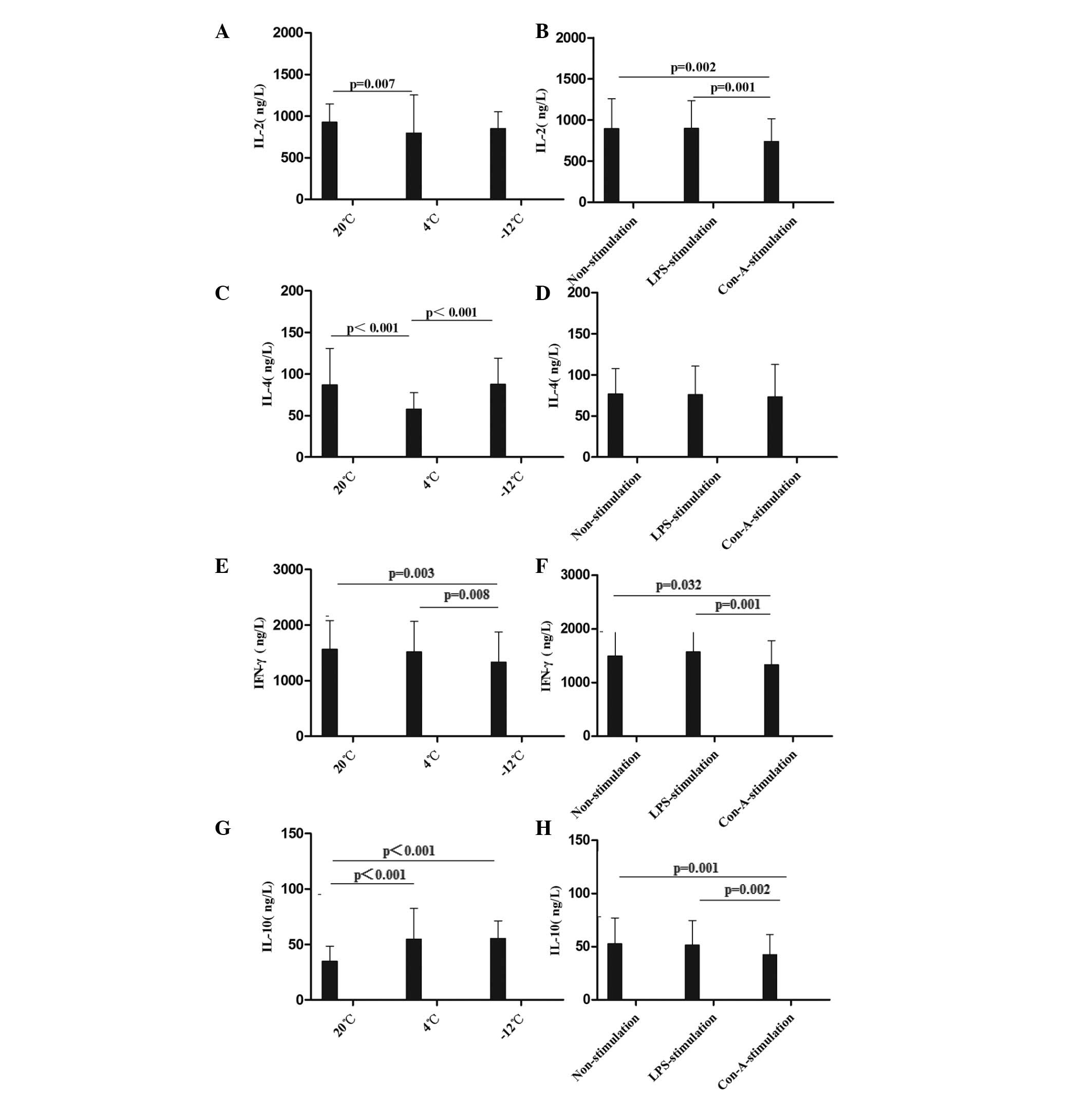 | Figure 3.Comparison of the mean levels of (A
and B) IL-2, (C and D) IL-4, (E and F) IFN-γ and (G and H) IL-10 in
the plasma at different temperatures and with different stimulants.
Data are presented in ng/l as the mean ± standard deviation. (A and
B) Analysis of the IL-2 data revealed the following: Comparison
among multiple temperatures, F (2,229)=5.323, P=0.006; comparison
among multiple stimulants: F (2,229)=4.603, P=0.011; comparison
between temperatures and stimulants, F (4,229)=7.796, P<0.001.
(C and D) Analysis of the IL-4 data revealed the following:
Comparison among multiple temperatures, F (2,228)=33.255,
P<0.001. (E and F) Analysis of the IFN-γ data revealed the
following: Comparison among multiple temperatures: F (2,229)=3.624,
P=0.029; comparison among multiple stimulants: F (2,229)=4.242,
P=0.016. (G and H) Analysis of the IL-10 data revealed the
following: Comparison among multiple temperatures: F
(2,230)=27.150, P<0.001; comparison among multiple stimulants: F
(2,230)=4.732, P=0.010; comparison between temperatures and
stimulants: F (4,230)=5.710, P<0.001. IL, interleukin; IFN-γ,
interferon-γ; LPS, lipopolysaccharide; Con-A, concanavalin A. |
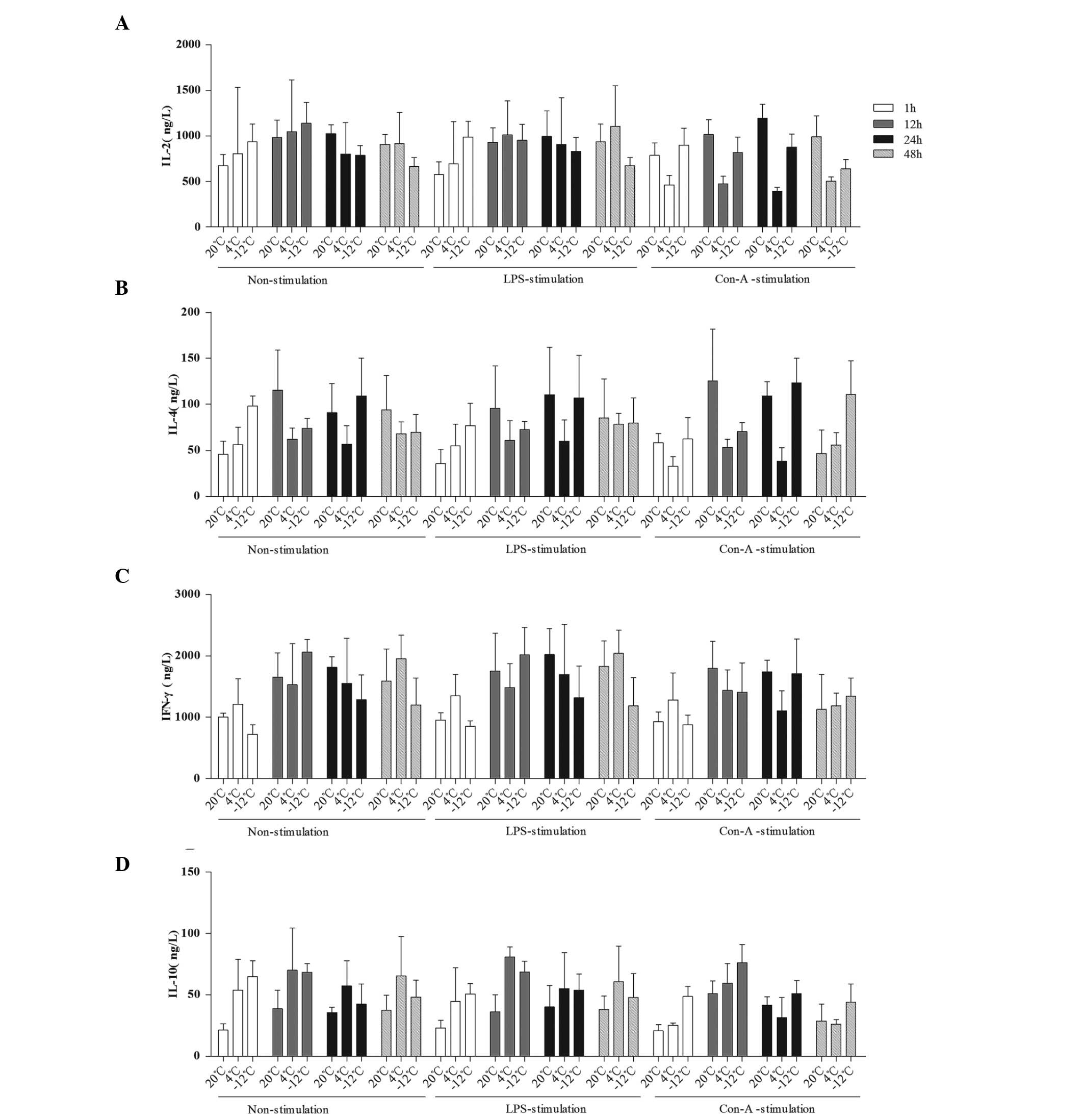 | Figure 4.Comparison of the mean levels of (A)
IL-2, (B) IL-4, (C) IFN-γ and (D) IL-10 in the plasma at different
times, temperatures and with different stimulants. Data are
presented in ng/l as the mean ± standard deviation. (A) For the
comparison of the IL-2 data among multiple times: F (3,229)=3.208,
P=0.024; 1 h vs. 12 h, P=0.003. For the comparison of the IL-2 data
between temperatures and times: F (6,229)=3.979, P=0.001. (B) For
the comparison of the IL-4 data among multiple times: F
(3,228)=12.828, P<0.001; 1 h vs. 12 h, P<0.001; 1 h vs. 24 h,
P<0.001; 1 h vs. 48 h, P=0.001; 24 h vs. 48 h, P=0.044. For the
comparison of the IL-4 data between temperatures and times: F
(6,228)=9.068, P<0.001. For the comparison of the IL-4 data
among temperatures, times and stimulants: F (12,228)=2.130,
P=0.017. (C) For the comparison of the IFN-γ data among multiple
times: F (3,229)=22.894, P<0.001; 1 h vs. 12 h, P<0.001; 1 h
vs. 24 h, P<0.001; 1 h vs. 48 h, P<0.001. For the comparison
of the IFN-γ data between temperatures and times: F (6,230)=5.445,
P<0.001. (D) For the comparison of the IL-10 data among multiple
times: F (3,230)=15.342, P<0.001; 1 h vs. 12 h, P<0.001; 1 h
vs. 24 h, P<0.001; 12 h vs. 24 h, P<0.001; 12 h vs. 48 h,
P<0.001. For the comparison of the IL-10 data between
temperatures and times: F (6,230)=2.326, P=0.034. IL, interleukin;
IFN-γ, interferon-γ; LPS, lipopolysaccharide; Con-A, concanavalin
A. |
In summary, it was found in the present study that
the levels of IL-2, IL-4 and IFN-γ in the rat plasma were decreased
and the level of IL-10 in the rat plasma was increased following
cold stress; these immunosuppressive states were not improved using
LPS and Con-A stimulation. These results were similar to those
described by Liu et al (7),
Belay and Woart (9), Shu et
al (13) and Aviles and Monroy
(14). The data suggest that rats
are induced to a state of autoimmune response by cold stress.
Effect of different temperatures and
stimulants on the expression of TLR4+ MNCs in the rat
spleen
The mean levels of TLR4+ MNCs in the
spleen were compared at different times, temperatures, and in
response to various stimulants (Figs.
5 and 6). Compared with the
results at 20°C, the levels of TLR4+ MNCs in the spleen
were decreased at 12 and 48 h in the 4°C non-stimulation group, at
1, 12 and 48 h in the −12°C non-stimulation group, at the 12–48-h
time-points in the 4°C LPS-stimulation group, at the 1–48-h
time-points in the −12°C LPS-stimulation group and at 1, 12 and 48
h in the 4 and −12°C Con-A-stimulation groups (Figs. 5A and B and 6A). Greater temperature decreases resulted
in more evident decreases in the TLR4 expression rate. These data
demonstrate that TLR4 expression in immunocompetent cells in the
spleen was decreased when EPI and IL-10 expression increased, and
that the innate immunity function in rats was damaged following
cold stress.
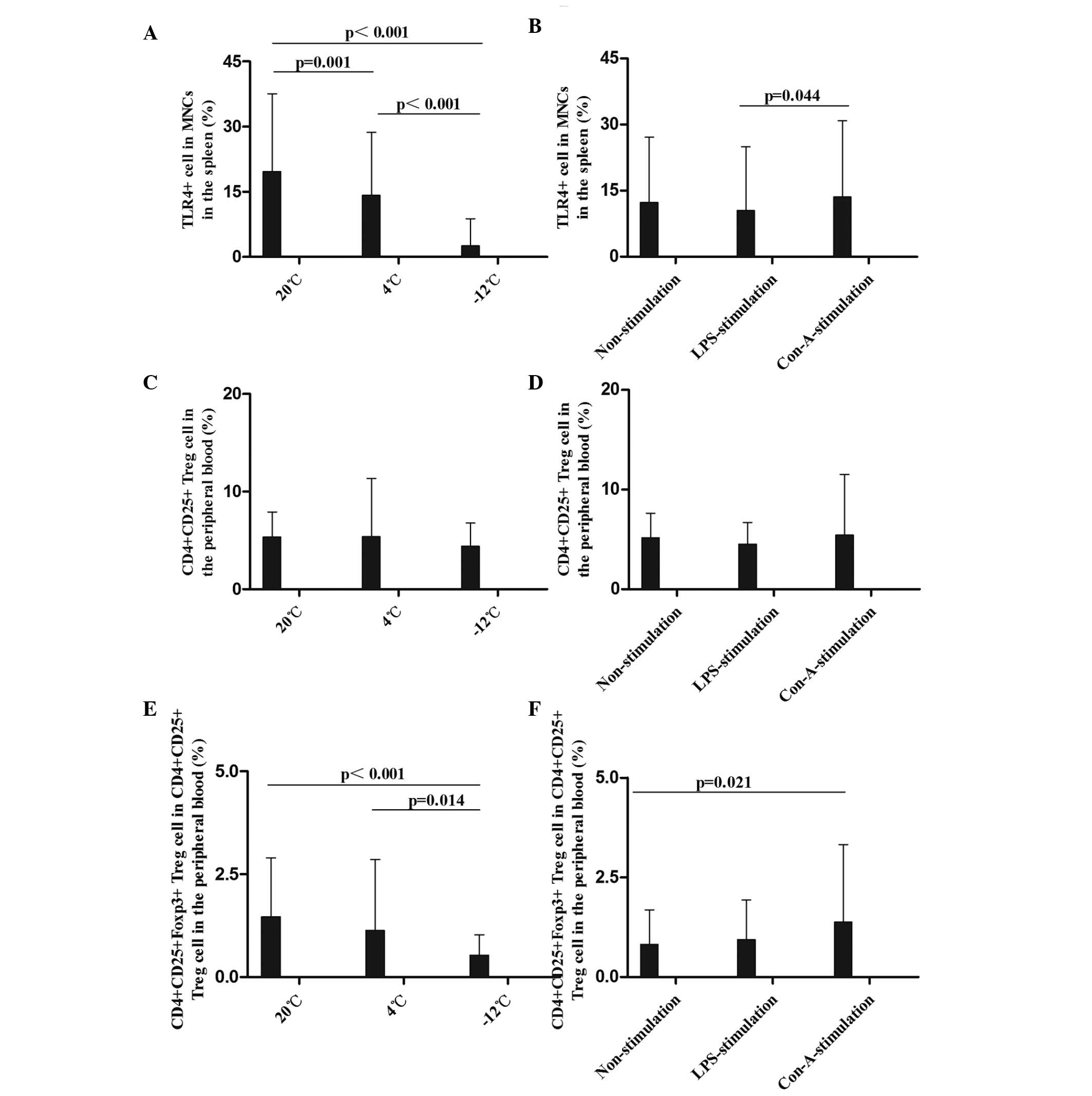 | Figure 5.Comparison of the mean levels of (A
and B) TLR4+ MNCs in the spleen, (C and D)
CD4+CD25+ Treg cells and (E and F)
CD4+CD25+Foxp3+ Treg cells in the
peripheral blood at different temperatures and with different
stimulants. Data (%) are presented as the mean ± standard
deviation. (A and B) Analysis of the TLR4+ MNC data
revealed the following: Comparison among multiple temperatures, F
(2,108)=67.199, P<0.001; comparison among multiple stimulants, F
(2,108)=2.129, P=0.126; LPS vs. Con-A, P=0.044. (C and D) Analysis
of the CD4+CD25+ Treg cell data revealed the
following: Comparison among multiple temperatures, F (2,108)=0.754,
P=0.474; comparison among multiple stimulants, F (2,108)=0.532,
P=0.589. (E and F) Analysis of the
CD4+CD25+Foxp3+ Treg cell data
revealed the following: Comparison among multiple temperatures, F
(2,108)=7.984, P=0.001; comparison between temperatures and
stimulants, F (4,108)=1.907, P=0.118; comparison between 20°C and
−12°C, P<0.001; comparison between 4°C and −12°C, P=0.014. TLR,
Toll-like receptor 4; CD, cluster of differentiation; Foxp3,
Forkhead box P3; MNC, mononuclear cell; Treg, regulatory T; LPS,
lipopolysaccharide; Con-A, concanavalin A. |
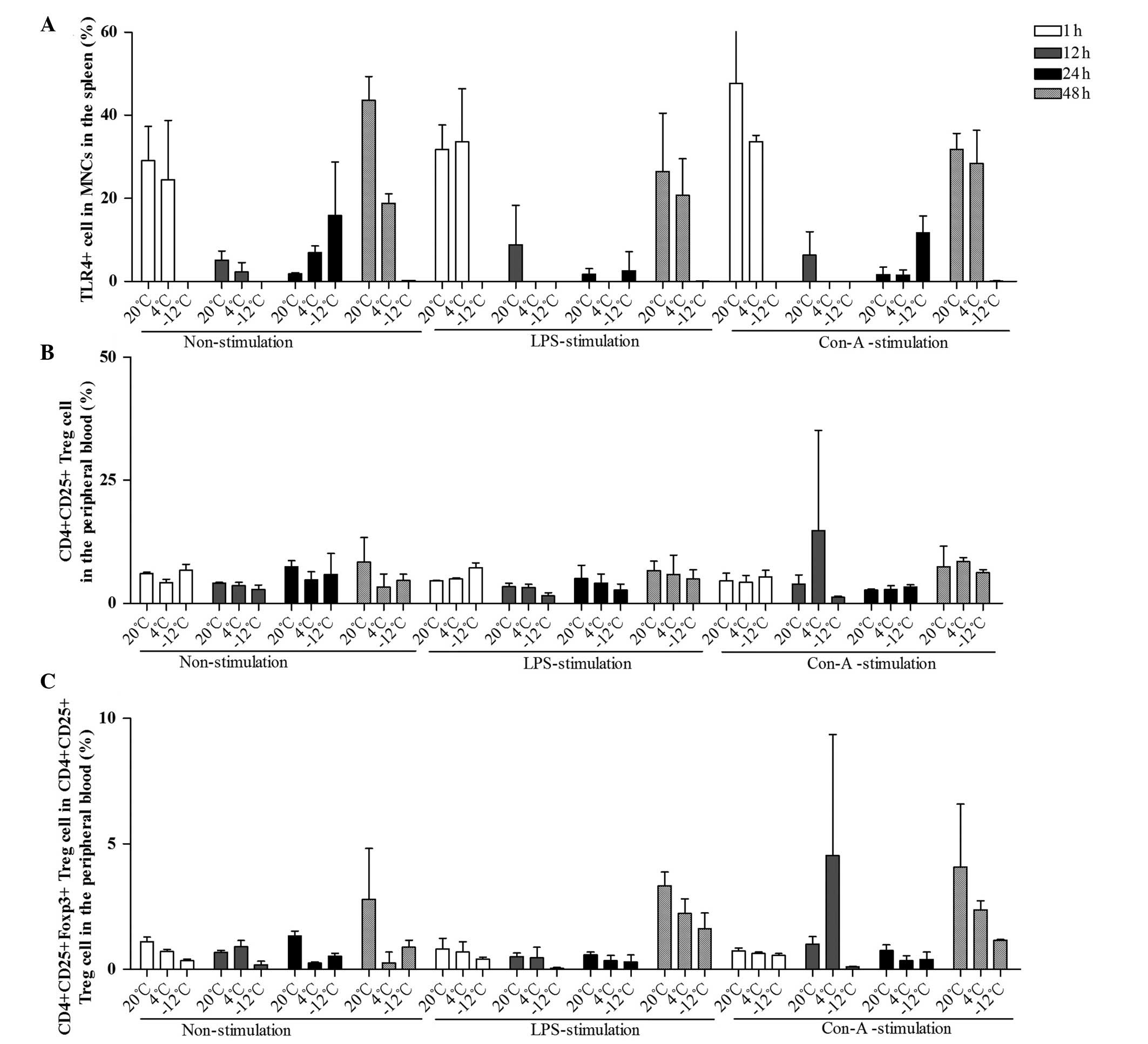 | Figure 6.Comparison of the mean levels of (A)
TLR4+ MNCs in the spleen, (B)
CD4+CD25+ Treg cells and (C)
CD4+CD25+Foxp3+ Treg cells in the
peripheral blood at different times, temperatures and with
different stimulants. Data (%) are presented as the mean ± standard
deviation. (A) For the comparison of the TLR4+ data
among multiple times: F (3,108)=64.432, P<0.001; 1 h vs. 12 h,
P<0.001; 1 h vs. 24 h, P<0.001; 12 h vs. 48 h, P<0.001; 24
h vs. 48 h, P<0.001. For the comparison of the TLR4+
data between temperatures and times: F (6,108)=29.595, P<0.001.
For the comparison of the TLR4+ data between times and
stimulants: F (6,108)=2.281, P=0.045. For the comparison of the
TLR4+ data among temperatures, times and stimulants: F
(12,228)=1.954, P=0.042. (B) For the comparison of the
CD4+CD25+ Treg data among multiple times: F
(3,108)=1.568, P=0.205. (C) For the comparison of the
CD4+CD25+Foxp3+ Treg cell data
among multiple times: F (3,108)=13.322, P<0.001; 1 h vs. 48 h,
P<0.001; 12 h vs. 48 h, P<0.001; 24 h vs. 48 h, P<0.001.
For the comparison of the
CD4+CD25+Foxp3+ Treg cell data
between temperatures and times: F (6,108)=4.434, P=0.001. For the
comparison of the CD4+CD25+Foxp3+
Treg cell data between times and stimulants: F (6,108)=2.397,
P=0.036). TLR, Toll-like receptor 4; CD, cluster of
differentiation; Foxp3, Forkhead box P3; MNC, mononuclear cell;
Treg, regulatory T; LPS, lipopolysaccharide; Con-A, concanavalin
A. |
In summary, the results of the present study showed
that the TLR4 expression rates in the MNCs in the rat spleen were
decreased following cold stress, and were not improved by LPS and
Con-A stimulation. This suggests that the innate immunity of rats
is suppressed by cold stress; consistently, TLR4 mutant mice have
been shown to be more susceptible to pulmonary tuberculosis and
enhanced mycobacterial outgrowth (4).
Effect of different temperatures and
stimulants on Treg cells in the peripheral blood in rats
The mean levels of CD4+CD25+
and CD4+CD25+Foxp3+ Treg cells in
the peripheral blood were compared at different times,
temperatures, and in response to various stimulants (Figs. 5 and 6). CD4+CD25+ Treg
cell rates in the peripheral blood were decreased at the 1–48-h
time-points at 4°C and at the 12–48-h time-points at −12°C in the
non-stimulation group; at the 12–48-h time-points in the −12°C
LPS-stimulation group; and at 1 h in the −12°C Con-A-stimulation
group, compared with the results at 20°C (Figs. 5C and D and 6B). With regard to the percentage of
CD4+CD25+Foxp3+ Treg cells,
significant decreases were observed at 1, 24 and 48 h at 4°C and at
the 1–48-h time-points at −12°C in the non-stimulation,
LPS-stimulation and Con-A-stimulation groups, particularly at −12°C
(Figs. 5E and F and 6C). Greater temperature decreases resulted
in more marked decreases in the percentage of
CD4+CD25+Foxp3+ Treg cells.
To the best of our knowledge, the results of the
present study have demonstrated for the first time that the
percentage of CD4+CD25+Foxp3+ Treg
cells was significantly decreased in the rat peripheral blood
following cold stress, which suggests that the adaptive immunity
and immunoregulation of rats are disturbed by cold stress. In a
previous study by Kelley et al (6), it was shown that cold exposure
increased the DTH reaction by 42% in calves.
Discussion
It has been shown in a number of studies that the
HPA axis, AHS and RAAS are activated in animals exposed to cold
stress (8–12); however, T-helper 1 (Th1) cytokine
(INF-γ) production is suppressed in animals exposed to cold stress
(11,15). In experimental autoimmune
encephalomyelitis mice, inflammatory foci of the central nervous
system were decreased and splenic
CD4+CD25+Foxp3+ Treg cells were
doubled following ACTH oral administration (16). ANG-II caused a 43% decrease in the
number of Foxp3+ cells in the renal cortex compared with
the control (17). The present study
also demonstrated that ACTH, EPI and ANG-II levels were increased
in rats following cold stress, which suggests that cold stress
excites the HPA axis, the AHS and the RAAS, which may be a cause of
immune disorders in rats suddenly exposed to the cold.
The TLR4 expressed on immunocompetent cells not only
participates in phagocytizing bacteria, viruses and protozoans as
part of the innate immunity to protect the body, but also mediates
the maturation and activation of antigen-presenting cells and the
proliferation, differentiation and development of T and B
lymphocytes (18,19) as part of the adaptive immune response
to protect the body. Du et al (20) demonstrated that the stress hormone
EPI downregulated TLR4 mRNA expression in macrophages in a time-
and dose-dependent manner. In the present study, a reduction in the
TLR4+ MNC rates in the spleen in rats exposed to cold
stress was observed. These findings suggest that innate
immunosuppression may be due to an increase in stress hormones and
a decrease in the TLR4 expression of immunocompetent cells in rats
exposed to the cold. These data are consistent with the observed
sudden increase in the morbidity of upper respiratory tract
infections following sudden environmental temperature decreases
(1).
It has been found in previous studies that the IL-2
level was decreased, even following Con-A and LPS stimulation, and
the corticosterone level was increased in rats exposed to acute and
chronic cold stress (7,13). In addition, mice subjected to cold
stress exhibited decreased IL-4 and IFN-γ production and an
increased IL-10 level (9,14). The results of the present study also
demonstrated that the levels of IL-2, IL-4 and IFN-γ were decreased
and that the level of IL-10 was increased in rats exposed to cold
stress, even with LPS and Con-A stimulation, compared with the
findings at 20°C. These results suggest that cold stress not only
excites ACTH, EPI and ANG-II production, but also alters the
balance of immunity and decreases the innate and adaptive immunity
in rats. Previous studies have found that ACTH inhibits
Con-A-stimulated T-lymphocyte mitogenesis (21) and decreases IL-2 and IFN-γ production
in mice (16). Furthermore, EPI
potently stimulates IL-10 production (22), and IL-10 has been shown to enhance
corticotropin-releasing factor and ACTH production in hypothalamic
and pituitary tissues (23).
Cold stress affects the host adaptive immune system.
Kelley et al (6) demonstrated
that cold exposure increased the DTH reaction by 42% in calves,
which suggests that the immunosuppression ability declines when
calves are exposed to cold stress. Foxp3 expressed on
CD4+CD25+ Treg cells is an important marker
of suppressive Treg cells (24).
Although ACTH significantly decreases CD4+ T-cell
counts, immunoglobulin levels did not change. In a previous study,
the splenic CD4+CD25+Foxp3+ Treg
frequency doubled in mice fed with ACTH compared with control mice
(16). Blocking ANG-II production
with angiotensin-converting enzyme inhibitors or inhibiting ANG-II
signaling with AT(1) receptor
blockers suppressed autoreactive Th1 and Th17 cells and promoted
antigen-specific CD4+Foxp3+ Treg cell
production (25). In the present
study it was demonstrated that the percentage of
CD4+CD25+Foxp3+ Treg cells was
significantly decreased following cold stress, and that greater
temperature decreases resulted in more evident decreases in the
percentage of CD4+CD25+Foxp3+ Treg
cells, which suggests that sudden cold stress facilitates the
development and occurrence of autoimmune disease.
Acknowledgements
This study was supported by grants from the
‘Eleventh Five-year Plan’ and support plans of the National
Ministry of Science and Technology (no. 2008 Bai68b01).
References
|
1
|
Smith AP: Effects of upper respiratory
tract illnesses and stress on alertness and reaction time.
Psychoneuroendocrinology. 38:2003–2009. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Temajo NO and Howard N: The mosaic of
environment involvement in autoimmunity: the abrogation of viral
latency by stress, a non-infectious environmental agent, is an
intrinsic prerequisite prelude before viruses can rank as
infectious environmental agents that trigger autoimmune diseases.
Autoimmun Rev. 13:635–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baccan GC, Oliveira RD and Mantovani B:
Stress and immunological phagocytosis: possible nongenomic action
of corticosterone. Life Sci. 75:1357–1368. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Branger J, Leemans JC, Florquin S, Weijer
S, Speelman P and Van Der Poll T: Toll-like receptor 4 plays a
protective role in pulmonary tuberculosis in mice. Int Immunol.
16:509–516. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zieziulewicz TJ, Mondal TK, Gao D and
Lawrence DA: Stress-induced effects, which inhibit host defenses,
alter leukocyte trafficking. Cell Stress Chaperones. 18:279–291.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kelley KW, Greenfield RE, Evermann JF,
Parish SM and Perryman LE: Delayed-type hypersensitivity, contact
sensitivity, and phytohemagglutinin skin-test responses of heat-
and cold-stressed calves. Am J Vet Res. 43:775–779. 1982.PubMed/NCBI
|
|
7
|
Liu YL, Bi H, Chi SM, Fan R, Wang YM, Ma
XL, Chen YM, Luo WJ, Pei JM and Chen JY: The effect of compound
nutrients on stress-induced changes in serum IL-2, IL-6 and
TNF-alpha levels in rats. Cytokine. 37:14–21. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ablimit A, Kühnel H, Strasser A and Upur
H: Abnormal Savda syndrome: long-term consequences of emotional and
physical stress on endocrine and immune activities in an animal
model. Chin J Integr Med. 19:603–609. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Belay T and Woart A: Cold-induced stress
increases the intensity of Chlamydia genital infection in mice. J
Microbiol Immunol Infect. 46:330–337. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vernikos J, Dallman MF, Bonner C, Katzen A
and Shinsako J: Pituitary-adrenal function in rats chronically
exposed to cold. Endocrinology. 110:413–420. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang G, Xi ZX, Wan Y, Wang H and Bi G:
Changes in circulating and tissue angiotensin II during acute and
chronic stress. Biol Signals. 2:166–172. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Israel A, Zavala LE, Cierco M, Gutierrez A
and Garrido Mdel R: Effect of AT(1) angiotensin II receptor
antagonists on the sympathetic response to a cold pressor test in
healthy volunteers. Am J Ther. 14:183–188. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shu J, Stevenson JR and Zhou X: Modulation
of cellular immune responses by cold water swim stress in the rat.
Dev Comp Immunol. 17:357–371. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Aviles H and Monroy FP: Immunomodulatory
effects of cold stress on mice infected intraperitoneally with a
50% lethal dose of Toxoplasma gondii. Neuroimmunomodulation.
9:6–12. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goldstein DS and Kopin IJ:
Adrenomedullary, adrenocortical, and sympathoneural responses to
stressors: a meta-analysis. Endocr Regul. 42:111–119.
2008.PubMed/NCBI
|
|
16
|
Brod SA and Hood ZM: Ingested (oral) ACTH
inhibits EAE. J Neuroimmunol. 232:131–135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barhoumi T, Kasal DA, Li MW, Shbat L,
Laurant P, Neves MF, Paradis P and Schiffrin EL: T regulatory
lymphocytes prevent angiotensin II-induced hypertension and
vascular injury. Hypertension. 57:469–476. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dabbagh K, Dahl ME, Stepick-Biek P and
Lewis DB: Toll-like receptor 4 is required for optimal development
of Th2 immune responses: role of dendritic cells. J Immunol.
168:4524–4530. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hayashi EA, Akira S and Nobrega A: Role of
TLR in B cell development: signaling through TLR4 promotes B cell
maturation and is inhibited by TLR2. J Immunol. 174:6639–6647.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Du Q, Min S, Chen LY, Ma YD, Guo XL, Wang
Z and Wang ZG: Major stress hormones suppress the response of
macrophages through down-regulation of TLR2 and TLR4. J Surg Res.
173:354–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Johnson EW, Hughes TK Jr and Smith EM:
ACTH enhancement of T-lymphocyte cytotoxic responses. Cell Mol
Neurobiol. 25:743–757. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smith EM, Cadet P, Stefano GB, Opp MR and
Hughes TK Jr: IL-10 as a mediator in the HPA axis and brain. J
Neuroimmunol. 100:140–148. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Elenkov IJ, Kvetnansky R, Hashiramoto A,
Bakalov VK, Link AA, Zachman K, Crane M, Jezova D, Rovensky J,
Dimitrov MA, Gold PW, Bonini S, Fleisher T, Chrousos GP and Wilder
RL: Low-versus high-baseline epinephrine output shapes opposite
innate cytokine profiles: presence of Lewis- and Fischer-like
neurohormonal immune phenotypes in humans? J Immunol.
181:1737–1745. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim JM and Rudensky A: The role of the
transcription factor Foxp3 in the development of regulatory T
cells. Immunol Rev. 212:86–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Platten M, Youssef S, Hur EM, Ho PP, Han
MH, Lanz TV, Phillips LK, Goldstein MJ, Bhat R, Raine CS, Sobel RA
and Steinman L: Blocking angiotensin-converting enzyme induces
potent regulatory T cells and modulates TH1- and TH17-mediated
autoimmunity. Proc Natl Acad Sci USA. 106:14948–14953. 2009.
View Article : Google Scholar : PubMed/NCBI
|














