Introduction
Toll-like receptors (TLRs) are pattern recognition
receptors that have important functions within innate immunity.
TLRs can trigger a number of cellular processes, including the
translocation of transcription factors, cytokine expression and the
activation of protein kinases, through the recognition of conserved
pathogen-associated molecular patterns, which ultimately lead to
the induction of inflammatory responses (1). In total, 11 human TLRs have been
identified, with the functions of human TLR-1–9 confirmed. Among
the TLR members, TLR-1, −2, −4, −5 and −6 are located on the cell
surface and are designed for the engagement of extracellular
pathogens, while TLR-3, −7, −8 and −9 are located on the endosome
and recognize intracellular pathogen-derived products (2). TLRs play a critical role in defending
against invaded pathogens by recognizing pathogen components and
chemical synthesis. In addition, TLRs can protect the host from
self-immune responses through the recognition of endogenous ligands
released from damaged tissues (3).
TLR-5 has been demonstrated to play an important
role in the defense against bacterial infection by binding to
flagellin, the major protein found in bacterial flagella, which are
critical for bacterial survival (4).
TLR-5 is expressed on monocytes, dendritic cells, T lymphocytes,
natural killer cells and epithelial cells, and provides a useful
tool for the host to fight against the infection of flagellated
bacteria. The activation of TLR-5 induces the secretion of a number
of immune-associated molecules, including interleukin (IL)-2, IL-10
and IL-12, as well as type I interferons (IFNs) (5). However, a comprehensive list of the
cytokines and chemokines induced following stimulation with a TLR-5
agonist has yet to be established.
In the present study, peripheral blood mononuclear
cells (PBMCs) were isolated from healthy volunteers and stimulated
with the TLR-5 agonist, flagellin. Subsequently, quantitative
polymerase chain reaction (PCR) was performed to detect the
expression levels of cytokines and chemokines in the PBMCs
stimulated by flagellin. In addition, an antibody chip array was
used to analyze the secretion of immune molecules in the
supernatant of the flagellin-stimulated PBMCs.
Materials and methods
Isolation of human PBMCs and
stimulation with flagellin
In total, 5-ml blood samples were collected from
healthy individuals (2 males, aged 25 and 27 years), and PBMCs were
isolated using lymphocyte separation medium (MP Biomedicals, LLC,
Santa Ana, CA, USA). The PBMCs were subsequently washed twice with
phosphate-buffered saline. In total, 2×106 PBMCs were
maintained in 10% 1640 culture medium (Invitrogen, Carlsbad, CA,
USA) and stimulated with 100 ng/ml flagellin (IMG2205; Novus
Biologicals, LLC, Littleton, CO, USA), the TLR-5 agonist. The
present study was approved by the Ethics Committee of the West
China Hospital of Sichuan University (Chengdu, China). Written
informed consent was obtained from all the individuals prior to
their participation.
RNA extraction, reverse transcription
and quantitative PCR
At 4 h after stimulation, the flagellin-treated and
untreated control PBMCs were washed three times to remove any
remaining flagellin. The total RNA was extracted using a RNeasy
mini kit (Qiagen, Dusseldorf, Germany), and a ReverTra Ace reverse
transcription kit (FSQ-101; Toyobo Co., Ltd., Kagoshima, Japan) was
used to synthesize cDNA following the removal of genomic DNA by
DNaseI (RT411; Tiangen Biotech Co., Ltd., Beijing, China). The
conditions for the reverse transcription reaction were as follows:
65°C for 5 min, followed by 37°C for 15 min and 98°C for 5 min.
Quantitative PCR was performed using SYBR Green
RealMasterMix (FP202; Tiangen Biotech Co., Ltd.). The quantitative
PCR assay was performed in an iCycler iQTM Optical Module (Beckman
Coulter, Inc., Fullerton, CA, USA) under the following conditions:
Initial denaturation at 95°C for 30 sec, followed by 40 cycles of
denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec and
elongation at 72°C for 30 sec. A melting curve was constructed
between temperatures of 55–95°C, in 0.5°C increments and 10-sec
intervals. The primers used for quantitative PCR are shown in
Table I, and glyceraldehyde
3-phosphate dehydrogenase was used as an internal control.
Untreated PMBCs were used as a negative control, and all
experiments were performed in triplicate.
 | Table I.List of oligonucleotides used for
quantitative polymerase chain reaction detection. |
Table I.
List of oligonucleotides used for
quantitative polymerase chain reaction detection.
| Gene | Forward primer | Reverse primer | GenBank number |
|---|
| CCL5 |
GACACCACACCCTGCTGCT |
TACTCCTTGATGTGGGCACG | NM_002985 |
| CCL8 |
GTTTCTGCAGCGCTTCTGTG |
TGGCTGAGCAAGTCCCTGA | Y10802 |
| CCL17 |
CCATCGTTTTTGTAACTGTGCAG |
TGCATTCTTCACTCTCTTGTTGTTG | NM_002987 |
| CCL19 |
GGCACCAATGATGCTGAAGA |
GAAGTTCCTCACGATGTACCCAG | NM_006274 |
| CCL20 |
TCCTGGCTGCTTTGATGTCA |
TCAAAGTTGCTTGCTGCTTCTG | NM_004591 |
| CCL22 |
TGCGCGTGGTGAAACACT |
GGTTAGCAACACCACGCCA | NM_002990 |
| CCL26 |
CCAAGACCTGCTGCTTCCAA |
GAATTCATAGCTTCGCACCCA | NM_006072 |
| CCL28 |
CTCGCCATCGTGGCCTT |
GCAATGGGAAGTATGGCTTCTG | AF220210 |
| c-Myc |
CAAGACTCCAGCGCCTTCTC |
GTTGAGTAACGAGCTGACCCC | AM393287 |
| CTNNB |
CATCGTGAGGGCTTACTGGC |
GAGCAAGGCAACCATTTTCTG | XM_006712984 |
| CXCL2 |
AGGTGAAGTCCCCCGGAC |
GCCCATTCTTGAGTGTGGCT | NM_002089 |
| CXCL6 |
GCTGAGAGTAAACCCCAAAACG | GGAGCACTGCGGGCC | NM_002993 |
| CXCL12 |
GCCCGTCAGCCTGAGCTA |
GACGTTGGCTCTGGCAACAT | NM_000609 |
| GAPDH |
GAAGGTGAAGGTCGGAGTC |
GAAGATGGTGATGGGATTTC | J04038 |
| IFN-β |
CAGCAATTTTCAGTGTCAGAAGCT |
TCATCCTGTCCTTGAGGCAGT | M28622 |
| IFN-γ |
CCAACGCAAAGCAATACATGA |
CGCTTCCCTGTTTTAGCTGC | J00219 |
| IL-1β |
ACGAATCTCCGACCACCACT |
CCATGGCCACAACAACTGAC | M15330 |
| IL-2 |
CAAGAATCCCAAACTCACCAGG |
GACACTGAAGATGTTTCAGTTCTGT | J00264 |
| IL-6 |
GACCCAACCACAAATGCCA |
GTCATGTCCTGCAGCCACTG | M14584 |
| IL-8 |
CTGGCCGTGGCTCTCTTG |
CCTTGGCAAAACTGCACCTT | NM_000584 |
| IL-12 |
CGGTCATCTGCCGCAAA |
CAAGATGAGCTATAGTAGCGGTCCT | M65272 |
| IL-15 |
GACCCCACCAAAGCTGGAC |
TCACAGTGCTGCTGTCTGCTG | M90391 |
| IP-10 |
TGAAATTATTCCTGCAAGCCAA |
CAGACATCTCTTCTCACCCTTCTTT | NM_001565 |
| JNK |
GCTAATTCTGTACCAATGTC |
GAAGAGTGCACGTCAGGAAC | NM_139049 |
| MAPK |
CCTGCTCGGTGCACGATGCTG |
CTCTGTCTCTTCACGTGGCGG | NM_003954 |
| MEKK1 |
CTTTTAAGTCAGAAGTTGCTG |
CTTCTCCATTTTCAACCTGC | AF042838 |
| NF-κB |
AGAGTGCTGGAGTTCAGGATA |
AAGGTGGATGATTGCTAAGTGT | AJ271718 |
| P53 |
CTTGCATTCTGGGACAGCCAAG |
CACGCAAATTTCCTTCCACTCGG | DQ892492 |
| PTEN |
ACCATAACCCACCACAGC |
CAGTTCGTCCCTTTCCAG | NM_058074 |
| pSTAT3 |
CCTACAAAGGGGACCCCATTGTAC |
CAGGGAATTTGACCAGCAACC | NM_213662 |
| TGF-β |
TATCGACATGGAGCTGGTGAAG |
CAGCTTGGACAGGATCTGGC | X02812 |
| TNF-α |
GGTGCTTGTTCCTCAGCCTC |
CAGGCAGAAGAGCGTGGTG | M10988 |
| VEGF |
GACTTGAGTTGGGAGGGGAA |
GAGGCTCAGCGCCAGGGCTGGG | AF024710 |
Detection of secretory molecules using an antibody
chip array. At 4 h after stimulation with flagellin, the cell
supernatants were collected from the treated and untreated PBMCs. A
RayBio® Human Antibody Array (C series; RayBiotech, Inc., Norcross,
GA, USA) was applied for the screening of the secretory proteins,
according to the manufacturer's instructions.
Statistical analysis
Data obtained from the quantitative PCR assay were
analyzed using Bio-Rad iQ5 software (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The results are expressed as the mean ±
standard error of the mean, and were statistically analyzed by a
χ2 test using SPSS 17.0 software (SPSS, Inc., Chicago,
IL, USA). P<0.05 and P<0.001 were considered to indicate a
statistically significant difference. The figures were constructed
using GraphPad Prism 5 software (GraphPad Software, Inc., La Jolla,
CA, USA).
Results
Cytokine expression is induced by the
activation of TLR-5
Expression levels of cytokines were analyzed using
quantitative PCR, in order to determine the candidate molecules
that were important in the TLR-5-mediated biological functions of
PBMCs.
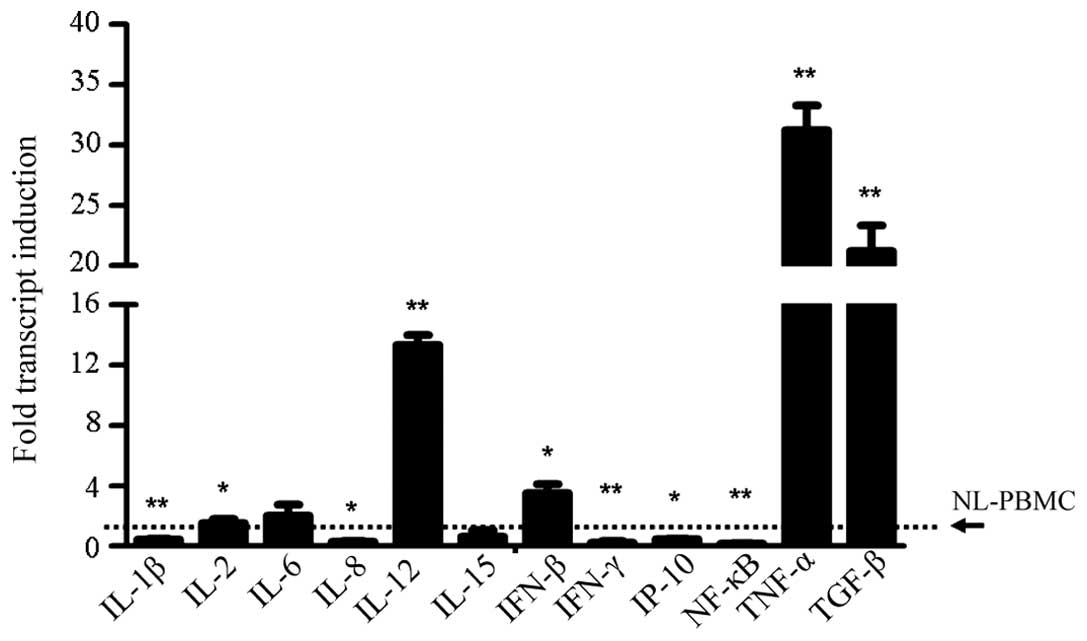
In the detection of cytokine expression, several
important molecules were found to be highly expressed in the
flagellin-stimulated PBMCs, which were IL-12 (P<0.001), tumor
necrosis factor-α (P<0.001) and transforming growth factor
(TGF)-β, as well as IL-2 and IFN-β (P<0.05; Fig. 1). By contrast, the expression levels
of IL-1β (P<0.001) and IFN-γ-induced protein (IP)-10 (P<0.05)
were inhibited upon TLR-5 activation, while the expression levels
of IL-6 and IL-15 were found to be unchanged. Notably, the
expression levels of IL-8 (P<0.05), IFN-γ (P<0.001) and
nuclear factor-κB (P<0.001) were markedly inhibited following
stimulation with the TLR-5 ligand (Fig.
1). These results were in contrast to a former study, which
demonstrated that increased expression levels of IL-8 and IFN-γ
upon pathogen stimulation were important in the immune response
against foreign invaders.
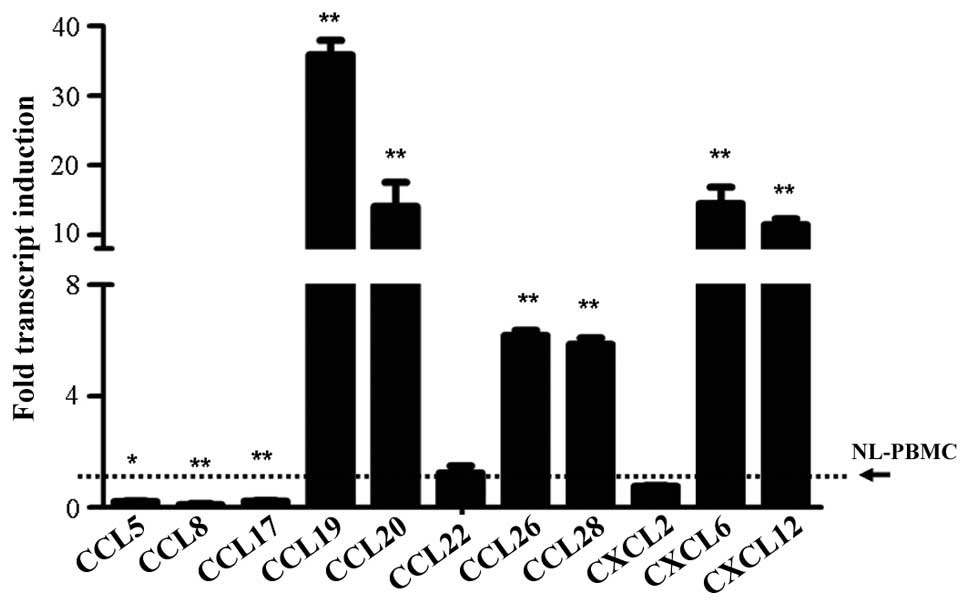
Variation in the expression levels of
chemokines stimulated by flagellin
In the study, the expression levels of a number of
important chemokines from the chemokine (C-C motif) ligand (CCL)
and chemokine (C-X-C) ligand (CXCL) families were detected. The
results demonstrated that flagellin stimulation significantly
increased the expression levels of CCL19 (P<0.001), CCL20
(P<0.001), CCL26 (P<0.001), CCL28 (P<0.001), CXCL6
(P<0.001) and CXCL12 (P<0.001) when compared with the
untreated cells (Fig. 2). However,
inhibited expression was observed for CCL5 (P<0.05), CCL8
(P<0.001) and CCL17 (P<0.001), and the expression levels of
CCL22 and CXCL2 were shown to remain at the same level as the
control cells (Fig. 2).
Detection of tumor-associated gene
expression upon TLR-5 activation
A number of molecules are known to play an important
role in tumor initiation. The molecules selected for the study
included the antitumor genes, P53 and phosphatase and tensin
homolog (PTEN), the oncogenes, c-Myc and proliferating cell nuclear
antigen (PCNA), as well as a number of important kinase signaling
molecules, including β-catenin (CTNNB), c-Jun N-terminal kinase
(JNK), mitogen-activated protein kinase (MAPK), mitogen-activated
protein kinase kinase kinase 1 and phosphorylated signal transducer
and activator of transcription (pSTAT3). The results indicated that
the expression levels of P53 (P<0.001) and PTEN (P<0.001)
were highly induced by flagellin, while c-Myc expression was
inhibited (P<0.05; Fig. 3).
However, the expression levels of PCNA and vascular endothelial
growth factor were not significantly influenced by TLR-5
activation. With regard to the kinase signaling molecules, induced
expression was observed for MAPK and pSTAT3, while CTNNB and JNK
expression levels were inhibited (Fig.
3).
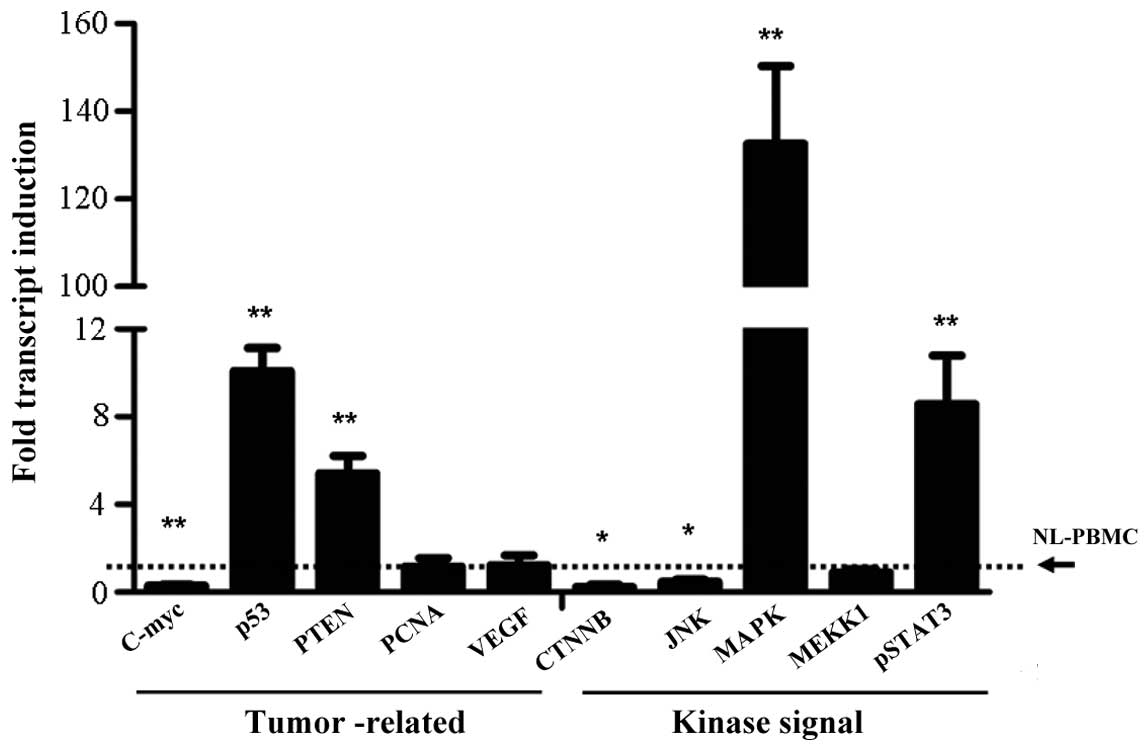 | Figure 3.Variation in the expression levels of
tumor-associated genes and kinase signaling molecules, as assessed
by quantitative polymerase chain reaction. **P<0.001 and
*P<0.05, vs. control group. In quantitative polymerase chain
reaction analysis, the level of NL-PBMC was set as the control
group and the expression level was set as 1. PTEN, phosphatase and
tensin homolog; PCNA, proliferating cell nuclear antigen; VEGF,
vascular endothelial growth factor; CTNNB, β-catenin; JNK, c-Jun
N-terminal kinase; MAPK, mitogen-activated protein kinase; MEKK,
mitogen-activated protein kinase kinase kinase; pSTAT,
phosphorylated signal transducer and activator of
transcription. |
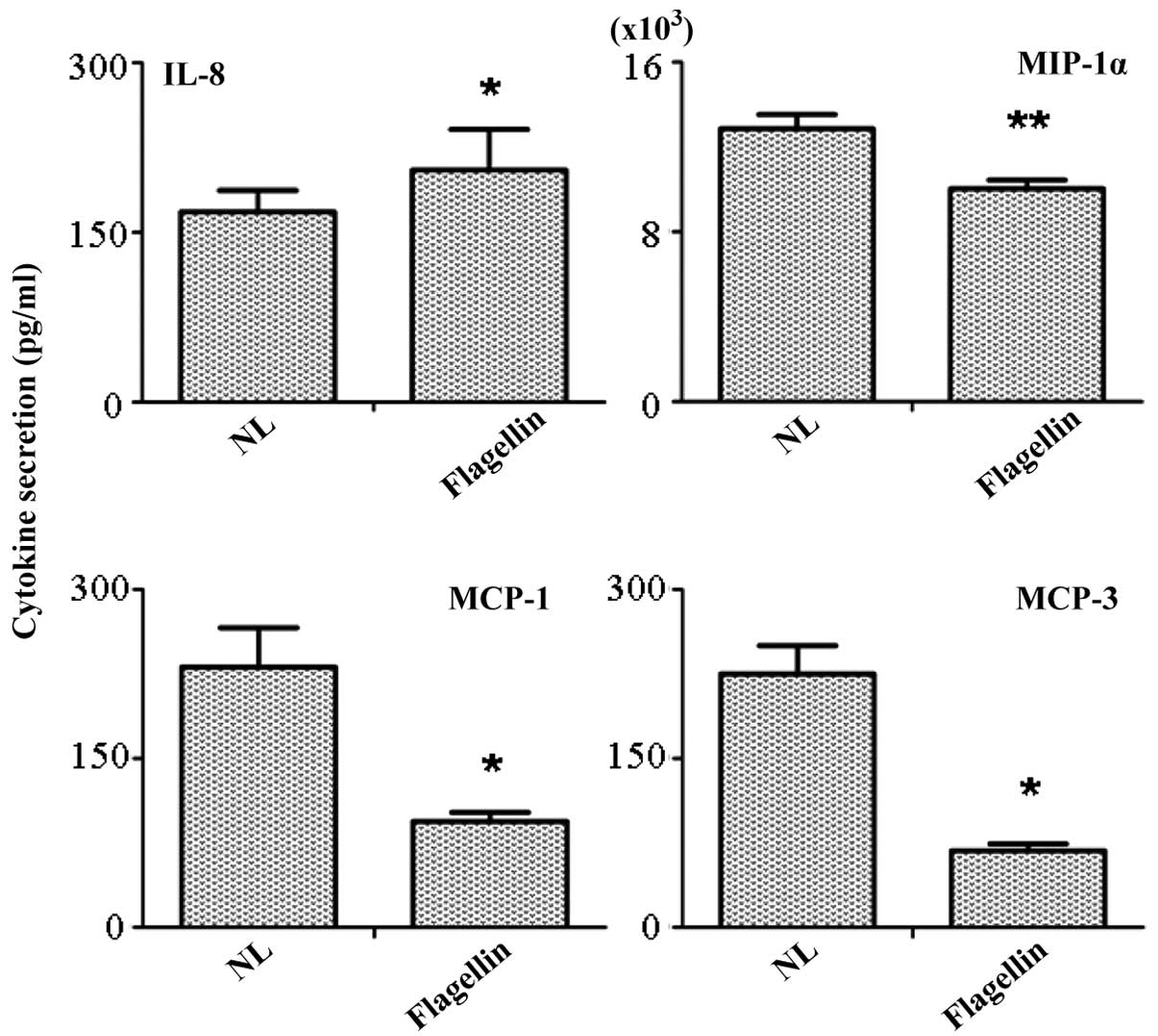
Antibody chip array of the secretory
molecules in the supernatant
Supernatants of the TLR-5-stimulated and
unstimulated PBMCs were collected, and the detection of the
secreted molecules was performed using a RayBio® antibody chip
array. In total, 20 important cytokines and chemokines were
included in the array. The results indicated that only four
molecules produced a positive response. Among these molecules, the
secretion of monocyte chemoattractant protein (MCP)-1 (P<0.05),
MCP-3 (P<0.05) and macrophage inflammatory protein-1α were
significantly inhibited (Fig. 4).
However, the secretion of IL-8 was identified in the supernatant,
which contradicted with the results from the quantitative PCR that
indicated that IL-8 expression was inhibited by flagellin
stimulation. An explanation for this phenomenon is that changes in
gene expression are not always consistent with those in protein
secretion.
Discussion
TLRs comprise a family of transmembrane proteins
that differentially recognize pathogens and initiate inflammatory
signaling pathways (6). An important
function of TLRs is their ability to bind to microorganisms by
recognizing conserved structures, which are known as
pathogen-associated molecular patterns (7). All members of the TLR family are able
to trigger a specific immune response by activating MYD88-mediated
pathways, with the exception of TLR-3, which subsequently induces a
number of immune molecules that are important in disease treatment
(8). TLR-5 has been shown to
function in the defense against invading microbes via the
recognition of flagellin on bacteria (9). A previous study demonstrated that
activation of TLR-5 increased the expression levels of IL-10 and
IL-12, while inhibiting IFN-γ expression (10).
In the present study, PBMCs collected from healthy
volunteers were treated with the TLR-5 ligand, flagellin.
Subsequently, the variation in the expression levels of immune
molecules was detected to determine which molecules were important
for the biological functions of PMBCs, including tumor initiation,
the inflammatory response and cell proliferation. The results
demonstrated that flagellin stimulation induced the expression of
IL-12, IFN-β and TGF-β, which were consistent with the results of a
previous study (11). The major
difference between the present and previous studies was that the
current study covered a wider range of molecules that are known to
be important in mediating the biological functions of PBMCs,
including molecules that influence the proinflammatory status,
genes that mediate the antitumor response and kinase signaling
molecules. The results of the present study confirmed that
activation of the TLR-5 pathway inhibited the expression of IL-1β,
IL-8 and IP-10, as well as the chemokines, CCL5, CCL8 and CCL17,
indicating the unique role of TLR-5 in regulating the functions of
PBMCs. In addition, the present study revealed increased expression
levels of the antitumor genes, P53 and PTEN, and the kinase
signaling molecules, MAPK and pSTAT3, in the TLR-5 stimulated
cells, indicating the potential role of TLR-5 in the antitumor
response.
Although the present study has demonstrated that a
number of interesting candidate molecules are involved in the
mechanism of TLR-5, a limitation of the study was the short
treatment time (4 h) of flagellin, which was selected as this is
the common stimulation time in the presence of a TLR agonist
(12). In addition, certain
important molecules exhibited no response upon ligand stimulation,
particularly in the antibody chip array, with only four molecules
showing a positive response in an array of >20 molecules.
Therefore, further study should aim to investigate the roles of the
candidate molecules, identified in the present study, in mediating
the biological functions of PBMCs.
References
|
1
|
Akira S, Uematsu S and Takeuchi O:
Pathogen recognition and innate immunity. Cell. 124:783–801. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takeda K, Kaisho T and Akira S: Toll-like
receptors. Annu Rev Immunol. 21:335–376. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ozinsky A, Underhill DM, Fontenot JD, et
al: The repertoire for pattern recognition of pathogens by the
innate immune system is defined by cooperation between Toll-like
receptors. Proc Natl Acad Sci USA. 97:13766–13771. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Iwasaki A and Medzhitov R: Regulation of
adaptive immunity by the innate immune system. Science.
327:291–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Merio A, Calcaterra C, Mènard S and
Balsari A: Cross-talk between Toll-like receptors 5 and 9 on
activation of human immune responses. J Leukoc Biol. 82:509–518.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Akira S and Takeda K: Toll-like receptor
signalling. Nat Rev Immunol. 4:499–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Beutler B: Inferences questions and
possibilities in Toll-like receptor signaling. Nature. 430:257–263.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Balander JM and Medzhitov R: Regulation of
phagosome maturation by signals from Toll-like receptors. Science.
304:1014–1018. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hawn TR, Verbon A, Letinga KD, et al: A
common dominant TLR5 stop codon polymorphism abolishes flagellin
signaling and is associated with susceptibility to legionnaires'
disease. J Exp Med. 198:1563–1572. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hayashi F, Smith KD, Ozinsky A, et al: The
innate immune response to bacterial flagellin is mediated by
Toll-like receptor 5. Nature. 410:1099–1103. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andersen-Nissen E, Smith KD, Strobe KL, et
al: Evasion of Toll-like receptor 5 by flagellated bacteria. Proc
Natl Acad Sci USA. 102:9247–9252. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin Y, Zhang L, Cai AX, Lee M, Zhang W,
Neuberg D, Canning CM, Soiffer RJ, Alyea EP, Ritz J, Hacohen N,
Means TK and Wu CJ: Effective posttransplant antitumor immunity is
associated with TLR-stimulating nucleic acid-immunoglobulin
complexes in humans. J Clin Invest. 121:1574–1584. 2011. View Article : Google Scholar : PubMed/NCBI
|