Introduction
Cognitive impairment is a common manifestation of a
variety of neurological disorders and is characterized by an
impairment of learning and memory function (1). The pathological process of cognitive
impairment is accompanied by aphasia, apraxia, agnosia and a number
of other changes in behavioral performance (2). Cognitive impairment may develop into
Alzheimer's disease, which detrimentally affects the quality of
life of patients, and increases the burden on their family and
society (1). At present, a clinical
diagnosis of cognitive impairment relies on neuropsychological
tests, blood tests, neuroimaging, electroencephalography and
cerebrospinal fluid tests (3). The
pathogenesis of cognitive impairment has yet to be fully elucidated
(4). Several widely accepted
pathological causes of the disease include the cerebral ischemia,
hypoxia and neuronal necrosis in brain tissue caused by trauma,
cerebral vascular inflammation, vascular stenosis and brain
embolism (5). In particular,
research concerning the disease pathogenesis has focused on
vascular lesions in the brain (6).
Vascular diseases of the brain may be caused by an
abnormality in the blood rheology that may also be involved in the
pathological process of other diseases (7). Plasminogen activator inhibitor-1
(PAI-1) is a serine protease inhibitor that inactivates the
plasminogen activators t-PA and u-PA, and inhibits intravascular
fibrinolysis that results in further changes in blood rheology and
an increased risk of ischemic injury (8). PAI-1 has been recognized as a risk
factor for the pathogenesis and development of ischemic
cerebrovascular and cardiovascular diseases (9,10). It
has been demonstrated that PAI-1 is targeted by micro RNA (miR)-30b
to regulate the proliferation and apoptosis of gastric cancer cells
(11). However, the role of PAI-1
and its upstream regulators in cognitive impairment has not been
fully established.
In the present study, a mouse model of cognitive
impairment was induced by scopolamine, and the role of PAI-1 in the
disease pathogenesis was investigated. Behavioral performance tests
were performed and assessed by the Morris water maze (MWM) test.
The mRNA and protein expression levels of PAI-1 were determined,
and the underlying mechanism concerning miR-30b was explored.
Materials and methods
Animal model
A total of 80 male Kunming mice (weight, 18–22 g;
age, 4 weeks), were purchased from Xinteng Bier Animal Co., Ltd.
(SCXK 2012-0008; Chongqing, China) and housed in cages at 22°C, 56%
humidity with a 12 h light/dark cycle and ad libitum access
to food and water. Each animal experimental procedure was approved
by the Care of Experimental Animals Committee of Laiwu Hospital
Affiliated to Taishan Medical University (Laiwu, China). The animal
cognitive impairment model was established by daily intraperitoneal
injection of scopolamine hydrobromide for 13 days (2 mg/kg body
weight; H19994038; Furen Medicines Group, Henan, China) for 13
days. The normal control group was injected with saline.
MWM test
The behavioral performance of the model mice was
assessed using the MWM test (Institute of Materia Medica, Chinese
Academy of Medical Science and Peking Union Medical College,
Beijing, China). The test commenced following the 13 day period of
scopolamine hydrobromide injections. The navigation test was
performed for 4 days, and each mouse was subjected to 4 tests
daily. The swimming pool was 0.8 m in diameter and divided into
four quadrants of equal size. The water was 30 cm in depth and a
hidden platform was placed 1 cm under the surface in the center of
one quadrant. A cardinal point of the quadrant without the platform
was randomly selected as the start location. The test commenced
when a mouse was placed in the pool, and ended when the mouse
located the platform. The escape latency was recorded as time that
the mouse spent searching for the platform. If a mouse failed to
locate the platform within 60 sec, the escape latency was recorded
as 60 sec and the mouse was guided to the platform. After remaining
on the platform for 15 sec, the next test began. On day 5 the probe
test was performed, in which the platform was removed and each
mouse was allowed to swim freely in the pool for 60 sec. The
swimming path length, as well as the number of times that each
mouse swam across the region where the platform used to be, was
recorded.
Tissue sample preparation
Blood samples were obtained following the behavioral
test. The mice were fasted for 12 h and anesthetized by
intraperitoneal injection of 10% chloral hydrate (0.0004,l/g body
weight; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China).
Abdominal aortic blood was collected and the mice were sacrificed
by cervical dislocation. The serum was separated and stored at
−80°C. The mouse brains were removed and the hippocampus was
separated. The hippocampal tissue was washed with 0.9% cold saline
and stored at −80°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The mRNA levels of PAI-1 and the levels of miRNA-30b
were detected in the blood samples using qPCR. Total RNA (2 µl)
from the hippocampus was extracted using TRIzol reagent (Yeasen
Corporation, Shanghai, China) and RNA in the blood samples was
extracted using the miRNeasy SerumPlasma kit (Guangzhou Jianlun
Biological Technology Co., Ltd., Guangzhou, China). Residual
genomic DNA was removed using DNase (Cloud-Clone Corporation,
Houston, TX, USA) Reverse transcription was performed to obtain
cDNA using the miRcute miRNA cDNA First-strand Synthesis kit
[Tiangen Biotech (Beijing) Co., Ltd., Beijing, China]. qPCR was
performed using the miRcute miRNA Quantitative Fluorescence
Detection kit [Tiangen Biotech (Beijing) Co., Ltd.] and PCR-iQ5
Multicolor Real-time PCR Detection System (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). The 25 µl PCR system consisted of 12.5 µl
SuperReal PreMix [Tiangen Biotech (Beijing) Co., Ltd.], 1 µl
primer, 2 µl cDNA template and 5 µl ddH2O. For PAI-1
detection, the primer sequences were as follows: PAI-1 forward,
5′-TCTCCGCCATCACCAACATT-3′ and reverse,
5′-GAGAGAACTTAGGCAGGATGAGG-3′; β-actin forward,
5′-AACCCTAAGGCCAACAGTGAAAAG-3′ and reverse,
5′-TCATGAGGTAGTCTGTGAGGT-3′. The reaction conditions consisted of
denaturation at 95°C for 2 min, 95°C for 30 sec, 58°C for 30 sec
and 72°C for 30 sec for a total of 40 cycles. For miR-30b
detection, the primer sequences were as follows: miR-30b forward,
5′-GCGCCTGTAAACATCCTACAC-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′; U6
forward, 5′-GCTTCGGCAGCACATATACTAA-3′ and reverse,
5′AACGCTTCACGAATTTGCGT-3′. Primers were synthesized by Sangon
Biotech Co., Ltd. (Shanghai, China). The reaction conditions were
95°C for 30 sec, 95°C for 5 sec and 60°C for 30 sec for a total of
45 cycles. Three repeats were performed of each sample, without a
negative and RT control. The relative expression levels of the
target genes were calculated using the 2−ΔΔCq method
(12). The results were analyzed
using Image Lab software (version 3.0; Bio-Rad Laboratories,
Inc.).
Western blot analysis
The protein expression levels of PAI-1 in the
hippocampal tissues were detected using western blot analysis.
Tissues were lysed with lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) and the protein concentration was
determined using a bicinchoninic acid assay kit (RTP7102;
Real-Times Biotechnology Co., Ltd., Beijing, China). Each 20 µg
protein sample was subjected to 10% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (Wuhan Boster Biological
Technology, Ltd., Wuhan, China) and electronically transferred onto
a polyvinylidene fluoride membrane (Shanghai Yuanye Biotechnology
Co. Ltd., Shanghai, China). The membrane was blocked with 5%
fat-free milk at room temperature for 1 h and then incubated with
rabbit anti-mouse anti-PAI-1 primary antibody (1:1,000; ab66705;
Abcam, Cambridge, MA, USA), or rabbit anti-mouse anti-β-actin
antibody (1:5,000; ab129348; Abcam) at 4°C overnight. The membrane
was then incubated with goat anti-rabbit horseradish peroxidase
(HRP)-conjugated secondary antibody (1:3,000; ab6721; Abcam) at
room temperature for 1 h. The protein bands were visualized using
an enhanced chemiluminescence kit (ab65623; Abcam) and analyzed
using Image Lab software (version 3.0). β-actin was used as the
internal control.
Enzyme-linked immunosorbent assay
(ELISA)
The protein levels of PAI-1 in the blood samples
were determined using an ELISA kit (Abcam). Blood samples were
centrifuged at 1,500 rpm for 10 min to separate the serum from the
red blood cells. A 10 µl sample and 40 µl dilution solution was
added into each well on the microplate. Next, 100 µl HRP-labeled
detection antibody was added and the plate was sealed and incubated
for 1 h at 37°C, followed by washing with washing buffer included
in the kit. After washing for 5 times, 50 µl substrate A and 50 µl
substrate B was added into each well and the plate was incubated at
37°C for 15 min. Finally, 50 µl stop solution was added to stop the
reaction and the optical density was read at 450 nm using a
microplate reader (Bio-Rab Laboratories, Inc.) within 15 min. The
standard concentration curve was obtained using the standard
samples provided with the ELISA kit (ab108891; Abcam).
Bioinformatics
Bioinformatic analysis was performed using the
online target gene prediction tools, including miRanda (http://www.microrna.org/microrna/home.do), TargetScan
(www.targetscan.org), PITA (http://genie.weizmann.ac.il/pubs/mir07/mir07_data.html),
RNAhybrid (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/)
and PICTA (http://pictar.mdc-berlin.de/).
Statistical analysis
Data is expressed as the mean ± standard error. SPSS
software, version 18.0 (SPSS, Inc., Chicago, IL, USA) was used to
perform the statistical analysis. Data was subjected to the
normality test. One-way analysis of variance was performed for
multiple comparisons. The Least Significant Difference and
Student-Newman-Keuls tests were performed for equal variance, and
the Tamhane's T2 and Dunnett's T3 tests were performed when equal
variance was not assumed. P<0.05 was considered to indicate a
statistically significant difference.
Results
Learning and memory function is
reduced in model mice
To assess the learning and memory function of the
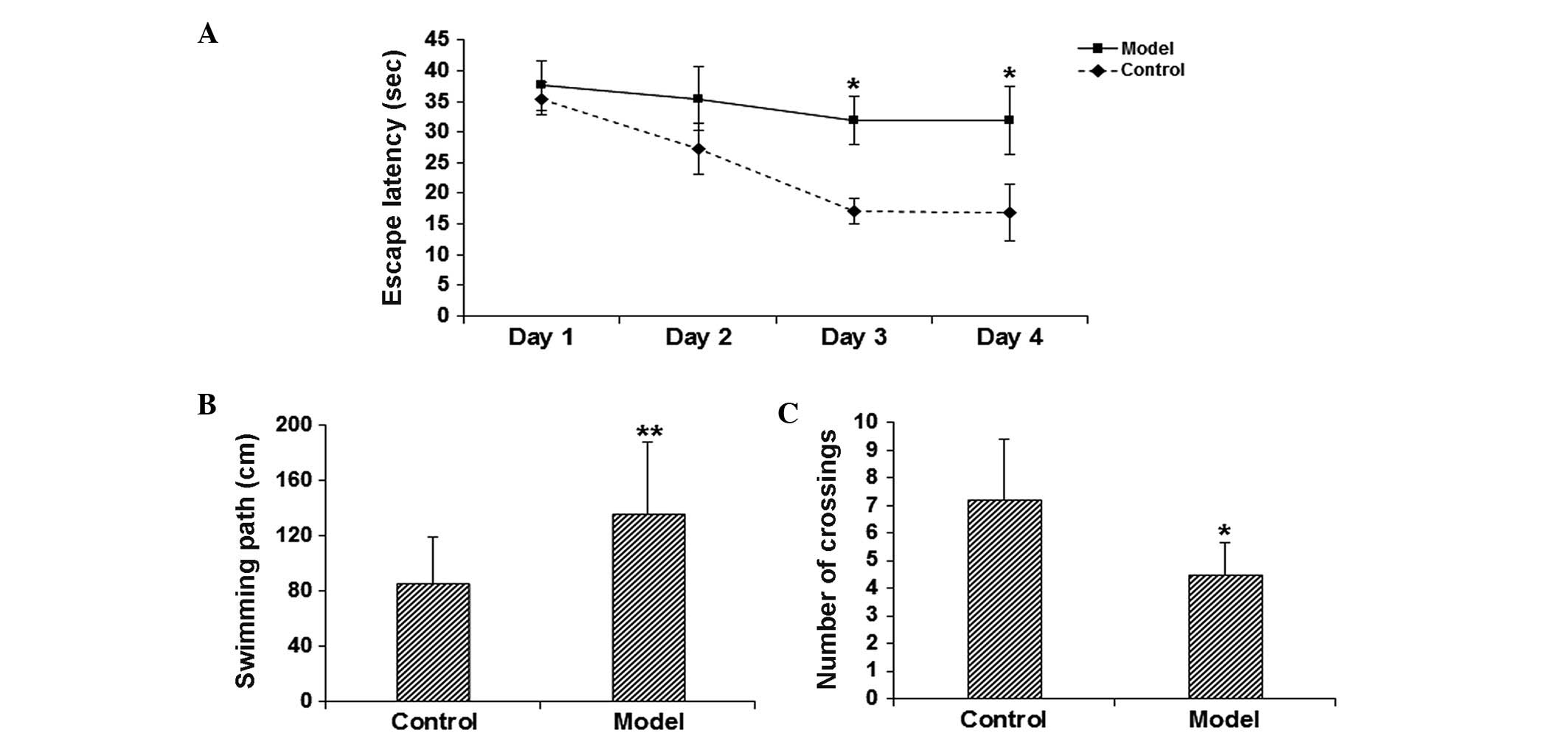
model mice, the MWM test was performed. The results demonstrated
that on days 3–4, the escape latency was significantly elevated in
the model group in comparison with the control group (P<0.05;
Fig. 1A). In addition, the probe
test on day 5 indicated that, compared with the control group, the
length of swimming path was significantly increased (P<0.05;
Fig. 1B), while the number of times
of crossing the platform location was significantly reduced
(P<0.05; Fig. 1C) in the model
group. These results suggest that the learning and memory function
is significantly impaired in the scopolamine-induced model
mice.
PAI-1 expression levels are increased
in the hippocampus and blood of model mice
To investigate the role of PAI-1 in the pathogenesis
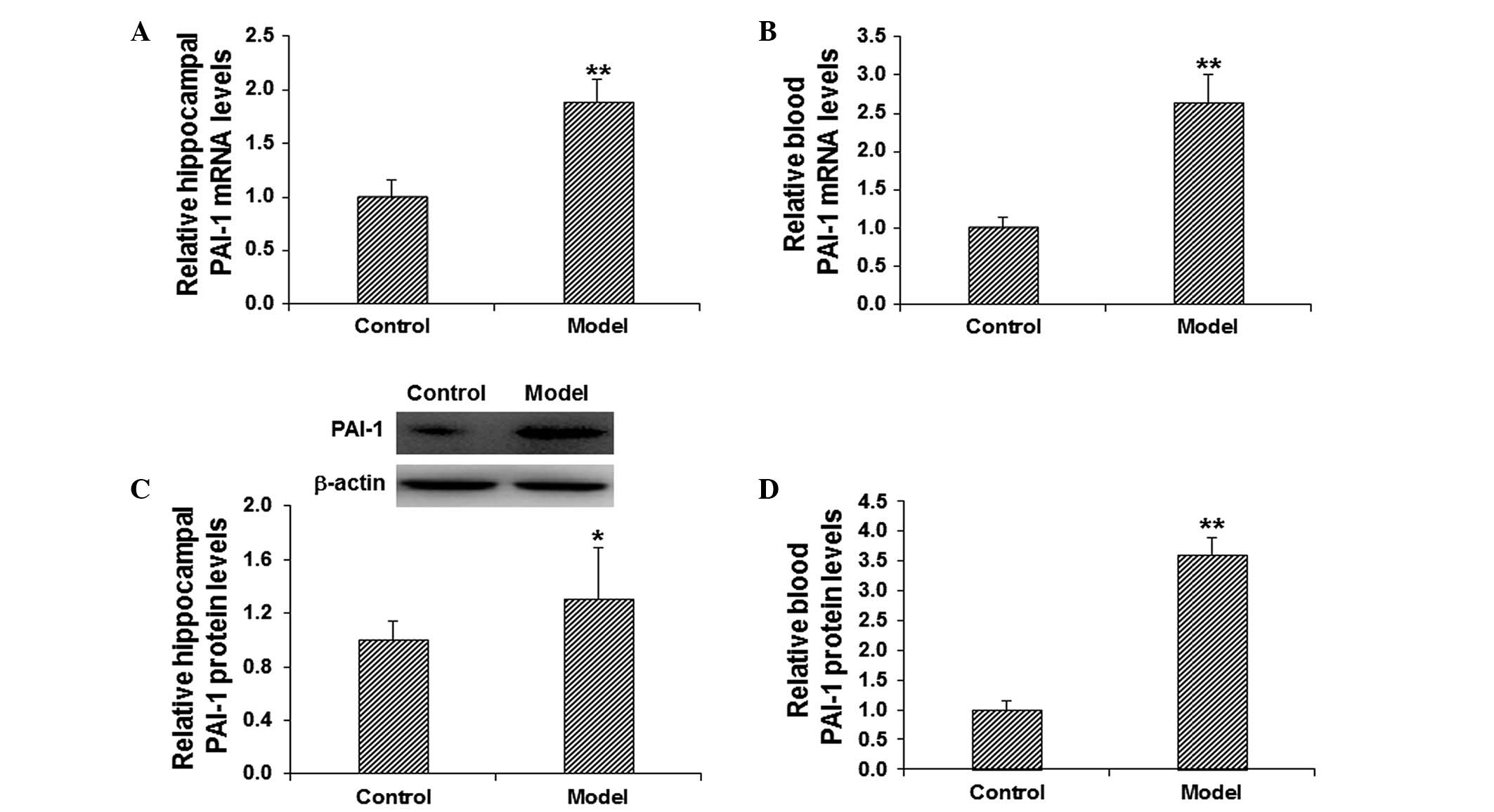
of cognitive impairment, the mRNA and protein expression levels of
PAI-1 in the hippocampus and blood were detected. The mRNA
expression level of PAI-1 was determined using RT-qPCR. The results
demonstrated that, in comparison with the control group, the mRNA
expression levels of PAI-1 in the model group were significantly
elevated in the hippocampus and blood (P<0.01; Fig. 2A and B). The protein expression
levels of PAI-1 in the hippocampus and blood were evaluated using
western blot analysis and ELISA. The results from the western blot
analysis demonstrated that the protein expression level of
hippocampal PAI-1 was significantly elevated in the model group in
comparison with the control group (P<0.05; Fig. 2C). In addition, the results from the
ELISA demonstrated that, in comparison with the control group, the
level of PAI-1 in the blood was significantly increased in the
model group (P<0.01; Fig. 2D).
These results suggest that PAI-1 may play a regulatory role in the
pathogenesis of scopolamine-induced cognitive impairment.
miR-30b levels are decreased in the
hippocampus and blood of model mice
It has been reported that PAI-1 may be regulated by
miR-30b in gastric cancer (11).
According to the bioinformatic analysis using miRanda, TargetScan,
PITA, RNAhybrid and PICTA software, it was concluded that miR-30b
may be a regulator of PAI-1 (Fig.
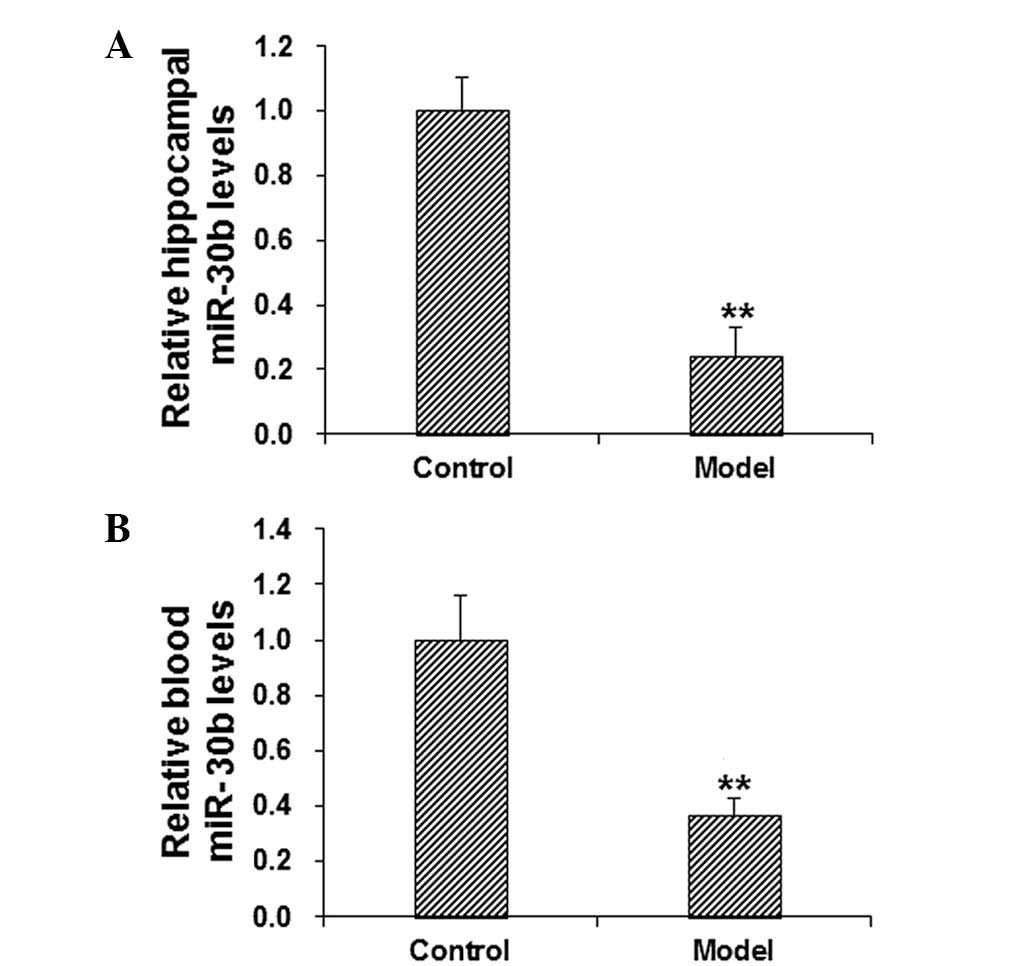
3). To further confirm the role of miR-30b in the regulation of
PAI-1 in cognitive impairment, the mRNA expression levels of
miR-30b in the hippocampus and blood in scopolamine-treated model
mice were detected using RT-qPCR. The results demonstrated that, in
comparison with the control group, the level of miR-30b in the
hippocampus and blood was significantly decreased in the model
group (P<0.01; Fig. 4). These
results suggest that miR-30b may be involved in the pathogenesis
and development of cognitive impairment.
Discussion
Cognitive impairment is a chronic central nervous
system disorder that is commonly observed in the elderly population
(13). Cognitive impairment is a
pathological state that, without treatment, may gradually develop
into different forms of dementia, such as Alzheimer's disease
(14). At present, blood tests are
primarily used to identify concomitant diseases, complications and
potential risks. Cerebrospinal fluid examination is characterized
by high sensitivity and specificity in the detection of disease;
however, due to the lack of standardized procedures for sample
collection and preparation, diagnosis based on blood tests or
cerebrospinal fluid examination alone may result in misdiagnosis
and/or missed (15). Therefore,
there is an urgent requirement for further stable genetic markers
for the diagnosis and the treatment of cognitive impairment.
The human brain is an extremely complex organ in
which the cerebral cortex forms the foundation of cognitive
function; therefore, an abnormality in the structure and function
of the cerebral cortex may result in cognitive impairment (16). Research into cognitive function
typically focuses on the hippocampus, in particular the hippocampal
neurons that serve a key role in cognitive regulation (17).
In the present study, a mouse model of cognitive
impairment was established using scopolamine (18), and the learning and memory function
of these mice was evaluated using the MWM test. Scopolamine is a
muscarinic receptor antagonist that specifically binds to, and
inhibits, muscarinic M1 receptors distributed in the hippocampus
and neocortex (19). It is
understood that scopolamine prevents the trophic and protective
effects of M1 receptor antagonists on neurons, inhibits neurite
growth, promotes β-amyloid peptide-induced cell apoptosis,
increases the phosphorylation of Tau protein (20) and causes a decline in memory
(21,22).
The results from the MWM test in the present study
demonstrated that the escape latency was significantly elongated in
model mice in comparison with control mice. In addition, the probe
test demonstrated that in model mice the navigation path was
significantly longer, and the number of times crossing the platform
region was significantly reduced. These results suggest that the
learning and memory function is markedly impaired in
scopolamine-treated mice, therefore identifying them as a suitable
model of cognitive impairment.
One of the pathological features of cognitive
impairment is the vascular amyloid deposition in cortical arteries
and arterioles, which is closely linked with PAI-1 (23). It has been demonstrated that PAI-1 is
able to regulate the formation of thrombosis, and that it is
involved in the accumulation of extracellular matrix and the
migration of smooth muscle cells (24–28).
Furthermore, PAI-1 has been observed to induce the deposition of
low density lipoprotein within the extracellular matrix of vascular
smooth muscle cells, promote the formation of fatty streaks and
atherosclerotic plaques, and thicken the vascular basement membrane
(29). These pathological changes
may result in vascular diseases and induce the occurrence and
development of atherosclerosis (24–28).
In the present study the results demonstrated that,
in comparison with the control group, the mRNA and protein
expression levels of PAI-1 were significantly elevated in the
hippocampus and blood of the model group, suggesting that PAI-1 may
serve a regulatory role in the development of cognitive impairment.
It is widely accepted that PAI-1 is closely associated with
vascular lesions, which would cause abnormal blood flow and other
pathological changes including cerebral ischemia, hypoxia,
metabolite accumulation and cognitive impairment (30,31).
Therefore, it is important to investigate the mechanisms by which
PAI-1 is regulated in disease pathogenesis.
miRs are a class of endogenous, small, non-coding
RNAs that modulate gene expression via the negative regulation of
target mRNA (32,33). miRs are important regulators under
physiological and/or pathological conditions, and have become
biomarkers for various pathological changes (34,35).
Using bioinformatic analysis in the current study, the upstream
regulatory genes of PAI-1 were predicted; miR-30b was identified as
a regulatory gene of PAI-1, and this is in accordance with a
previous study (11). Furthermore,
miR-30b has been observed to be associated with the pathogenesis of
schizophrenia (36). According to
the results in the present study, the levels of miR-30b were
significantly downregulated in scopolamine-induced cognitive
impairment, in contrast with the upregulation of the mRNA and
protein expression levels of PAI-1. These results are in accordance
with previous findings by Zhu et al (11), indicating that the downregulation of
miR-30b may be involved in the early onset and development of
cognitive impairment. Based on these findings, it can be
hypothesized that the change in expression levels of miR-30b in
blood can be used as a potential indicator for the diagnosis of
cognitive impairment, and may be applied in the clinical treatment
of the disease in the future.
In conclusion, the results from the current study
demonstrate that in scopolamine-induced mouse models of cognitive
impairment, the mRNA and protein expression levels of PAI-1 are
significantly elevated, while the miR-30b levels are significantly
declined, in the hippocampus and blood. These pathological changes
may be associated with the apoptotic processes and the vascular
lesions that can ultimately result in cognitive impairment. These
findings suggest that miR-30b is a potent regulator of PAI-1 in
cognitive impairment, and may have the potential to be used in the
diagnosis and treatment of cognitive impairment in the future.
Acknowledgements
The authors thank Director Fangyu Song from the
Department of Neurology, Laiwu Hospital Affiliated to Taishan
Medical University (Laiwu, China) for assistance with the
experimental design and performance, data collection and analysis
and manuscript preparation.
References
|
1
|
Shen X, Li Y and Xu L: Correlation between
arterial microemboli and vascular cognitive impairment in patients
with acute cerebral infarction. Zhong Guo Lao Nian Xue Za Zhi She.
33:1400–1402. 2013.(In Chinese).
|
|
2
|
Miura R and Hattori H: Overview and
assessment of cognitive function in interpreting postoperative
cognitive dysfunction. Masui. 63:1188–1195. 2014.(In Japanese).
PubMed/NCBI
|
|
3
|
Li S, Okonkwo O, Albert M and Wang MC:
Variation in Variables that Predict Progression from MCI to AD
Dementia over Duration of Follow-up. Am J Alzheimers Dis
(Columbia). 2:12–28. 2013.PubMed/NCBI
|
|
4
|
Petrella JR: Neuroimaging and the search
for a cure for Alzheimer disease. Radiology. 269:671–691. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de la Monte SM and Tong M: Brain metabolic
dysfunction at the core of Alzheimer's disease. Biochem Pharmacol.
88:548–559. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Banerjee G, Wilson D, Jäger HR and Werring
DJ: Novel imaging techniques in cerebral small vessel diseases and
vascular cognitive impairment. Biochim Biophys Acta.
10–Dec;2015.(Epub ahead of print).
|
|
7
|
Anisimova AV, Kolesnikova TI, Iutskova EV,
Galkin SS and Zimin IA: An impact of neuroprotective therapy on
blood rheological and morphodensitometric parameters in patients
with chronic cerebral ischemia. Zh Nevrol Psikhiatr Im S S
Korsakova. 114:72–80. 2014.(in Russian). PubMed/NCBI
|
|
8
|
Hua Y, Xi G, Keep RF, Wu J, Jiang Y and
Hoff JT: Plasminogen activator inhibitor-1 induction after
experimental intracerebral hemorrhage. J Cereb Blood Flow Metab.
22:55–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kohler HP and Grant PJ:
Plasminogen-activator inhibitor type 1 and coronary artery disease.
N Engl J Med. 342:1792–1801. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eitzman DT, Westrick RJ, Xu Z, Tyson J and
Ginsburg D: Plasminogen activator inhibitor-1 deficiency protects
against atherosclerosis progression in the mouse carotid artery.
Blood. 96:4212–4215. 2000.PubMed/NCBI
|
|
11
|
Zhu ED, Li N, Li BS, Li W, Zhang WJ, Mao
XH, Guo G, Zou QM and Xiao B: miR-30b, down-regulated in gastric
cancer, promotes apoptosis and suppresses tumor growth by targeting
plasminogen activator inhibitor-1. PLoS One. 9:e1060492014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-tie quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hildreth KL and Church S: Evaluation and
management of the elderly patient presenting with cognitive
complaints. Med Clin North Am. 99:311–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kapasi A and Schneider JA: Vascular
contributions to cognitive impairment, clinical Alzheimer's
disease, and dementia in older persons. Biochim Biophys Acta.
5–Jan;2016.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mishra D, Sharma S, Gupta S, Das M and
Chauhan D: Acute cysticercal meningitis – missed diagnosis. Indian
J Pediatr. 73:835–837. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Armato U, Chakravarthy B, Pacchiana R and
Whitfield JF: Alzheimer's disease: An update of the roles of
receptors, astrocytes and primary cilia. Int J Mol Med. 31:3–10.
2013.PubMed/NCBI
|
|
17
|
Christie BR and Cameron HA: Neurogenesis
in the adult hippocampus. Hippocampus. 16:199–207. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ikonomovic MD, Abrahamson EE, Isanski BA,
Wuu J, Mufson EJ and DeKosky ST: Superior frontal cortex
cholinergic axon density in mild cognitive impairment and early
Alzheimer disease. Arch Neurol. 64:1312–1317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ortega A, del Guante MA, Prado-Alcalá RA
and Alemán V: Changes in rat brain muscarinic receptors after
inhibitory avoidance learning. Life Sci. 58:799–809. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jean L, Thomas B, Tahiri-Alaoui A, Shaw M
and Vaux DJ: Heterologous amyloid seeding: Revisiting the role of
acetylcholinesterase in Alzheimer's disease. PLoS One. 2:e652–e660.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Geula C, Nagykery N, Nicholas A and Wu CK:
Cholinergic neuronal and axonal abnormalities are present early in
aging and in Alzheimer disease. J Neuropathol Exp Neurol.
67:309–318. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Minger SL, Esiri MM, McDonald B, Keene J,
Carter J, Hope T and Francis PT: Cholinergic deficits contribute to
behavioral disturbance in patients with dementia. Neurology.
55:1460–1467. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Erren M, Reinecke H, Junker R, Fobker M,
Schulte H, Schurek JO, Kropf J, Kerber S, Breithardt G, Assmann G
and Cullen P: Systemic inflammatory parameters in patients with
atherosclerosis of the coronary and peripheral arteries.
Arterioscler Thromb Vasc Biol. 19:2355–2363. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hamsten A, de Faire U, Walldius G, Dahlén
G, Szamosi A, Landou C, Blombäck M and Wiman B: Plasminogen
activator inhibitor in plasma: Risk factor for recurrent myocardial
infarction. Lancet. 2:3–9. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thögersen AM, Jansson JH, Boman K, Nilsson
TK, Weinehall L, Huhtasaari F and Hallmans G: High plasminogen
activator inhibitor and tissue plasminogen activator levels in
plasma precede a first acute myocardial infarction in both men and
women: Evidence for the fibrinolytic system as an independent
primary risk factor. Circulation. 98:2241–2247. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lupu F, Bergonzelli GE, Heim DA, Cousin E,
Genton CY, Bachmann F and Kruithof EK: Localization and production
of plasminogen activator inhibitor-1 in human healthy and
atherosclerotic arteries. Arterioscler Thromb. 13:1090–1100. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lupu F, Heim DA, Bachmann F, Hurni M,
Kakkar VV and Kruithof EK: Plasminogen activator expression in
human atherosclerotic lesions. Arterioscler Thromb Vasc Biol.
15:1444–1455. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Olexa P and Olexová M: Plasminogen
activator inhibitor-l (PAI-1), ischemic heart disease and diabetes
mellitus. Vnitr Lek. 49:222–226. 2003.(In Slovak). PubMed/NCBI
|
|
29
|
Rouch A, Vanucci-Bacqué C, Bedos-Belval F
and Baltas M: Small molecules inhibitors of plasminogen activator
inhibitor-1 - an overview. Eur J Med Chem. 92:619–636. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jokinen H, Gonçalves N, Vigário R,
Lipsanen J, Fazekas F, Schmidt R, Barkhof F, Madureira S, Verdelho
A, Inzitari D, et al: Early-stage white matter lesions detected by
multispectral MRI segmentation predict progressive cognitive
decline. Front Neurosci. 9:4552015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Matsuura Y, Yamashita A, Iwakiri T, Sugita
C, Okuyama N, Kitamura K and Asada Y: Vascular wall hypoxia
promotes arterial thrombus formation via augmentation of vascular
thrombogenicity. Thromb Haemost. 114:158–172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Inoue K: MicroRNA function in animal
development. Tanpakushitsu Kakusan Koso. 52:197–204. 2007.(In
Japanese). PubMed/NCBI
|
|
33
|
Williams AE, Moschos SA, Perry MM, Barnes
PJ and Lindsay MA: Maternally imprinted microRNAs are
differentially expressed during mouse and human lung development.
Dev Dyn. 236:572–580. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Yu Z, Li Y, Liu S, Gao C, Hou X, Yao
R and Cui L: The tumor suppressor miR-124 inhibits cell
proliferation by targeting STAT3 and functions as a prognostic
marker for postoperative NSCLC patients. Int J Oncol. 46:798–808.
2015.PubMed/NCBI
|
|
35
|
Lv ZC, Fan YS, Chen HB and Zhao DW:
Investigation of microRNA-155 as a serum diagnostic and prognostic
biomarker for colorectal cancer. Tumour Biol. 36:1619–1625. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mellios N, Galdzicka M, Ginns E, Baker SP,
Rogaev E, Xu J and Akbarian S: Gender-specific reduction of
estrogen-sensitive small RNA, miR-30b, in subjects with
schizophrenia. Schizophr Bull. 38:433–443. 2012. View Article : Google Scholar : PubMed/NCBI
|