Introduction
Osteosarcoma is the most frequently diagnosed type
of the malignant bone tumor found in adolescents and young adults
(1,2). Prognosis of patients with metastatic or
advanced osteosarcoma is ~20%, despite treatments with single or
combination of chemotherapy, radiotherapy, and surgery. Thus, novel
therapies are necessary, particularly for patients that exhibit
chemotherapy resistance (3,4).
Triptolide is a diterpene epoxide of
Tripterygium extracts, which has been suggested to possess
anti-cancer, anti-inflammatory, immunosuppressive and
anti-cystogenic activities (5).
Triptolide is effective against a number of malignancies, including
ovarian cancer, breast cancer, pancreatic cancer and neuroblastoma
(6). Triptolide supresses the
proliferation of prostate cancer cells by inhibition of expression
of SUMO-specific protease 1 (7).
Triptolide induces the apoptosis of pancreatic tumor cells by
decreasing the expression of O-GlcNac transferase to alter the
distribution of transcription factor specificity protein 1
(8). However, it is not clear if
triptolide can be used to treat osteosarcoma.
Mitogen-activated protein kinase phosphatases (MKPs)
are protein phosphatases with dual specificity (9). MKPs can dephosphorylate the
phospho-tyrosine and phospho-threonine residues on the
mitogen-activated protein kinases (MAPKs) (10). Since the MAPK family members of the
signaling molecules, such as c-Jun N-terminal kinase, p38 MAPK and
the extracellular signal-regulated kinase, serve crucial functions
in cellular signaling pathways, it may offer a potential
therapeutic strategy to control the MAPK-related pathways (11). MKP-1 is an endogenous MAPK
deactivator. MKP-1 is often overexpressed in tumors and is
considered to be related to the failure of various
chemotherapeutics (12,13).
Heat shock proteins (Hsps) are a group of proteins,
including Hsp10, 27, 40, 60, 70, 90 and 110 (14), that perform various roles in the
processes of all living organisms, from bacteria to humans. The
members of this group are functionally related proteins involved in
folding and unfolding of other proteins in the living organisms
(15,16). Under the normal growth conditions,
Hsp70s function as the ATP-molecular chaperones and facilitate
protein folding (17). Under stress
conditions, Hsp70 proteins cooperate with the increased
concentrations of unfolded and denatured proteins, avoiding toxic
aggregates via the induction of apoptosis (18,19).
Their expression is often upregulated when cells are exposed to
abnormal temperatures or extreme conditions. Changes in Hsp
expression levels are often regulated in the transcriptional steps
(20).
Hsp70 upregulation has been detected in patients
with certain types of cancers, and therefore it is speculated that
Hsp70 may contribute to resistance to chemotherapy (20). Inhibition of Hsp70 induction was
previously used as a method to benefit the anti-leukemia activity
of the Hsp90 inhibitor, 17-allylaminodemethoxy geldanamycin
(21). Ibuprofen has been found to
enhance the anti-tumor activities of cisplatin in lung cancer cells
by inhibiting Hsp70 (22). In
addition, the modulation of Hsp70 expression with quercetin
increased the chemoresponsiveness of pancreatic cancer cells to
gemcitabine (23). The aim of the
present study was to investigate whether triptolide, a diterpene
epoxide of Tripterygium extracts, can be used to treat
osteosarcoma in human cell lines.
Materials and methods
Cell lines and reagents
The human osteosarcoma cell lines (U-2 OS and MG-63)
were purchased from the American Type Culture Collection (Manassas,
VA, USA). U-2 OS cells were cultured in McCoy's 5A medium (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) at 37°C with 5%
CO2, supplemented with 10% fetal bovine serum (HyClone;
GE Healthcare Life Sciences, Logan, UT, USA), 100 U/ml penicillin
(Invitrogen; Thermo Fisher Scientific, Inc.), and 100 mg/ml
streptomycin (Invitrogen). MG-63 cells were cultured in Dulbecco's
modified Eagle's medium (Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2, supplemented with 20 mM HEPES (Thermo
Fisher Scientific, Inc.), 10% heat-inactivated fetal bovine serum,
100 U/ml penicillin (Invitrogen) and 100 mg/ml streptomycin
(Invitrogen). DNase and RNeasy isolation kits were purchased from
Qiagen (Valencia, CA, USA). Triptolide was dissolved in dimethyl
sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrozolium bromide (MTT)
assay
U-2 OS and MG-63 cells were treated with or without
triptolide (0, 5, 10, 25 or 50 nM) at 37°C for 48 h. Triptolide was
dissolved in DMSO. The cells treated with DMSO only were used as
the no drug control (0 nM). Subsequent to the experiments, cells
were incubated with 0.5 mg/ml MTT for 4 h at 37°C. The Vybrant MTT
Cell Proliferation Assay kit (V13154; Thermo Fisher Scientific,
Inc.) was used to perform the assay according to the manufacturer's
manual. The absorbance values were determined at 540 nm using a
Spectramax M2 microplate reader (Molecular Devices, Sunnyvale, CA,
USA). Viability of the treated cells relative to the control cells,
which were treated with medium only, was determined.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Extraction of the total RNAs was performed using an
RNeasy Protect Kit followed by DNase treatment according to the
manufacturer's instructions (Qiagen, Inc., Valencia, CA, USA). RNAs
were transcribed to cDNAs using the Superscript II kit according to
the manufacturer's instructions (Promega, Madison, WI, USA).
Product detection was performed by measuring fluorescence signals,
using a SYBR Green PCR kit (Takara Bio, Inc., Tokyo, Japan).
Quantification of cDNA was conducted via qPCR to a final reaction
volume of 20 µl, including 1 µl cDNA (equivalent to 50 ng input
RNA), 0.4 µM each primer and 1X SYBR, according to the
manufacturer's protocol. Thermal cycling was performed in a
LightCycler 480 Real-Time PCR system (Roche Diagnostics, Basel,
Switzerland) as follows: One cycle at 50°C for 2 min and 10 min at
95°C, followed by 40 cycles of amplification (denaturation for 15
sec at 95°C and annealing and extension for 1 min at 60°C).
Template-negative and RT-negative controls were used. Cycle
threshold values (Cq values) were calculated by using the
LightCycler software v.3.5. Expression levels of the detected
target genes were shown as the quantity relative to GAPDH, using
the 2−∆∆Cq method (24).
The analyses were performed at least three times for each sample.
Primers used in this study are listed in Table I.
 | Table I.Primers used in this study. |
Table I.
Primers used in this study.
| Primer | Sequence |
|---|
| MKP-1 | F,
5′-GTCGGTAGAGAGTCTACGCT-3′ |
|
| R,
5′-TCGTGTGGAACATTCATTC-3′ |
| Hsp70 | F,
5′-TACTATCTCCAGACTCTC-3′ |
|
| R,
5′-ATCCGCTGCTAAGCTGTG-3′ |
| GAPDH | F,
5′-GTAATGCACCCAGAGGTATG-3′ |
|
| R,
5′-CACCAATCTCATGCGGAACT-3′ |
Western blot assays
U-2 OS and MG-63 cells treated with various
conditions were harvested by centrifugation at 7,200 × g for 5 min
at 4°C, and washed twice with phosphate-buffered saline (PBS;
Beyotime Institute of Biotechnology, Beijing, China). Cell lysates
were prepared in radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology) supplemented with 0.5%
cocktail protease inhibitor (Roche Diagnostics) and 0.5 mM
phenylmethylsulfonyl fluoride. Cell lysates were subsequently
sonicated for 15 sec and centrifuged at 12,000 × g for 10 min.
Supernatants were collected and protein concentrations were
determined according to the bicinchoninic acid protocol using
bovine serum albumin as a standard, and the loading volumes of
protein samples were adjusted accordingly. Using 5X loading buffer
(250 nM Tris-Hcl, 10% SDS, 0.5% BPB, 50% glycerol and 5%
β-mercaptoethanol; pH 6.8; Proteintech; Wuhan Sanying
Biotechnology, Wuhan, China) total proteins were separated on 10%
SDS-PAGE gels (Beyotime Institute of Biotechnology), transferred
onto polyvinylidene difluoride membranes (Amresco, LLC, Solon, OH,
USA), and detected by immunoblot analyses. Membranes were blocked
with 5% skimmed milk in Tris-buffered saline with Tween 20 (TBST;
Proteintech), followed by incubation with primary antibodies
overnight at 4°C. The primary antibodies against MKP-1, Hsp70, and
β-actin were purchased from Santa Cruz Biotechnology, Inc. (Dallas,
TX, USA). The antibody information and dilution ratios were as
follows: Anti-MKP-1 (sc-370; 1:200), anti-Hsp70 (sc-32239; 1:200)
and anti-GAPDH (sc-130301; 1:5,000). Following washing three times
with TBST for 5 min, the membranes were subsequently incubated with
goat anti-mouse (sc-2005; 1:10,000) or goat anti-rabbit horseradish
peroxidase-conjugated IgG (sc-2004; 1:10,000; Santa Cruz
Biotechnology, Inc.) secondary antibodies for 1 h at room
temperature. Antibodies bound to the blots were detected using an
ECL system (32209; Pierce Biotechnology; Thermo Fisher Scientific,
Inc.). The immunoblot experiments were repeated at least 3 times.
Blot quantifications were performed using an ImageQuant LAS 500
software (GE Healthcare Life Sciences). The resulting values are
presented as the mean ± standard deviation (SD).
Statistical analysis
Experimental data are expressed as the mean ± SD.
SPSS 10.0 software (SPSS, Inc., Chicago, IL, USA) was used for
independent sample tests, followed by one-way variance analyses. In
these analyses, P<0.05 was considered to indicate statistically
significant differences.
Results
Triptolide reduces the viability of
osteosarcoma cells
To determine if triptolide was able to affect the
viability of osteosarcoma cells, the U-2 OS and MG-63 cells were
treated with triptolide (0, 5, 10, 25 or 50 nM) at 37°C for 48 h.
The cells treated with DMSO only were used as the no drug treatment
control (0 nM). Following the experiment, cells were incubated with
0.5 mg/ml MTT for 4 h at 37°C and the MTT kit was used to perform
the assay.
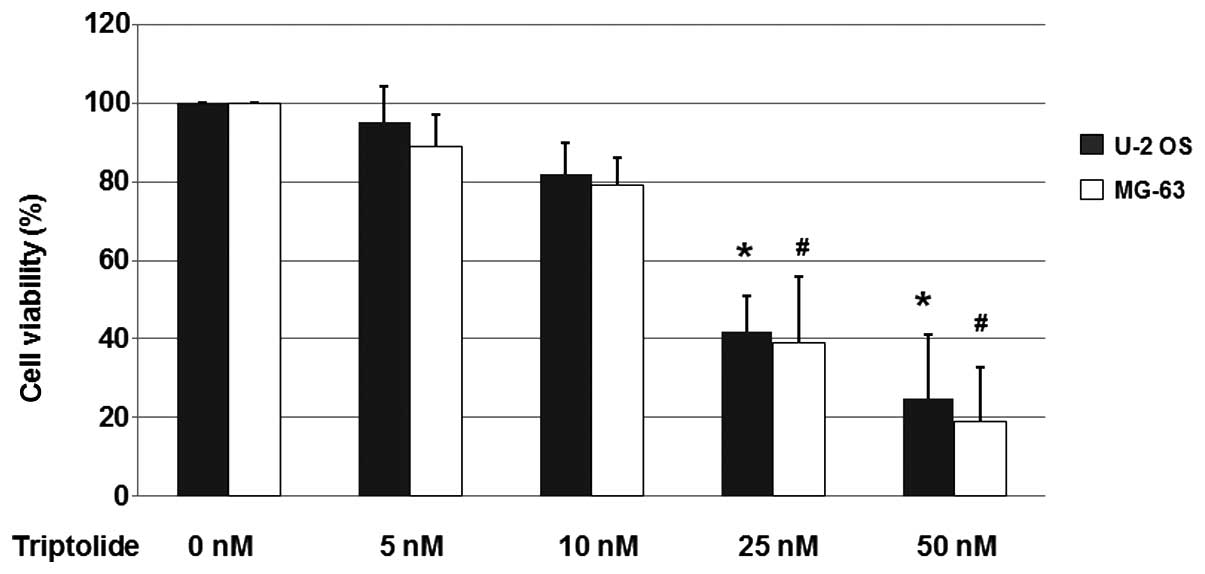
As shown in Fig. 1,
25 and 50 nM triptolide reduced cell viability significantly in the
U-2 OS and MG-63 cell populations. Treatment with 25 nM triptolide
reduced the cell viability to 42 and 39% in the U-2 OS and MG-63
cells, respectively. Furthermore, treatment with 50 nM triptolide
led to cell viabilities of 25 and 19% in the U-2 OS and MG-63
cells, respectively. These results indicated that triptolide
reduces the viability of osteosarcoma cells.
Triptolide effectively reduce MKP-1
expression
As MKP-1 is frequently overexpressed in tumor
tissues and possibly related to the chemoresistance of many
chemotherapeutics (12,13), the effects of triptolide on MKP-1
expression were determined. U-2 OS cells were treated with
triptolide (0, 5, 10, 25 and 50 nM) at 37°C for 48 h. Cells treated
with DMSO only were used as the no drug treatment control (0 nM).
Total proteins were harvested and subjected to western blot
analyses. GAPDH was used as a loading control.
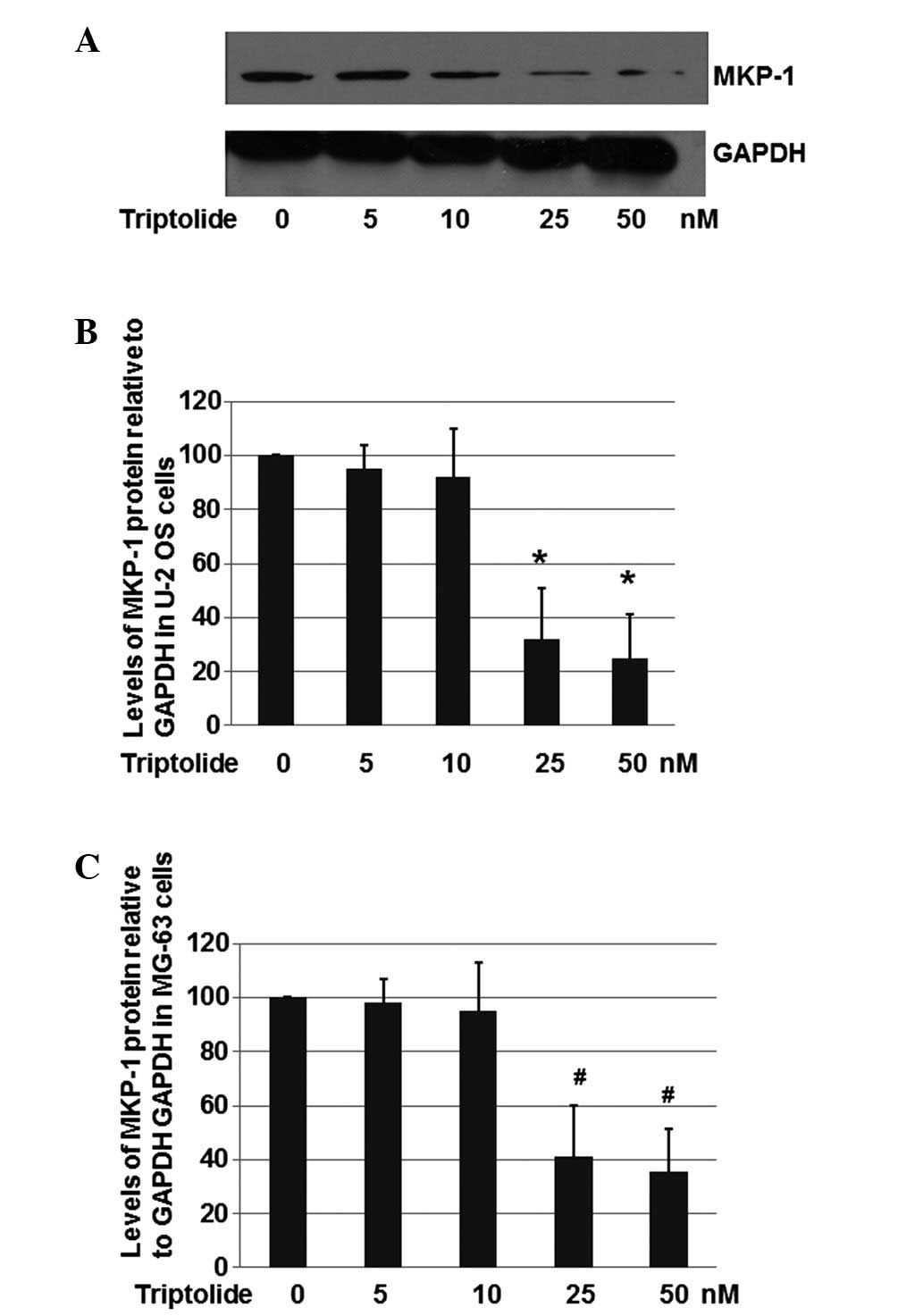
As shown in Fig. 2A and
B, triptolide with a concentration of 25 nM significantly
repressed the expression of MKP-1 to ~30.6% compared with the U-2
OS cells treated with DMSO only. In the cells treated with 50 nM
triptolide, the expression of MKP-1 was decreased to 24.2%. These
results suggest that triptolide is able to reduce MKP-1 expression
in U-2 OS cells.
The effect of triptolide on MKP-1 expression in the
human osteosarcoma cell line MG-63 was also evaluated. As shown in
Fig. 2C, triptolide at a
concentration of 25 nM inhibited the expression of MKP-1 to ~41.2%,
when compared with the level in the MG-63 cells treated with DMSO
only. In the cells treated with 50 nM triptolide, the expression of
MKP-1 was decreased to 35.6%. These results suggest that triptolide
is able to decrease MKP-1 expression in MG-63 cells. However, the
inhibitory effects on MKP-1 are slightly less than in U-2 OS
cells.
Triptolide effectively reduces Hsp70
levels
As Hsp70 is a chaperone molecule which can increase
drug resistance, the effects of triptolide on Hsp70 expression were
determined (23). U-2 OS cells were
treated with triptolide (0, 5, 10, 25 and 50 nM) at 37°C for 48 h.
Cells treated with DMSO alone were used as the no drug treatment
control (0 nM). Total proteins were harvested and subjected to
western blot analyses, with GAPDH used as a loading control.
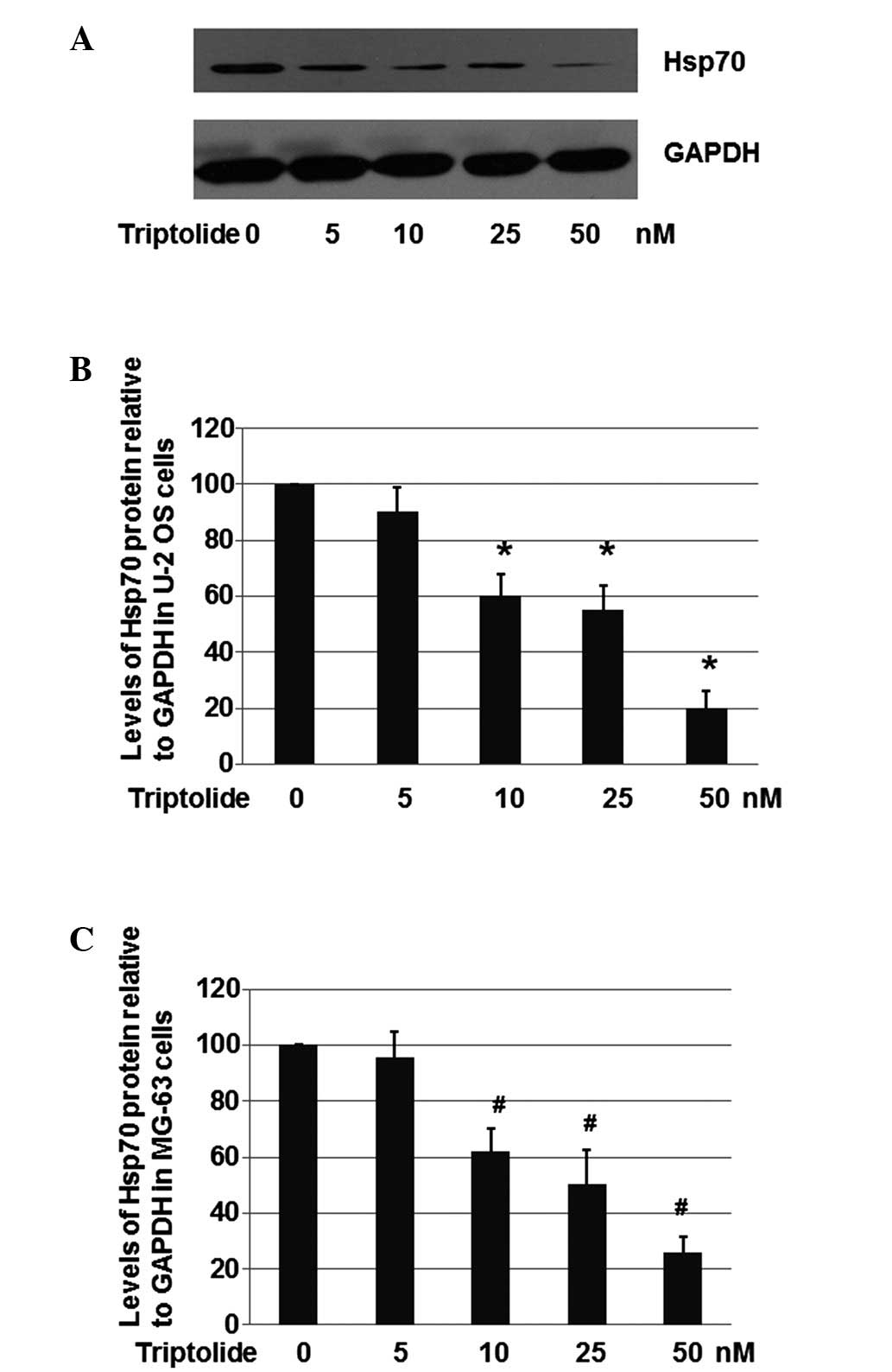
As shown in Fig. 3A and
B, triptolide at a concentration of 10, 25 or 50 nM
significantly repressed expression of Hsp70 to ~ 60.1, 54.9 and
20.6%, respectively, when compared with the level in the U-2 OS
control cells treated with DMSO only. These results suggest that
triptolide is able to decrease Hsp70 expression in U-2 OS
cells.
The present study also aimed to determine whether
triptolide decreases Hsp70 expression in MG-63 cells. As shown in
Fig. 3C, triptolide with a
concentration of 10, 25 or 50 nM significantly repressed the
expression of Hsp70 to ~62.2, 50.3 and 25.7%, respectively, when
compared with the level in the MG-63 cells treated with DMSO only.
These results suggested that triptolide is capable of decreasing
Hsp70 levels in MG-63 cells.
Triptolide effectively reduces MKP-1
mRNA expression levels in U-2 OS and MG-63 cells
In order to examine the possible effects of
triptolide on MKP-1 mRNA expression, U-2 OS cells were treated with
triptolide (0, 5, 10, 25 and 50 nM) at 37°C for 24 h. Cells treated
with DMSO only were used as the no drug treatment control (0 nM).
Total RNA was harvested and subjected to RT-qPCR analyses, with
GAPDH used as a loading control. The gene expression levels of
MKP-1 was determined using RT-qPCR detection.
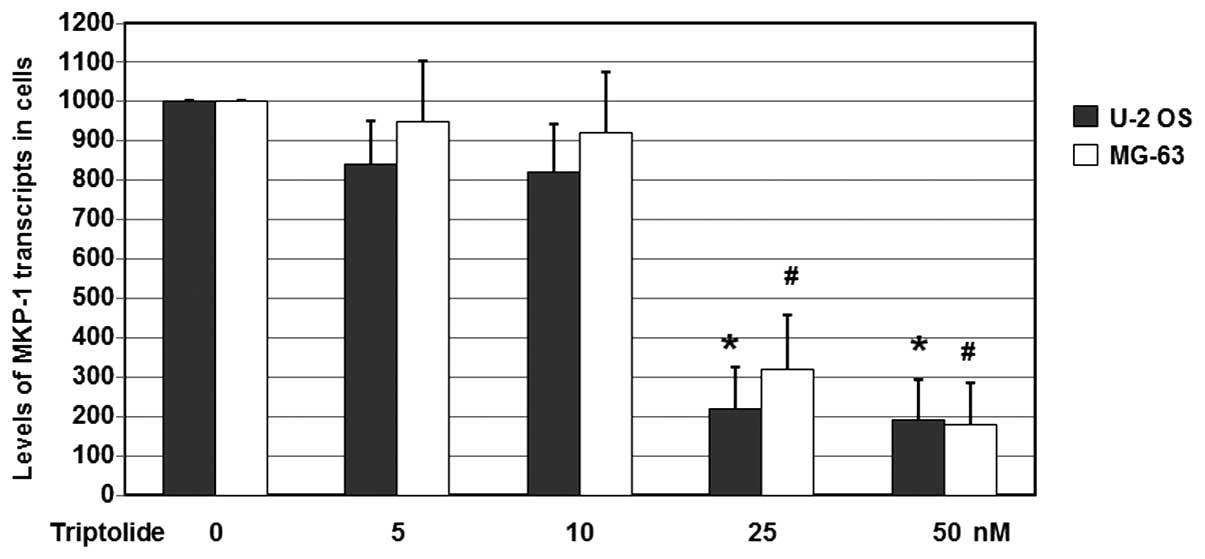
As shown in Fig. 4,
following treatment with various concentrations of triptolide, the
mRNA expression levels of MKP-1 in the U-2 OS cells were reduced
significantly. When compared with the untreated cells, the MKP-1
mRNA levels were reduced to 22.4 and 19.1% by treatment with 25 and
50 nM triptolide, respectively (P<0.05). When compared with the
untreated MG-63 cells, the MKP-1 mRNA levels were reduced to 32.2
and 18.1% (P<0.05). These results suggest that triptolide
decreases the mRNA expression levels of MKP-1 genes in U-2 OS and
MG-63 cells.
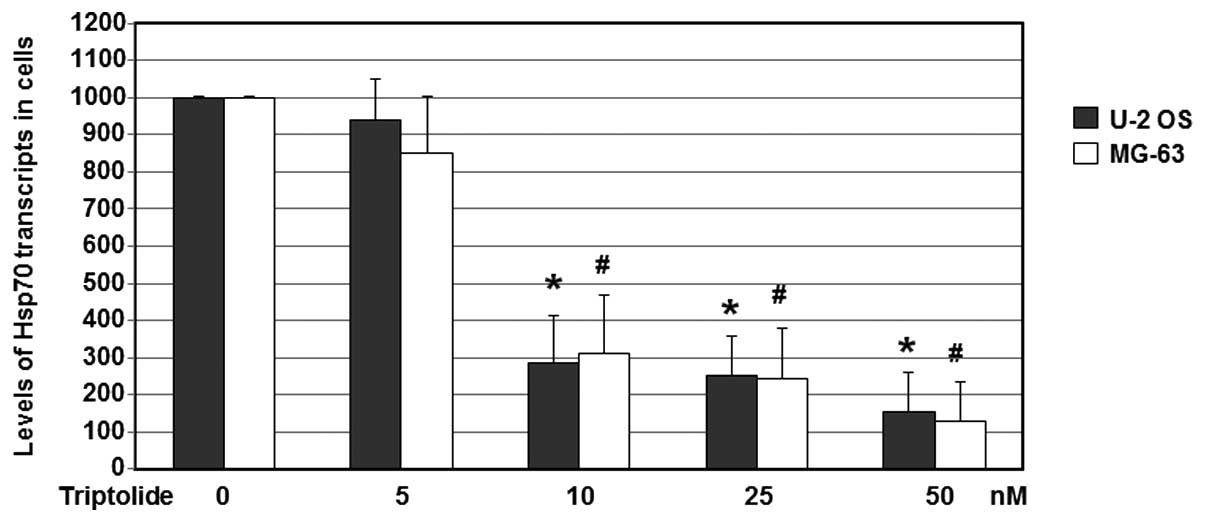
Triptolide effectively reduces Hsp70
mRNA expression levels in U-2 OS and MG-63 cells
In order to examine the effects of triptolide on
Hsp70 mRNA levels, U-2 OS and MG-63 cells were treated with
triptolide (0, 5, 10, 25 and 50 nM) at 37°C for 24 h. Cells treated
with DMSO only were used as the no drug treatment control. Total
RNA was harvested and subjected to RT-qPCR analyses, with GAPDH
used as a loading control. The gene expression of Hsp70 was
determined using RT-qPCR detection.
As shown in Fig. 5,
with treatments of various concentrations of triptolide, the mRNA
expression levels of Hsp70 in the U-2 OS and MG-63 cells were
reduced significantly. When compared with the untreated cells, the
Hsp70 mRNA levels were reduced to 15.6% following treatment with 50
nM triptolide (P<0.05). Hsp70 mRNA levels in the 50 nM
triptolide-treated MG-63 cells were reduced to 12.8% (P<0.05) of
the control expression levels. These results suggest that
triptolide is able to decrease the mRNA expression levels of Hsp70
in U-2 OS and MG-63 cells.
Discussion
Osteosarcoma is the most common type of the
malignant bone tumors found in adolescents and young adults.
Current treatments, including chemotherapy, radiotherapy and
surgery have not achieved satisfactory effects in clinical settings
(4). Triptolide has been suggested
to have anti-cancer, anti-inflammatory, immunosuppressive and
anti-cystogenic activity (5). MKP-1
protein is often overexpressed in tumors and is considered to be
related to chemoresistance (12,13).
Cancer cells express high levels of Hsps that are closely
correlated with poor prognosis (25). Under non-stress conditions, Hsp70 has
multiple functions, including protein folding and translocation of
newly synthesized proteins, and serving as a signaling molecule
(26). In colon and lung cancers,
Hsp70 expression is correlated with metastasis and poor prognosis
(27,28). Since Hsp70 is crucially involved in
multiple steps of cancer developments (29), the present study investigated the
potential of triptolide to be used as a treatment for osteosarcoma,
possibly via a mechanism associated with the regulation of MKP-1
and Hsp70.
In the present results indicated that triptolide
effectively reduced the viability of osteosarcoma cells. Treatments
of triptolide concentrations of 25 nM reduced the cell viability to
42 and 39% in the U-2 OS and MG-63 cells, respectively.
Furthermore, treatments with triptolide at a concentration of 50 nM
led to cell viabilities of 25 and 19%, respectively. These findings
suggest that a dose of triptolide as low as 25 nM is be effective
for reducing cell viability of osteosarcoma cells.
The present western blot analyses indicated that
triptolide effectively reduces MKP-1 and Hsp70 protein expression
levels. In addition, RT-qPCR assays showed that triptolide reduced
MKP-1 and Hsp70 mRNA expression in the U-2 OS and MG-63 cells.
Although the mechanisms underlying this downregulation of MKP-1 and
Hsp70 mRNA was not clear, the effects of triptolide were
confirmed.
Overexpression of Hsp70 may lead to increased tumor
growth, cancer cell migration and metastatic potential (29). Generally, increased levels of Hsp70
are frequently observed in cancer cells, where Hsp70 improves
resistance to stress-induced apoptosis, contributing to suppression
of senescence, and is also associated with metastatic development
and drug resistance (30–32). Triptolide is known to induce
apoptosis in gastric cancer cells via the inhibition of murine
double minute 2 overexpression (33). In animal model experiments,
triptolide reduced neuropathology in a mouse model of Alzheimer's
disease by upregulating the level of an insulin-degrading enzyme, a
major Aβ-degrading enzyme in the brain (34). The results of the present study
improve our understanding of triptolide as a potential therapy for
cancer.
The results of the present study indicated that
triptolide treatment significantly decreases the viability of
osteosarcoma cells. This effect may be associated with the
decreased expression levels of MKP-1 and Hsp70 following treatment
with triptolide. Future studies investigating the manipulation of
the molecular structure of triptolide are required in order to
obtain a series of derivatives and to evaluate the effects on
OS.
Acknowledgements
This study was supported by project of Medical
Technology Research Center, Ministry of Health (grant no.
W2014ZT119).
References
|
1
|
Kourti M, Sidi V and Papakonstantinou E:
Osteosarcoma in an adolescent previously treated for Hodgkin's
Disease. Hippokratia. 18:962014.PubMed/NCBI
|
|
2
|
Xu SF, Yu XC, Xu M and Chen X: Successful
management of a childhood osteosarcoma with epiphysiolysis and
distraction osteogenesis. Curr Oncol. 21:e658–e662. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou Y, Huang Z, Wu S, Zang X, Liu M and
Shi J: miR-33a is up-regulated in chemoresistant osteosarcoma and
promotes osteosarcoma cell resistance to cisplatin by
down-regulating TWIST. J Exp Clin Cancer Res. 33:122014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang J and Zhang W: New molecular insights
into osteosarcoma targeted therapy. Curr Opin Oncol. 25:398–406.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wong KF, Yuan Y and Luk JM:
Tripterygium wilfordii bioactive compounds as anticancer and
anti-inflammatory agents. Clin Exp Pharmacol Physiol. 39:311–320.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou ZL, Yang YX, Ding J, Li YC and Miao
ZH: Triptolide: Structural modifications, structure-activity
relationships, bioactivities, clinical development and mechanisms.
Nat Prod Rep. 29:457–475. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang W, He T, Chai C, Yang Y, Zheng Y,
Zhou P, Qiao X, Zhang B, Liu Z and Wang J: Triptolide inhibits the
proliferation of prostate cancer cells and down-regulates
SUMO-specific protease 1 expression. PLoS One. 7:e376932012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Banerjee S, Sangwan V, McGinn O, Chugh R,
Dudeja V, Vickers SM and Saluja AK: Triptolide-induced cell death
in pancreatic cancer is mediated by O-GlcNAc modification of
transcription factor Sp1. J Biol Chem. 288:33927–33938. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Crowell S, Wancket LM, Shakibi Y, Xu P,
Xue J, Samavati L, Nelin LD and Liu Y: Post-translational
regulation of mitogen-activated protein kinase phosphatase (MKP)-1
and MKP-2 in macrophages following lipopolysaccharide stimulation:
The role of the C termini of the phosphatases in determining their
stability. J Biol Chem. 289:28753–28764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Doddareddy MR, Rawling T and Ammit AJ:
Targeting mitogen-activated protein kinase phosphatase-1 (MKP-1):
structure-based design of MKP-1 inhibitors and upregulators. Curr
Med Chem. 19:163–173. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen FM, Chang HW, Yang SF, Huang YF, Nien
PY, Yeh YT and Hou MF: The mitogen-activated protein kinase
phosphatase-1 (MKP-1) gene is a potential methylation biomarker for
malignancy of breast cancer. Exp Mol Med. 44:356–362. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lei YY, Wang WJ, Mei JH and Wang CL:
Mitogen-activated protein kinase signal transduction in solid
tumors. Asian Pac J Cancer Prev. 15:8539–8548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Park J, Lee J, Kang W, Chang S, Shin EC
and Choi C: TGF-β1 and hypoxia-dependent expression of MKP-1 leads
tumor resistance to death receptor-mediated cell death. Cell Death
Dis. 4:e5212013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jolly C and Morimoto RI: Role of the heat
shock response and molecular chaperones in oncogenesis and cell
death. J Natl Cancer Inst. 92:1564–1572. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Borges JC, Seraphim TV, Mokry DZ, Almeida
FC, Cyr DM and Ramos CH: Identification of regions involved in
substrate binding and dimer stabilization within the central
domains of yeast Hsp40 Sis1. PLoS One. 7:e509272012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dougan DA, Mogk A and Bukau B: Protein
folding and degradation in bacteria: to degrade or not to degrade?
That is the question. Cell Mol Life Sci. 59:1607–1616. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kelley WL: Molecular chaperones: How J
domains turn on Hsp70s. Curr Biol. 9:R305–308. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nylandsted J, Brand K and Jäättelä M: Heat
shock protein 70 is required for the survival of cancer cells. Ann
N Y Acad Sci. 926:122–125. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi W, Zhou Y, Wild J, Adler J and Gross
CA: DnaK, DnaJ and GrpE are required for flagellum synthesis in
Escherichia coli. J Bacteriol. 174:6256–6263. 1992.PubMed/NCBI
|
|
20
|
Rérole AL, Jego G and Garrido C: Hsp70:
Anti-apoptotic and tumorigenic protein. Methods Mol Biol.
787:205–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo F, Rocha K, Bali P, Pranpat M, Fiskus
W, Boyapalle S, Kumaraswamy S, Balasis M, Greedy B and Armitage ES:
Abrogation of heat shock protein 70 induction as a strategy to
increase antileukemia activity of heat shock protein 90 inhibitor
17-allylamino-demethoxy geldanamycin. Cancer Res. 65:10536–10544.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hyun JJ, Lee HS, Keum B, Seo YS, Jeen YT,
Chun HJ, Um SH and Kim CD: Expression of heat shock protein 70
modulates the chemoresponsiveness of pancreatic cancer. Gut Liver.
7:739–746. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Endo H, Yano M, Okumura Y and Kido H:
Ibuprofen enhances the anticancer activity of cisplatin in lung
cancer cells by inhibiting the heat shock protein 70. Cell Death
Dis. 5:e10272014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kelley WL: The J-domain family and the
recruitment of chaperone power. Trends Biochem Sci. 23:222–227.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Calderwood SK, Khaleque MA, Sawyer DB and
Ciocca DR: Heat shock proteins in cancer: Chaperones of
tumorigenesis. Trends Biochem Sci. 31:164–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Liang X, Wu S, Murrell GA and Doe
WF: Inhibition of colon cancer metastasis by a 3′-end antisense
urokinase receptor mRNA in a nude mouse model. Int J Cancer.
92:257–262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhuang H, Jiang W, Zhang X, Qiu F, Gan Z,
Cheng W, Zhang J, Guan S, Tang B and Huang Q: Suppression of HSP70
expression sensitizes NSCLC cell lines to TRAIL-induced apoptosis
by upregulating DR4 and DR5 and downregulating c-FLIP-L
expressions. J Mol Med (Berl). 91:219–235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Juhasz K, Lipp AM, Nimmervoll B,
Sonnleitner A, Hesse J, Haselgruebler T and Balogi Z: The complex
function of hsp70 in metastatic cancer. Cancers (Basel). 6:42–66.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gabai VL, Yaglom JA, Waldman T and Sherman
MY: Heat shock protein Hsp72 controls oncogene-induced senescence
pathways in cancer cells. Mol Cell Biol. 29:559–569. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rohde M, Daugaard M, Jensen MH, Helin K,
Nylandsted J and Jäättelä M: Members of the heat-shock protein 70
family promote cancer cell growth by distinct mechanisms. Genes
Dev. 19:570–582. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gurbuxani S, Schmitt E, Cande C,
Parcellier A, Hammann A, Daugas E, Kouranti I, Spahr C, Pance A,
Kroemer G and Garrido C: Heat shock protein 70 binding inhibits the
nuclear import of apoptosis-inducing factor. Oncogene.
22:6669–6678. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang BY, Cao J, Chen JW and Liu QY:
Triptolide induces apoptosis of gastric cancer cells via inhibiting
the overexpression of MDM2. Med Oncol. 31:2702014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng S, LeBlanc KJ and Li L: Triptolide
preserves cognitive function and reduces neuropathology in a mouse
model of Alzheimer's disease. PLoS One. 9:e1088452014. View Article : Google Scholar : PubMed/NCBI
|