Introduction
Systematic juvenile idiopathic arthritis (SJIA) is a
type of chronic arthritis that is characterized by systematic
symptoms (high fever, rash, and enlargement of the liver, spleen
and lymph nodes) and various increased inflammatory factors, such
as C reactive proteins (CRP) and ferritin (FER) (1). SJIA is the most severe type of juvenile
idiopathic arthritis (JIA), and can induce macrophage activation
syndrome that results in mortality (2). Different from adult rheumatoid
arthritis (RA), which presents as significant joint deformity,
patients with JIA (specifically patients with SJIA) are diagnosed
according to common symptoms and high levels inflammatory factors
(3). However, there is a current
requirement for more specific markers for the condition.
MicroRNA (miRNA) is a type of non-coding single
stranded RNA that is widespread in the body and has a regulatory
function (4). It binds specifically
to the 3′-untranslated region of target mRNA in order to degrade or
inhibit its translation, thereby participating in many basic
bioprocesses, such as cell development, proliferation and
differentiation, and the cell cycle (4). Therefore, miRNA has an important role
in numerous diseases, including cancer, metabolic diseases, nerve
disorders and infectious and autoimmune diseases (2). Recently, a number of studies suggested
that miRNA serves an important role in regulating chronic
inflammation, in which some miRNAs, such as miR-155 (5) and miR-146a (6), participated in the proliferation and
differentiation of immune cells, such as T and B cells, and
regulated their abnormal expression in autoimmune diseases.
Increased expression levels of miR-146a partially contributes
towards upregulated expression of TNF-α; however, the mechanism
underlying this increase in expression and the site of action of
miR-146a in signaling pathways remains unclear (7). Meanwhile, it could not explain JIA with
normal TNF-α and bad efficacy of TNF-α antagonist to systematic
arthritis, thus it was assumed that there was an interaction of a
different signaling pathway in JIA. Despite recent random control
studies suggesting that there are various expression levels of
numerous types of miRNA, including miR-155, miR-19a, miR-203,
miR-21 and miR-124a, in synovioblasts, PBMC and T cells from
patients with RA (7–14), the studies did not investigate the
target genes of miRNAs, which could demonstrate the immunological
dysregulation, inflammation and differentiation of cells.
Therefore, in the present study, miR-19a, miR-21 and their
associated target genes involved in SJIA were used to investigate
the roles of miRNA in SJIA.
Materials and methods
Inclusion criteria
A total of 20 patients with active SJIA (fever for
>2 weeks; high fever type with or without articular pain; liver,
spleen and lymph node enlargement; increased expression levels of
FER; CRP ≥10 mg/l, or erythrocyte sedimentation rate ≥28 ram/1 h)
were diagnosed in Guangzhou Women and Children's Medical Center
(Guangdong, China) from January to March 2015, and 20 normal
control patients were recruited from the Health Care Section of the
hospital. Patients were diagnosed and classified according to the
Task-force of the Pediatric Standing Committee of International
League of Associations for Rheumatology discussion draft (15). Patients with recurrent bacterial,
mycoplasma, viral, fungal and mycobacterial infections, an
infection history, an active infection, or blood diseases detected
by bone marrow cytology were excluded from the study. Baseline
characteristics of the control subjects and patients with SJIA are
presented in Table I. The study
protocol was approved by the ethics committee of Guangzhou Women
and Children's Medical Center, and written-informed consent was
obtained from all subjects.
 | Table I.Baseline data of the patients with
systematic juvenile idiopathic arthritis and healthy controls. |
Table I.
Baseline data of the patients with
systematic juvenile idiopathic arthritis and healthy controls.
| Clinical
information | SJIA group
(n=20) | Control group
(n=20) |
|---|
| Age (years) | 7.36±3.64 | 8.90±3.69 |
| Male proportion
(%) | 65 | 70 |
| Weight (kg) | 13.45±6.34 | 12.67±7.83 |
| DAS28 scores | 6.23±2.34 | – |
| Abnormal BMD or
synovitis suggested by MRI | 16 (80%) | – |
| Anti-cytokeratin
antibody | 1 (12.5%) | – |
| Anti-cyclic
citrullinated peptide antibody (U/ml x±s) | 9.77±8.07 | – |
| RF-IgG (U/ml
x±s) | 13.60±9.99 | – |
| Ferritin (ng/ml,
x±s) | 1079.47±900.87 | – |
| CRP (mg/l, x±s) | 98.61±63.80 | – |
| ESR (mm/h, x±s) | 53.12±26.59 | – |
| AST median | 56.23 (6,74) | – |
| ALT median | 47.69 (11,16) | 24 (5,20) |
Reagents
Ficoll Paque PLUS solution and phosphate-buffered
saline (PBS) were purchased from GE Healthcare Life Sciences
(Logan, UT, USA); the RNA extraction kit (9112; Takara
Biotechnology Co., Ltd. (Dailan, China), reverse transcription kit,
RNAiso (9753A), miR-X iRNA First Strand Synthesis kit (638315) and
SYBR® Premix Ex Taq (Tli RNaseH Plus; RR420A)
kits were purchased from Takara Biotechnology Co., Ltd.. The
primers of miR-19a, miR-21 and U6 were synthesized by Takara
Biotechnology Co., Ltd. The primer sequences were as follows: STAT3
forward, 5′-GCCAGAGAGCCAGGAGCA-3′, and reverse,
5′-ACACAGATAAACTTGGTCTTCAGGTATG-3′; SOCS3 forward,
5′-CAGCTCCAAGAGCGAGTACC-3′, and reverse,
5′-TGACGCTGAGCGTGAAGAAG-3′; TNF-α forward,
5′-ACCCTCACACTCAGATCATC-3′, and reverse,
5′-GAGTAGACAAGGTACAACCC-3′; IL-6 forward,
5′-AGCCAGAGTCCTTCAGAGAG-3′, and reverse,
5′-GATGGTCTTGGTCCTTAGCC-3′; β-actin forward,
5′-GAGCTACGAGCTGCCTGACG-3′, and reverse,
5′-GTAGTTTCGTGGATGCCACAG-3′. The primers were synthesized by
Shenzen Huada Gene Technology Co., Ltd. (Shenzen, China)
company.
Separation of PBMCs and extraction of
total RNA
Fasting blood was drawn from the patients with SJIA
and the control subjects, and placed in 4 ml
ethylenediaminetetraacetic acid. Subsequently, 4 ml of PBS was
added in order to dilute the sample. The same volume of Ficoll
Paque PLUS solution and diluted blood was added into 15 ml
centrifuge tubes and centrifuged at 160 × g for 20 min at 18–21°C.
The cell pellet was then collected and added into pentaploid PBS,
and centrifuged twice at 160 × g for 20 min. Waste liquid was
discarded, and 1 ml RNAiso and 200–300 Ml chloroform
(Sigma-Aldrich, St. Louis, MO, USA) was added. The samples were
incubated for 5 min at 25–28°C and centrifuged at 12,000 × g for 15
min at 4°C. Approximately 400 µl supernatant was obtained, to which
1 ml cold isopropanol (Sigma-Aldrich) was added, mixed reversely
and incubated at 4°C for 10 min, prior to being centrifuged at
12,000 × g for 10 min at 4°C. The supernatant was then discarded,
and 10 ml 75% ethanol prepared with diethylpyrocarbonate (DEPC)
(Sigma-Aldrich) water was added (ethanol:DEPC; 3:1), followed by
centrifugation at 7,500 × g for 5 min at 4°C. Finally, 20 µl DEPC
water was added to the samples, and the absorbance was measured
using a spectrophotometer (Thermo NanoDrop 2000; Thermo Fisher
Scientific, Inc., Waltham, MA, USA).
Reverse transcription-quantitative PCR
(RT-qPCR)
A PrimeScript II 1st Strand cDNA Synthesis Kit
(Takara Biotechnology Co., Ltd.) was used. RNAiso was used to
extract total RNA. A total of 3.8 µl RNA was added into 0.2 ml PCR
tubes, followed by the addition of 1.2 µl mRQ enzyme mix and 5 µl
buffer. The samples were then incubated at 37°C for 1 h, and 95°C
for 5 min. Following incubation, 90 µl ddH2O was added and the
samples were stored at −20°C for the subsequent experiments. For
qPCR, 9 µl dddH20, 12.5 µl SYBR Green, 0.5 µl upstream primers, 0.5
µl downstream primers, ROC 50×0.5 µl and 2 µl cDNA were used, and
the samples were incubated at 95°C for 10 sec, 95°C for 5 sec, 60°C
for 20 sec, then 40 cycles at 72°C for 5 min using a Roche
LightCycler 480II (Roche Diagnostics, Basel, Switzerland). Each
gene had three repeats. U6 was used to normalize the expression of
miRNA, and β-actin was used to normalize the expression of STAT3,
SOCS3, IL-6 and TNF-α. The ΔCq and ΔΔCq values were calculated, and
the relative expression levels of mRNA, associated target genes and
U6 were presented as the 2−ΔΔCq value (16).
Statistical analysis
Data were analyzed using SPSS version 13.0 software
(SPSS, Inc., Chicago, IL, USA), and miR-19a and miR-21 were
expressed as relative expression levels. A Kruskal-Wallis test was
used for intergroup comparisons, and a Pearson's correlation
coefficient was used to calculate the correlation between miR-19a,
miR-21 and inflammatory factors. P<0.05 was considered to
indicate a statistically significant difference.
Results
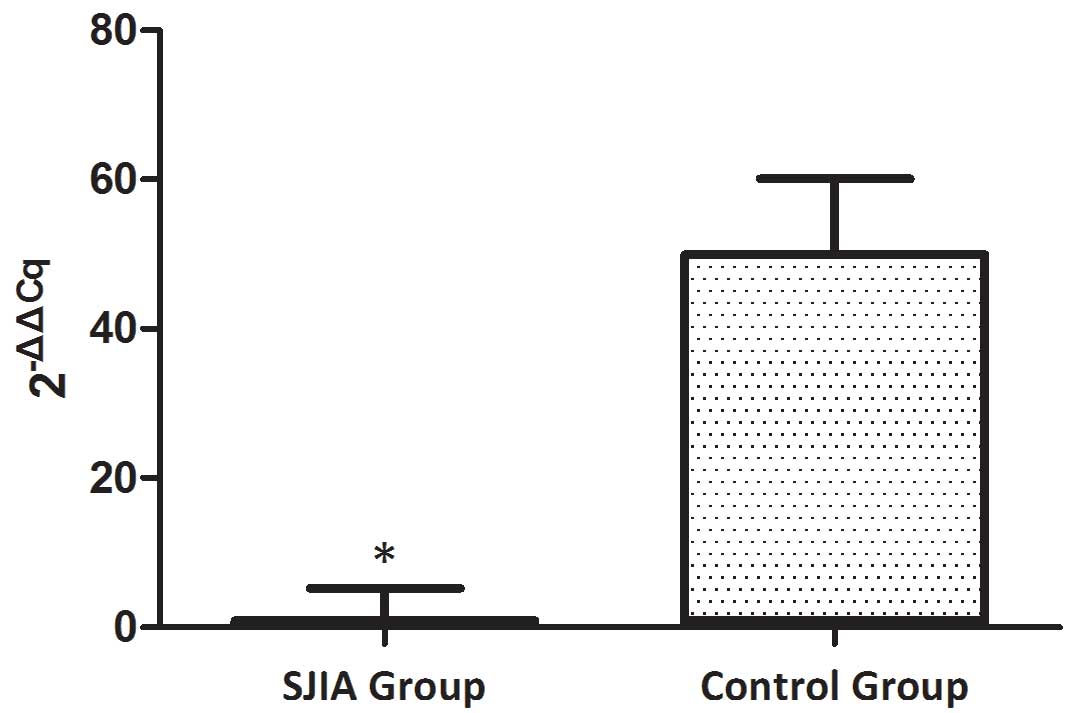
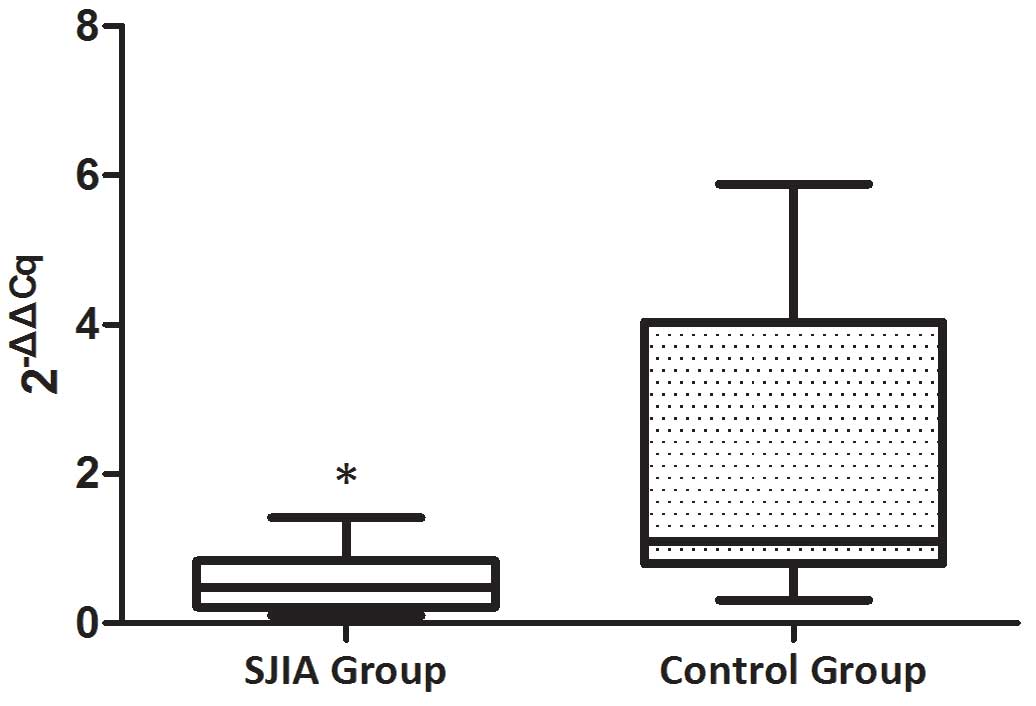
The expression levels of miR-21 and
miR-19a are decreased in patients with SJIA compared with the
controls
The expression levels of miR-21 and miR-19a were
investigated in PBMCs from active state patients with SJIA and
control subjects. The expression levels of miR-21 were lower in
patients with SJIA compared with the control group (Fig. 1), and the relative expression levels
of miR-21 in the control group were 7.7-fold higher compared with
those of the patients with SJIA (Table
II; P=0.036). In addition, the expression levels of miR-19a
were lower in patients with SJIA compared with the control group
(Fig. 2), and the relative
expression levels of miR-19a in the control group were 11.3-fold
higher compared with those of the patients with SJIA (Table III; P=0.014).
| Table II.Expression levels of miR-21 in PBMCs
of patients with SJIA and the control group
(2−ΔΔCq). |
Table II.
Expression levels of miR-21 in PBMCs
of patients with SJIA and the control group
(2−ΔΔCq).
| Group | miR-21 Cq | U6 Cq | ΔCq | ΔΔCq |
2−ΔΔCq |
|---|
| SJIA
groupa | 25.2±1.04 | 18.7±1.07 | 6.4±0.3 | 0.0±0.3 | 1.0
(0.8–1.2)b |
| Control
groupa | 24.6±0.94 | 21.1±1.13 | 3.5±0.4 | −2.9±0.4 | 7.7
(7–8.5)b |
| Table III.Expression levels of miR-19a in PBMCs
of patients with SJIA and the control group
(2−ΔΔCq). |
Table III.
Expression levels of miR-19a in PBMCs
of patients with SJIA and the control group
(2−ΔΔCq).
| Group | miR-19a Cq | U6 Cq | ΔCq | ΔΔCq |
2−ΔΔCq |
|---|
| SJIA
groupa | 28.0±0.71 | 15.7±1.78 | 12.3±1.1 | 0.0±1.1 | 1.0
(0.9–1.3)b |
| Control
groupa | 29.9±0.69 | 21.1±1.08 | 8.8±0.91 | −3.5±0.91 | 11.3
(10–12.1)b |
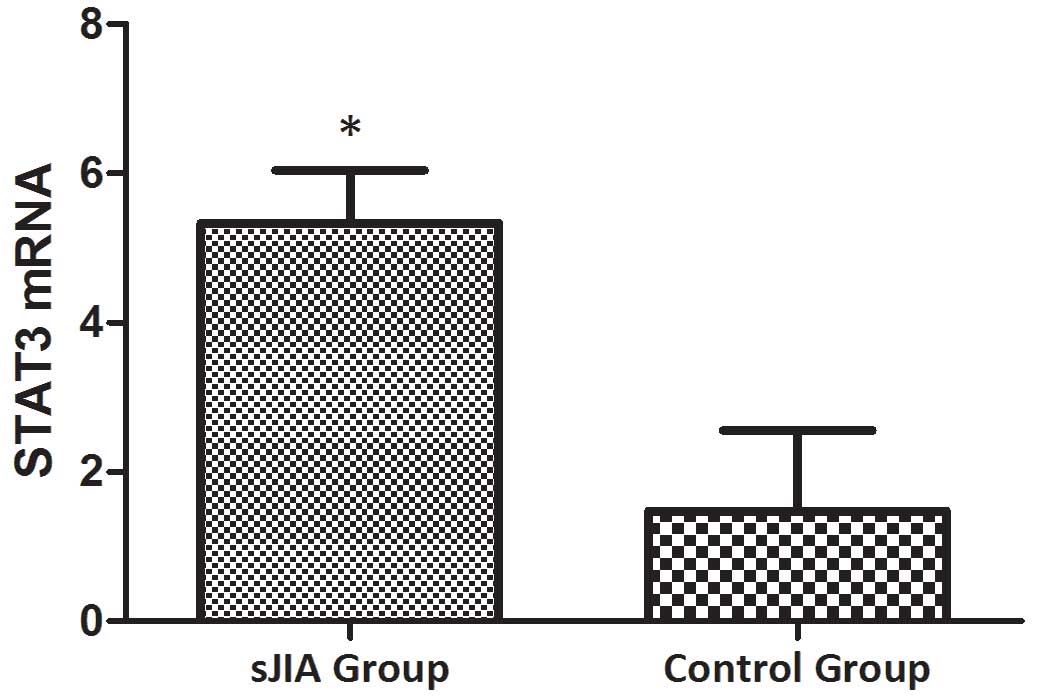
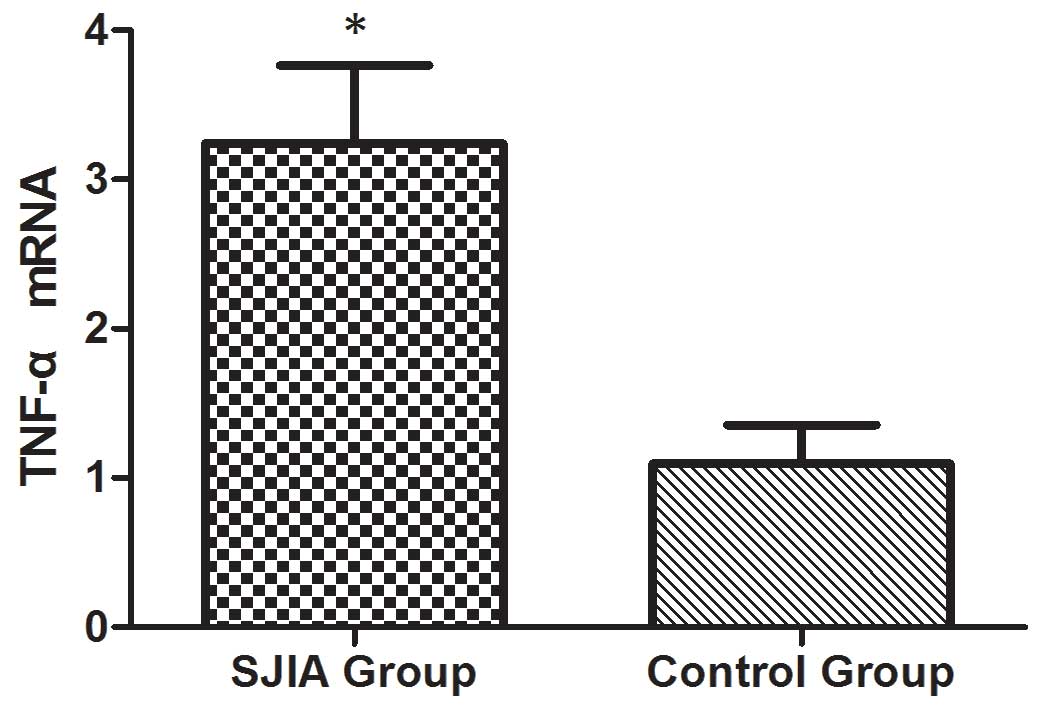
The mRNA expression levels of STAT3,
TNF-α, IL-6 and SOCS3 are higher in patients with SJIA compared
with the controls
The mRNA expression levels of STAT3, TNF-α, IL-6 and
SOCS3 were expressed in the peripheral blood of patients with SJIA.
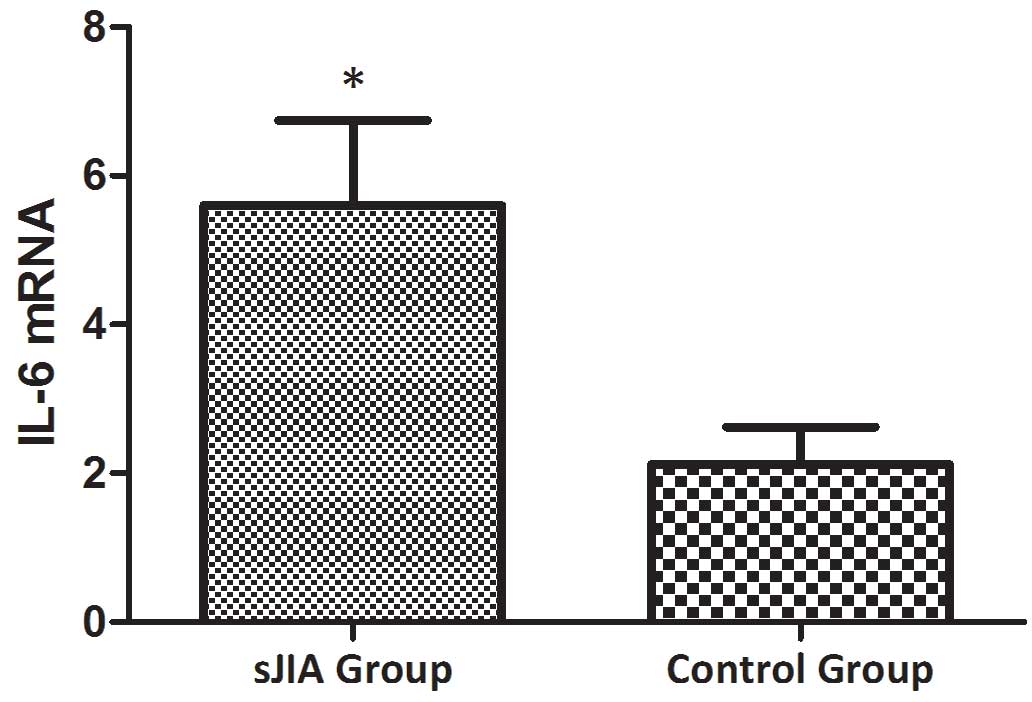
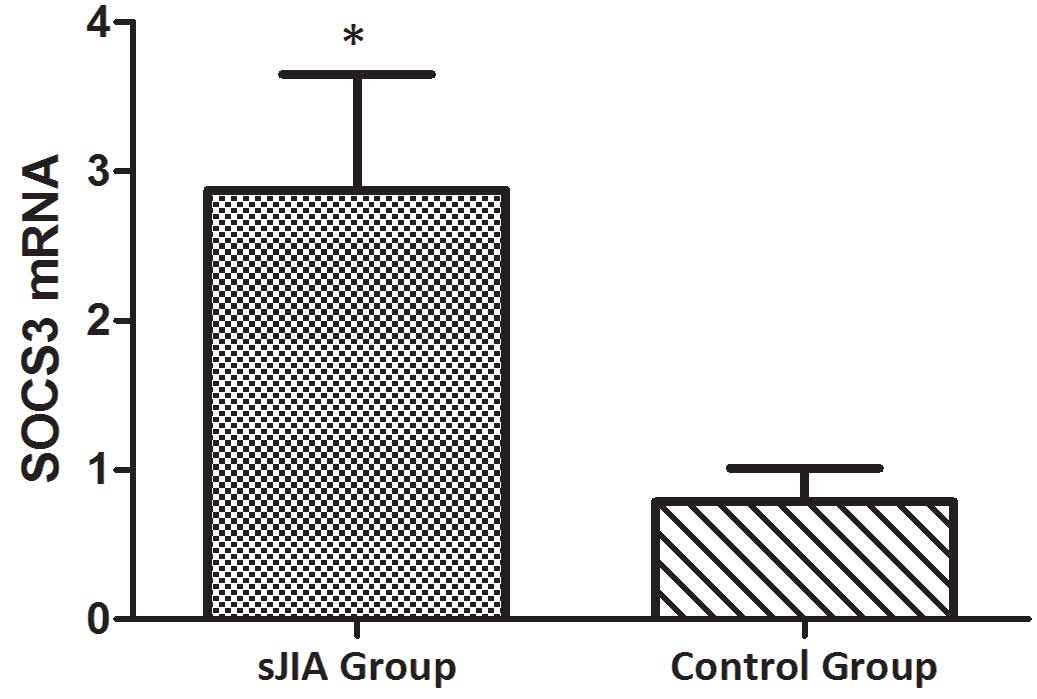
The relative expression levels of STAT3 (Fig. 3), TNF-α (Fig. 4), IL-6 (Fig. 5) and SOCS3 (Fig. 6) mRNA were higher in patients with
SJIA compared with the control group. In addition, the expression
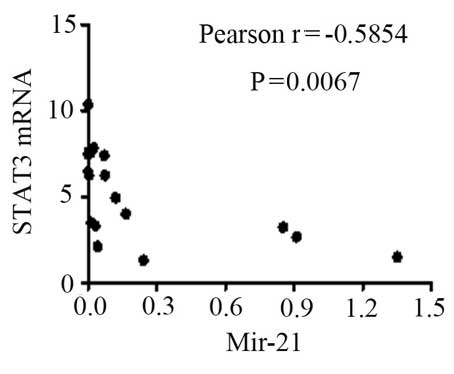
levels of STAT3 mRNA were negatively correlated with miR-21
2−ΔΔCq (Fig. 7;
r=−0.5854; P=0.0067); this suggests that miR-21 may participate in
the regulation of the expression of STAT3. TNF-α is the target gene
of miR-21 (17); therefore, the
expression of miR-21 may be related to TNF-α. However, no
correlation was observed in the results of the present study
(r=2.138; P=0.43). STAT3 can improve the expression of TNF-α mRNA,
and this may restrict the negative regulation of miR-21. The
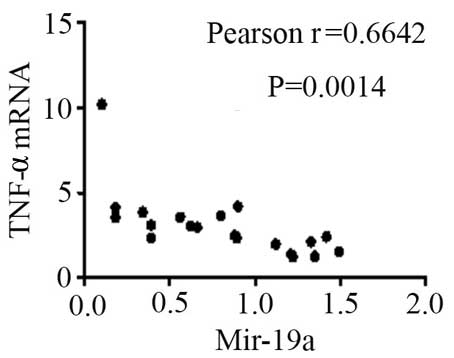
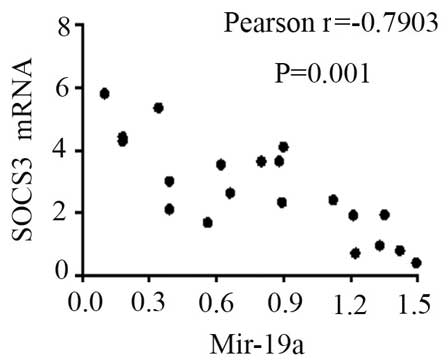
expression levels of TNF-α (Fig. 8)
and SOCS3 (Fig. 9) mRNA were
negatively correlated with miR-19a 2−ΔΔCq (r=0.6642;
P=0.0014 and r=−0.7903; P=0.001, respectively).
Discussion
Continuous or overexpression of the components of
the JAK/STAT signaling pathway are the primary factors involved in
the pathogenic mechanism underlying SJIA (18). IL-6 binds to the JAK receptor which
results in the phosphorylation of STAT3, leading to the
transcription of associated inflammatory genes, induced by
downstream genes (18). Activated
STAT3 is able to promote the differentiation of T-helper (Th)17
cells, induce high levels of receptor activator of nuclear
factor-κB ligand protein expression in synovioblasts (19), and stimulate the production of
inflammatory cytokines, including TNF-α, IL-1 and IL-6 (20). In addition, activated STAT3 is able
to induce osteoclast formation and promote joint destruction
(21).
The results from the present study suggest that
STAT3 is expressed at high levels in patients with SJIA. miR-21
expression is upregulated in the majority of tumors, and may induce
the transcription of STAT3 (22).
Recently, numerous studies have suggested that miR21 participates
in the development of inflammation, and is associated with the
maturation and differentiation of T cells (23–26).
miR-21 serves an important regulatory role in the interaction of
Th1 and Th2 cells. A previous study demonstrated that the secretion
of IL-12, IFN-α and IL-4 was decreased in murine Th cells lacking
miR-21 following the induction of lipopolysaccharide, and may
promote Th1 delayed hypersensitivity (23); the transcription repressor B cell
lymphoma 6 (Bcl6) is an important regulator of Th cells, and could
inhibit Th2-type inflammation (27).
At present, it is thought that miR-21 is the target of Bcl16, and
is able to downregulate Bcl16 expression and activate STAT3
(27). In addition, STAT3 is thought
to upregulate miR-21 expression by binding to the specific miR-21
promoter site. It may therefore be hypothesized that miR-21 can
promote the differentiation of Th2 cells (27).
Wang et al (27) demonstrated that in a transplanted
tumor model, the knockdown of the miR-21 gene can inhibit the
growth of a tumor, and inhibit the expression and phosphorylation
of STAT3. In patients with RA, the expression levels of miR-21 gene
are low, and those of STAT5/phosphorylated STAT5 proteins and
forkhead box P3 mRNA were decreased following the expression and
activation of STAT3, in which the negative feedback mechanism
influenced the cell balance of Th17/Treg.
miR-19a may be involved in the development of SJIA
by regulating the expression of SOCS3 and TNF-α. miR-19a is able to
inhibit the expression of SOCS3, enhance the signal transduction of
the JAK/STAT3 signaling pathway and promote the transcription of
transcription factors (28). Collins
et al (28) demonstrated
using qPCR that miR-19a is able to regulate the JAK/STAT signaling
pathway. The target genes of miR-19a, including the signaling
factors of the JAK/STAT signaling pathway, were identified using
bioinformatics to be SOCS1, SOCS3, SOCS5 and cullin 5 (28). A functional study demonstrated that
miR-19a is able to downregulate the expression of SOCS3 mRNA and
its proteins, whereas an antagonist of miR-19a is able to
significantly reverse this inhibition (28). In addition, miR-19a decreases the
expression levels of SOCS3, and enhances the transduction of IFN-α
and IL-6/STAT3 (29). These results
suggest that miR-19a positively regulates the JAK/STAT signaling
pathway, and influences the incidence and development of
inflammation.
Numerous studies demonstrated that miR-19a serves a
negative regulatory role in inflammation. Philippe et al
(30) reported that toll-like
receptor 2 (TLR2) was highly expressed in the synovial cells of RA
induced by LPS. In an miRNA chip, miR-19a/b was demonstrated to be
downregulated in RA synovial cells, and miR-19a mimics were
demonstrated to decrease the expression levels of TLR2, IL-6 and
MMP3. Gantier et al (31)
reported that miR-19a was able to inhibit A20/Tnfaip3, Rnf11,
Fbxl11/Kdm2a, and Zbtb16, and promote the transduction of nuclear
factor-κB, and that the downregulation of Rnf11 expression could
decrease the expression levels of IL-6 and IL-8. In colitis tissue
samples, miR-19a expression was downregulated, and the expression
levels of TNF-α were increased (32). However, in the colitis mouse model, a
luciferase vector demonstrated that TNF-α is the target gene of
miR-19a (33).
The present study demonstrated that the relative
expression levels of miR-19a in PBMCs of patients with SJIA were
significantly lower compared with the control subjects. In
addition, the mRNA of miR-19a target proteins, such as TNF-α and
SOCS3, were highly expressed in patients with SJIA, and their
expression was negatively correlated with miR-19a 2−ΔΔCq
values (P<0.05). These results suggest that the low expression
levels of miR-19a in patients with SJIA contribute towards the high
expression levels of TNF-α, indirectly resulting in the increased
expression levels of IL-6. Furthermore, SOCS3 was the negative
feedback protein involved in the JAK/STAT signaling pathway, but
high expression levels of SOCS3 did not downregulate the activation
of STAT3; this may be associated with the functional disorder of
SOCS3. These findings remain to be further studied.
An increasing number of miRNAs are being identified,
and are thought to be closely associated with numerous diseases,
resulting from their regulatory effect on signaling pathways
(34). Investigation into the
regulatory mechanism underlying the effect of miRNAs on the
JAK/STAT signaling pathway may provide novel biomarkers for the
diagnosis of SJIA. Further studies are required to examine the
sites of action and effects of miR-21 and miR-19a on their target
proteins, in order to identify their role in the immune mechanisms
underlying SJIA.
Acknowledgements
The present study was funded by the Technology
Project of Guangdong Province (grant no. 2014A020212010).
References
|
1
|
Beukelman T: Treatment advances in
systemic juvenile idiopathic arthritis. F1000Prime Rep. 6:212014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roderburg C and Luedde T: Circulating
microRNAs as markers of liver inflammation, fibrosis and cancer. J
Hepatol. 61:1434–1437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kemper AR, Van Mater HA, Coeytaux RR,
Williams JW Jr and Sanders GD: Systematic review of
disease-modifying antirheumatic drugs for juvenile idiopathic
arthritis. BMC Pediatr. 12:292012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tian TJ, Wang J and Zhou X: A review:
microRNA detection methods. Org Biomol Chem. 13:2226–2238. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kurowska-Stolarska M, Alivernini S,
Ballantine LE, Asquith DL, Millar NL, Gilchrist DS, Reilly J, Ierna
M, Fraser AR, Stolarski B, et al: MicroRNA-155 as a proinflammatory
regulator in clinical and experimental arthritis. Proc Natl Acad
Sci USA. 108:11193–11198. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou Q, Haupt S, Kreuzer JT, Hammitzsch A,
Proft F, Neumann C, Leipe J, Witt M, Schulze-Koops H and Skapenko
A: Decreased expression of miR-146a and miR-155 contributes to an
abnormal Treg phenotype in patients with rheumatoid arthritis. Ann
Rheum Dis. 74:1265–1274. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie Q, Wang SC, Zhong J and Li J:
MicroRNA-146a, a good biomarker and potential therapeutic target
for rheumatoid arthritis. Genet Test Mol Biomarkers. 17:91–92.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Long L, Yu P, Liu Y, Wang S, Li R, Shi J,
Zhang X, Li Y, Sun X, Zhou B, et al: Upregulated microRNA-155
expression in peripheral blood mononuclear cells and
fibroblast-like synoviocytes in rheumatoid arthritis. Clin Dev
Immunol. 2013:2961392013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen SY: MicroRNA-223: A double-edged
sword in rheumatoid arthritis. Rheumatol Int. 34:285–286. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong L, Wang X, Tan J, Li H, Qian W, Chen
J, Chen Q, Wang J, Xu W, Tao C and Wang S: Decreased expression of
microRNA-21 correlates with the imbalance of Th17 and Treg cells in
patients with rheumatoid arthritis. J Cell Mol Med. 18:2213–2224.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shibuya H, Nakasa T, Adachi N, Nagata Y,
Ishikawa M, Deie M, Suzuki O and Ochi M: Overexpression of
microRNA-223 in rheumatoid arthritis synovium controls osteoclast
differentiation. Mod Rheumatol. 23:674–685. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng ZT, Li J, Ren J and Lv Z: Expression
of miR-146a and miR-16 in peripheral blood mononuclear cells of
patients with rheumatoid arthritis and their correlation to the
disease activity. Nan Fang Yi Ke Da Xue Xue Bao. 31:320–323.
2011.(In Chinese). PubMed/NCBI
|
|
13
|
Stanczyk J, Ospelt C, Karouzakis E, Filer
A, Raza K, Kolling C, Gay R, Buckley CD, Tak PP, Gay S and Kyburz
D: Altered expression of microRNA-203 in rheumatoid arthritis
synovial fibroblasts and its role in fibroblast activation.
Arthritis Rheum. 63:373–381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu S, Pan W, Song X, Liu Y, Shao X, Tang
Y, Liang D, He D, Wang H, Liu W, et al: The microRNA miR-23b
suppresses IL-17-associated autoimmune inflammation by targeting
TAB2, TAB3 and IKK-α. Nat Med. 18:1077–1086. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
ILAR 2001. Abstracts of the 20th Congress
of the International League of Associations for Rheumatology.
Edmonton, Alberta, Canada. August 26–30, 2001. J Rheumatol Suppll.
63:1–120. 2001.
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao W, Dong Y, Wu C, Ma Y, Jin Y and Ji
Y: MiR-21 overexpression improves osteoporosis by targeting RECK.
Mol Cell Biochem. 405:125–133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sasi W, Sharma AK and Mokbel K: The role
of suppressors of cytokine signalling in human neoplasms. Mol Biol
Int. 2014:6307972014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ju JH, Heo YJ, Cho ML, Jhun JY, Park JS,
Lee SY, Oh HJ, Moon SJ, Kwok SK, Park KS, et al: Modulation of
STAT3 in rheumatoid synovial T cells suppresses Th17
differentiation and increases the proportion of Treg cells.
Arthritis Rheum. 64:3543–3552. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garbers C, Aparicio-Siegmund S and
Rose-John S: The IL-6/gp130/STAT3 signaling axis: Recent advances
towards specific inhibition. Curr Opin Immunol. 34:75–82. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kotake S, Udagawa N, Takahashi N,
Matsuzaki K, Itoh K, Ishiyama S, Saito S, Inoue K, Kamatani N,
Gillespie MT, et al: IL-17 in synovial fluids from patients with
rheumatoid arthritis is a potent stimulator of osteoclastogenesis.
J Clin Invest. 103:1345–1352. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sawant DV, Wu H, Kaplan MH and Dent AL:
The Bcl6 target gene microRNA-21 promotes Th2 differentiation by a
T cell intrinsic pathway. Mol Immunol. 54:435–442. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu TX, Hartner J, Lim EJ, Fabry V, Mingler
MK, Cole ET, Orkin SH, Aronow BJ and Rothenberg ME: MicroRNA-21
limits in vivo immune response-mediated activation of the
IL-12/IFN-gamma pathway, Th1 polarization and the severity of
delayed-type hypersensitivity. J Immunol. 187:3362–3373. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Iliopoulos D, Jaeger SA, Hirsch HA, Bulyk
ML and Struhl K: STAT3 activation of miR-21 and miR-181b-1 via PTEN
and CYLD are part of the epigenetic switch linking inflammation to
cancer. Mol Cell. 39:493–506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park HK, Jo W, Choi HJ, Jang S, Ryu JE,
Lee HJ, Lee H, Kim H, Yu ES and Son WC: Time-course changes in the
expression levels of miR-122, −155, and −21 as markers of liver
cell damage, inflammation, and regeneration in
acetaminophen-induced liver injury in rats. J Vet Sci. 4:64–71.
2015.
|
|
26
|
Peacock O, Lee AC, Cameron F, Tarbox R,
Vafadar-Isfahani N, Tufarelli C and Lund JN: Inflammation and
MiR-21 pathways functionally interact to downregulate PDCD4 in
colorectal cancer. PLoS One. 9:e1102672014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang YY, Sun G, Luo H, Wang XF, Lan FM,
Yue X, Fu LS, Pu PY, Kang CS, Liu N and You YP: MiR-21 modulates
hTERT through a STAT3-dependent manner on glioblastoma cell growth.
CNS Neurosci Ther. 18:722–728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Collins AS, McCoy CE, Lloyd AT, O'Farrelly
C and Stevenson NJ: miR-19a: An effective regulator of SOCS3 and
enhancer of JAK-STAT signalling. PLoS One. 8:e690902013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qin S, Ai F, Ji WF, Rao W, Zhang HC and
Yao WJ: miR-19a promotes cell growth and tumorigenesis through
targeting SOCS1 in gastric cancer. Asian Pac J Cancer Prev.
14:835–840. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Philippe L, Alsaleh G, Suffert G, Meyer A,
Georgel P, Sibilia J, Wachsmann D and Pfeffer S: TLR2 expression is
regulated by MicroRNA miR-19 in rheumatoid fibroblast-like
synoviocytes. J Immunol. 188:454–461. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gantier MP, Stunden HJ, McCoy CE, Behlke
MA, Wang D, Kaparakis-Liaskos M, Sarvestani ST, Yang YH, Xu D, Corr
SC, et al: A miR-19 regulon that controls NF-kB signaling. Nucleic
Acids Res. 40:8048–8058. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen B, She S, Li D, Liu Z, Yang X, Zeng Z
and Liu F: Role of miR-19a targeting TNF-α in mediating ulcerative
colitis. Scand J Gastroenterol. 48:815–824. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou P, Chen B, Hu P and Sun Y: Role of
miR-19a in ulcerative colitis in mice. Nan Fang Yi Ke Da Xue Xue
Bao. 33:1325–1328. 2013.(In Chinese). PubMed/NCBI
|
|
34
|
Mitchell PS, Parkin RK, Kroh EM, Fritz BR,
Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Notebook J, O'Brian
KC, et al: Circulating microRNAs as stableblood-based markers for
cancer detection. Proc Natl Acad Sci USA. 105:10513–10518. 2008.
View Article : Google Scholar : PubMed/NCBI
|