Introduction
Chronic cerebral hypoperfusion (CCH), which has
previously been associated with the onset of vascular dementia and
cognitive dysfunction (1,2), induces oxidative stress and
inflammatory reactions that may contribute to cognitive impairment
and neuronal damage in various regions of the brain, including the
hippocampus and cerebral cortex (1,3–7). Previous reports of CCH-induced
neuropathological alterations have focused on the hippocampus,
likely due to its critical involvement in learning and memory
(6–9). In addition, the hippocampal CA1 region
was shown to be vulnerable to the effects of CCH and cerebral
ischemia (6–11). A rat model of permanent bilateral
common carotid artery occlusion (2VO) is widely used to investigate
CCH and CCH-related vascular dementia, as the 2VO procedure results
in a significant reduction in cerebral blood flow (4,6,7,9).
Furthermore, previous studies have suggested that an association
exists between 2VO-induced cognitive impairment and hippocampal
damage (1,4,7,12,13).
Melatonin (N-acetyl-5-methoxytryptamine), which is a
well-known and potent free radical scavenger and antioxidant,
sequesters hydroxyl radicals and stimulates antioxidative enzymes
(14,15). Furthermore, melatonin has been
reported to exert neuroprotective effects in various animal models
of neurodegenerative diseases (16,17),
including reducing the infarct volume and neurological deficit in
an animal model of focal cerebral ischemia, and attenuating
hippocampal CA1 region pyramidal neuronal cell death/damage in
various animal models of global cerebral ischemia (10,18,19). The
neuroprotective effects of melatonin have previously been
associated with decreased oxidative stress and neuroinflammation
(20,21).
Ozacmak et al (6) reported that melatonin was able to
protect hippocampal neurons against the effects of CCH by reducing
oxidative stress and heat shock protein 70 expression levels in an
ovariectomized rat; however, the underlying mechanisms by which
melatonin may protect against CCH-induced cognitive impairment and
neuropathological alterations are yet to be fully elucidated. The
present study aimed to investigate the neuroprotective effects of
melatonin against CCH-induced cognitive impairment and neuronal
damage/death in a rat model of 2VO-induced CCH.
Materials and methods
Experimental rats
A total of 48 male Sprague-Dawley rats (age, 16
weeks; weight, 350±20 g) were obtained from RaonBio Inc. (Yongin,
South Korea). The rats were housed in individual cages
(temperature, 23°C; humidity, 60%) under a 12 h light/dark cycle,
and provided with ad libitum access to water and commercial
chow throughout the experimental period. The rat handling and care
procedures adhered to guidelines that are in compliance with
current international laws and policies (22), and the experimental protocol was
approved by the Institutional Animal Care and Use Committee of
Kangwon National University (Chuncheon, South Korea). All
experiments performed were designed to minimize the number of rats
used and their suffering.
Experimental groups and melatonin
treatment
In order to investigate the effects of melatonin
(Sigma-Aldrich, St. Louis, MO, USA) on CCH-induced cognitive
impairment and hippocampal neuronal damage, the rats were
designated into three groups (n=16/group), as follows: i) A
vehicle-treated (1% ethanol in saline), sham-operated group
(sham-group); ii) a vehicle-treated, 2VO-operated group
(2VO-group); and iii) a 10 mg/kg melatonin-treated, 2VO-operated
group (mel-2VO-group). Melatonin, dissolved in ethanol and saline
(final concentration, 1%), was intraperitoneally administered once
daily, between days 1 (surgery) and 28 (sacrifice). The dose of
melatonin administered to the rats was selected on the basis of
previous studies (6,10).
Production of a rat CCH model
A rat model of CCH was generated via 2VO, as
outlined in previous studies (4,6,7). Briefly, the rats were anesthetized
using a mixture of 2.5% isoflurane (Baxter, Deerfield, IL, USA),
33% oxygen and 67% nitrous oxide. A midline cervical incision was
used to expose the bilateral common carotid arteries and to
separate them from the vagus nerve. In the 2VO-group and
mel-2VO-group rats, the bilateral common carotid arteries were
ligated using a 5-0 silk suture. The body temperatures of the rats
under free-regulating or normothermic (37±0.5°C) conditions were
monitored using rectal temperature probes (TR-100; Fine Science
Tools, Foster City, CA, USA), and were maintained using a
thermometric blanket prior to, during and following surgery, until
the rats had recovered from the anesthesia. Thereafter, the animals
were maintained in a thermal incubator (Mirae Medical Industry,
Seoul, Korea), prior to sacrifice. The sham-group rats were
subjected to an identical surgical procedure; however, the
bilateral common carotid arteries were not occluded. All rats that
did not survive the surgery were replaced.
Morris water maze test
The spatial learning and memory functions of all
rats were evaluated using a Morris water maze test, as outlined in
previous studies (4,7,8,23). Briefly, the maze consisted of a black
circular tank (diameter, 120 cm; height, 50 cm) filled with water
(temperature, 23°C; depth, 40 cm). A black platform was submerged
1.5 cm beneath the water surface throughout the duration of the
test. The rats were allowed a maximum of 120 sec to locate the
platform, during which the escape latencies and swimming paths onto
the platform were recorded using a video camera linked to a
computer via using the SMART video tracking system (Panlab,
Barcelona, Spain). All rats were subjected to four trials per day
for five consecutive days (between days 23 and 27 following
surgery).
Tissue processing and neuronal
damage
The rats (n=6/group) were anesthetized using 30%
chloral hydrate (10 ml/kg; Sigma-Aldrich), 28 days following
surgery, after which they were treated with 0.1 M
phosphate-buffered saline (PBS; pH 7.4), and then 4%
paraformaldehyde in 0.1 M phosphate-buffer (pH 7.4), via a
transcardial perfusion. Subsequently, the brains were removed and
postfixed in 4% paraformaldehyde for 6 h, after which the brain
tissues were cryoprotected via incubation with 30% sucrose
overnight. Thereafter, frozen tissue samples were serially
sectioned using a cryostat (Leica Microsystems GmbH, Wetzlar,
Germany) into 30 µm coronal sections, which were subsequently
distributed into 6-well plates containing PBS.
In order to examine neuronal damage and the
neuroprotective effects of melatonin in the hippocampal CA1 region,
Neuronal Nuclei (NeuN) immunohistochemistry and Fluoro-Jade B (F-J
B) histofluorescence staining were conducted, as outlined in
previous studies (10,23,24).
Briefly, the tissue sections were treated with 0.3% hydrogen
peroxide in PBS for 30 min, followed by 10% normal goat serum
(Vector Laboratories, Inc., Burlingame, CA, USA) in 0.05 M PBS for
30 min. Subsequently, the tissue sections were incubated with
diluted mouse anti-NeuN (cat. no. MAB377; dilution, 1:1,000;
Chemicon International, Inc., Temecula, CA, USA) overnight at 4°C,
after which they were incubated with streptavidin
peroxidase-conjugated biotinylated goat anti-mouse immunoglobulin G
(cat. no. BA-9200-1.5; dilution, 1:200; Vector Laboratories, Inc.).
For visualization, the tissue sections were stained with
3,3′-diaminobenzidine (Sigma-Aldrich) in 0.1 M Tris-HCl buffer (pH
7.2), and mounted on gelatin-coated slides. Following dehydration,
the tissue sections were mounted with Canada balsam (Kanto
Chemical, Co., Inc., Tokyo, Japan).
For F-J B histofluorescence staining, the tissue
sections were incubated in a 1% sodium hydroxide solution,
supplemented with first 80%, and then 70% ethanol. Subsequently,
the sections were treated with 0.06% potassium permanganate,
followed by staining with 0.0004% F-J B staining solution
(Histo-Chem, Inc., Jefferson, AR, USA). After washing, the sections
were mounted on a slide warmer (temperature, ~50°C), and were
examined using an epifluorescent microscope (Carl Zeiss AG,
Oberkochen, Germany), with a blue (450–490 nm) excitation light and
a barrier filter. Neurons undergoing degeneration exhibited bright
fluorescence, as compared with the background.
In order to evaluate the neuroprotective effects of
melatonin, NeuN-immunoreactive neurons and F-J B-positive cells in
the hippocampal CA1 region were counted (magnification, 40x) and
digital images were captured using an Axio Imager 2 light
microscope (Carl Zeiss AG), equipped with a digital camera
(Axiocam; Carl Zeiss AG) connected to a PC monitor. Six coronal
sections with 150 µm intervals were selected for each rat, and cell
counts were obtained by averaging the counts from each rat.
Measurement of lipid peroxidation
In order to examine the effects of melatonin on the
rates of lipid peroxidation in the hippocampus of the CCH rats,
malondialdehyde (MDA) formation was measured using the
Bioxytech® MDA-586 kit (OxisResearch, Portland, OR,
USA), according to our previous study (23). Briefly, the rats (n=5/group) were
sacrificed 28 days following surgery, after which the hippocampus
was removed and homogenized in 20 mM PBS (pH 7.4) containing 5 mM
butylated hydroxytoluene, using a Vibra-Cell ultrasonic processor
(Sonics and Materials, Inc., Newtown, CT, USA). Homogenates were
centrifuged at 3,000 × g for 10 min at 4°C, and the supernatants
were collected for MDA quantification. Probucol (10 µl) and diluted
R1 reagent (640 µl; 1:3 of methanol:N-methyl-2-phenylindole) were
added to the supernatants and subsequently mixed with 150 µl 12 N
hydrochloric acid. Each reaction was incubated for 60 min at 45°C,
after which the supernatants were centrifuged at 10,000 × g for 10
min. Absorbance at 586 nm was measured using a BioTek ELx800
absorbance microplate reader (Bio-Tek Instruments Inc., Winooski,
VT, USA) as an indication of MDA content.
Microglial activation
In order to investigate microglial activation,
immunohistochemical staining using rabbit anti-ionized
calcium-binding adapter molecule-1 (Iba-1) antibodies (cat. no.
019-19741; 1:200; Wako Pure Chemical Industries, Ltd., Osaka,
Japan) was performed, as outlined in a previous study (25). The density of Iba-1-immunoreactive
structures was evaluated on the basis of optical density (OD),
which was calculated following the transformation of the mean gray
level with ImageJ version 1.42 software (imagej.nih.gov/ij) using the formula: OD=log (256/mean
gray level). The OD of the background was subtracted from areas
adjacent to the measured area. After the background density was
subtracted, a ratio of the OD of the image file was calibrated as a
percentage (relative optical density, ROD) using Adobe Photoshop
version 8.0 (Adobe Systems, San Jose, CA, USA), and then analyzed
using ImageJ software. A ratio of the ROD was calibrated as %, with
the sham-group designated as 100%.
Quantification of the protein
expression levels of pro-inflammatory cytokines
In order to examine the effects of melatonin on the
protein expression levels of specific pro-inflammatory cytokines in
the hippocampus of the CCH rat model, enzyme linked immunosorbent
assays (ELISA) for tumor necrosis factor (TNF)-α and interleukin
(IL)-1β were performed, in accordance with a previous study
(26). Briefly, the rats (n=5/group)
were sacrificed 28 days following surgery, and the hippocampi were
removed and homogenized using a Vibra-Cell ultrasonic processor.
After centrifugation of the homogenates at 14,000 × g for 20 min at
4°C, the supernatant was collected. The levels of TNF-α and IL-1β
were examined using a commercial Invitrogen ELISA kit (Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol.
Statistical analysis
The data are presented as the mean ± standard error
of the mean. Statistical differences were analyzed using two-way
analysis of variance followed by Dunnett's test, using SPSS 12.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
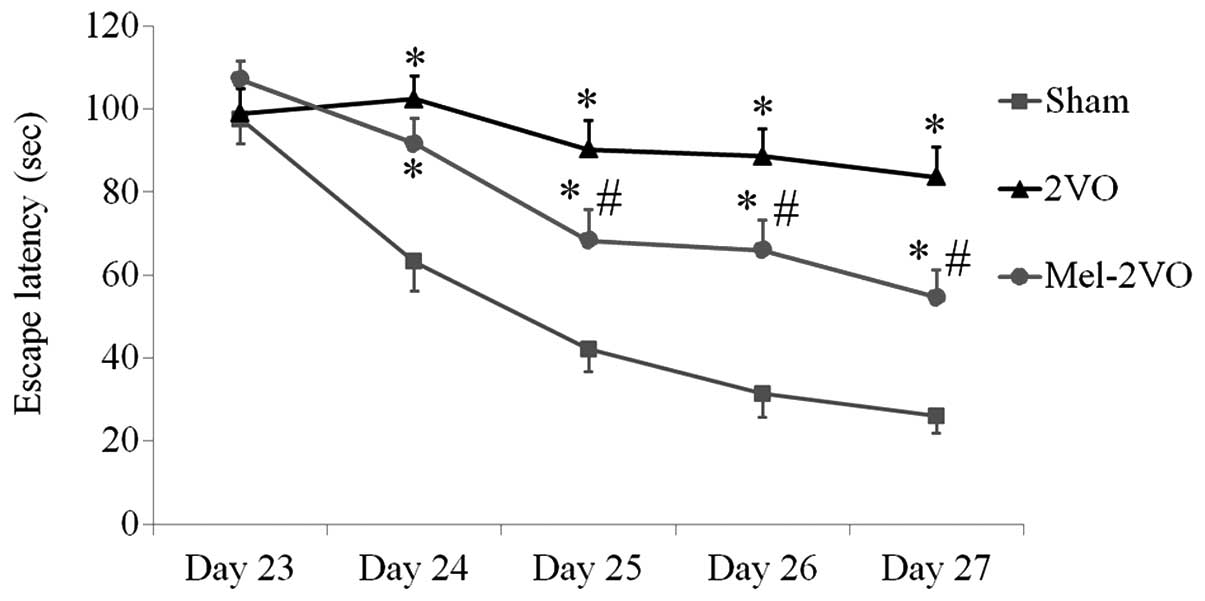
Morris water maze test
The spatial learning and memory functions of all
rats were examined using the Morris water maze test, which was
conducted four times per day between days 23 and 27 following
surgery. The escape latencies for the rats in the sham-group
markedly decreased over the five days, whereas the escape latencies
for the rats in the 2VO-group were significantly longer, as
compared with the rats in the sham-group, and did not significantly
decrease over the trial period (P<0.05, Fig. 1). Conversely, the escape latencies of
the rats in the mel-2VO-group significantly decreased from day 3 in
the trial period, as compared with that in the 2VO-group rats
(P<0.05, Fig. 1). However, the
rats in the mel-2VO-group had markedly longer escape latencies, as
compared with the rats in the sham-group (Fig. 1).
Neuroprotective effects of
melatonin
CCH-induced neuronal cell damage and the
neuroprotective effects of melatonin in the hippocampal CA1 region
were examined 28 days following surgery using NeuN
immunohistochemistry and F-J B histofluorescence staining (Fig. 2). High numbers of NeuN-immunoreactive
(+) cells were detected in the CA1 region of the sham-group rats,
whereas no F-J B-positive (+) cells were detected in this region
(Fig. 2A, D and G). Conversely, in
the 2VO-group rats, the number of NeuN+ cells was significantly
reduced, and the number of F-J B+ cells were markedly increased in
the stratum pyramidale of the hippocampal CA1 region (Fig. 2B, E and G). Elevated numbers of NeuN+
cells were detected in the mel-2VO-group rats: 73.7% of the
pyramidal cells were protected, as compared with that in the
sham-group (Fig. 2C and G). In
addition, the number of F-J B+ cells in the mel-2VO-group was
significantly reduced, as compared with the rats in the 2VO-group
(Fig. 2F and G).
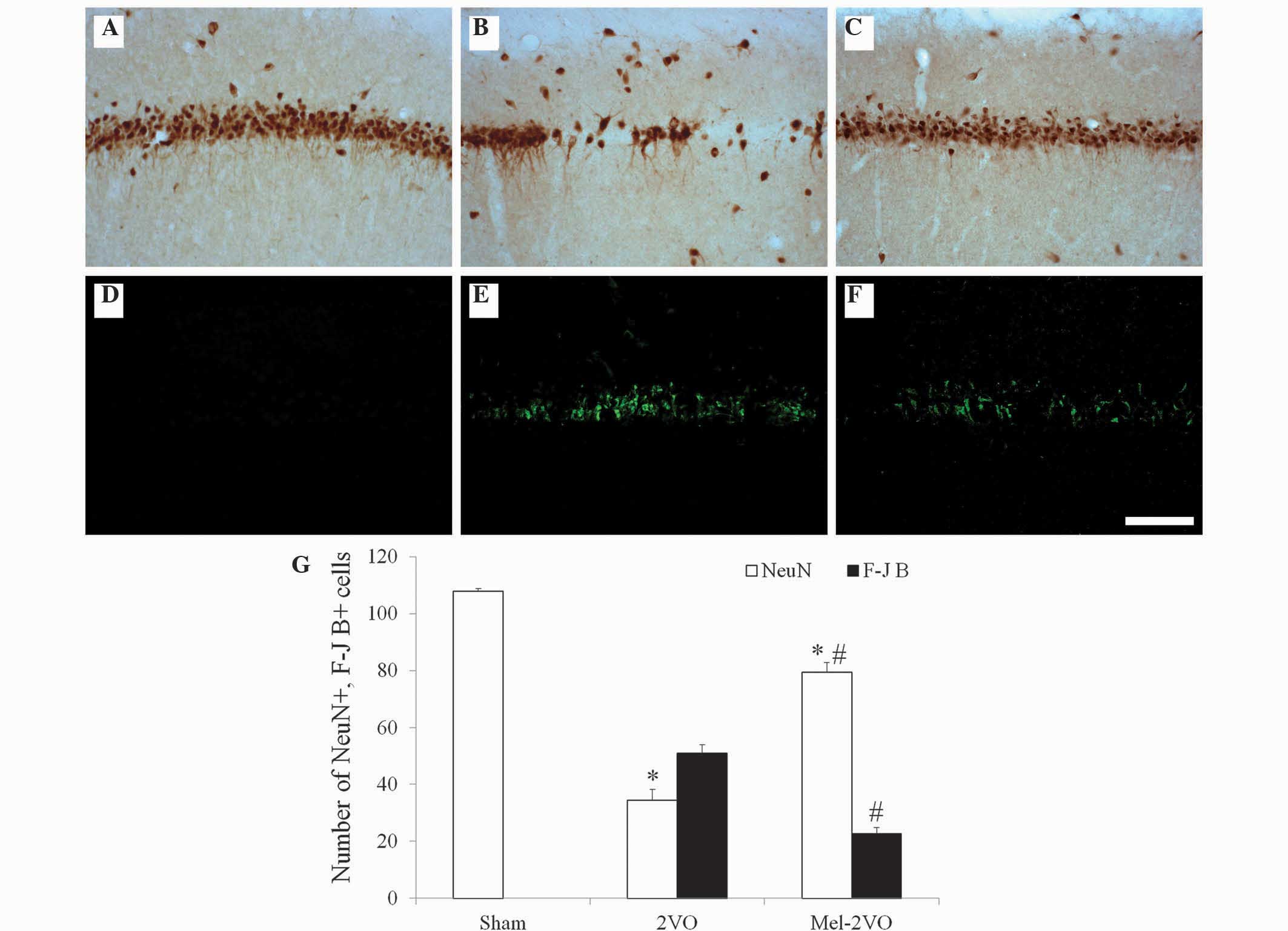 | Figure 2.(A-C) NeuN immunohistochemistry and
(D-F) F-J B histofluorescence staining in the hippocampal CA1
region of the (A and D) sham-, (B and E) 2VO- and (C and F)
mel-2VO- groups. NeuN+ neurons significantly decreased, whereas F-J
B+ cells (arrows) markedly increased, in number in the SP of the
2VO-group rats. Conversely, NeuN+ neurons significantly increased,
and F-J B+ cells significantly decreased, in number in the SP of
the mel-2VO-group rats, as compared with the 2VO-group rats (scale
bar=100 µm). (G) Analysis of the number of NeuN+ neurons and F-J B+
cells in the CA1 region of the sham-, 2VO- and mel-2VO-group rats.
Data are presented as the mean ± standard error of the mean.
*P<0.05 vs. the sham-group, #P<0.05 vs. the
2VO-group. SO, stratum oriens; SR; stratum radiatum; SP, stratum
pyramidale; NeuN, Neuronal Nuclei; F-J B, Fluoro-Jade B; 2VO,
bilateral common carotid arteries occlusion-operated; mel,
melatonin. |
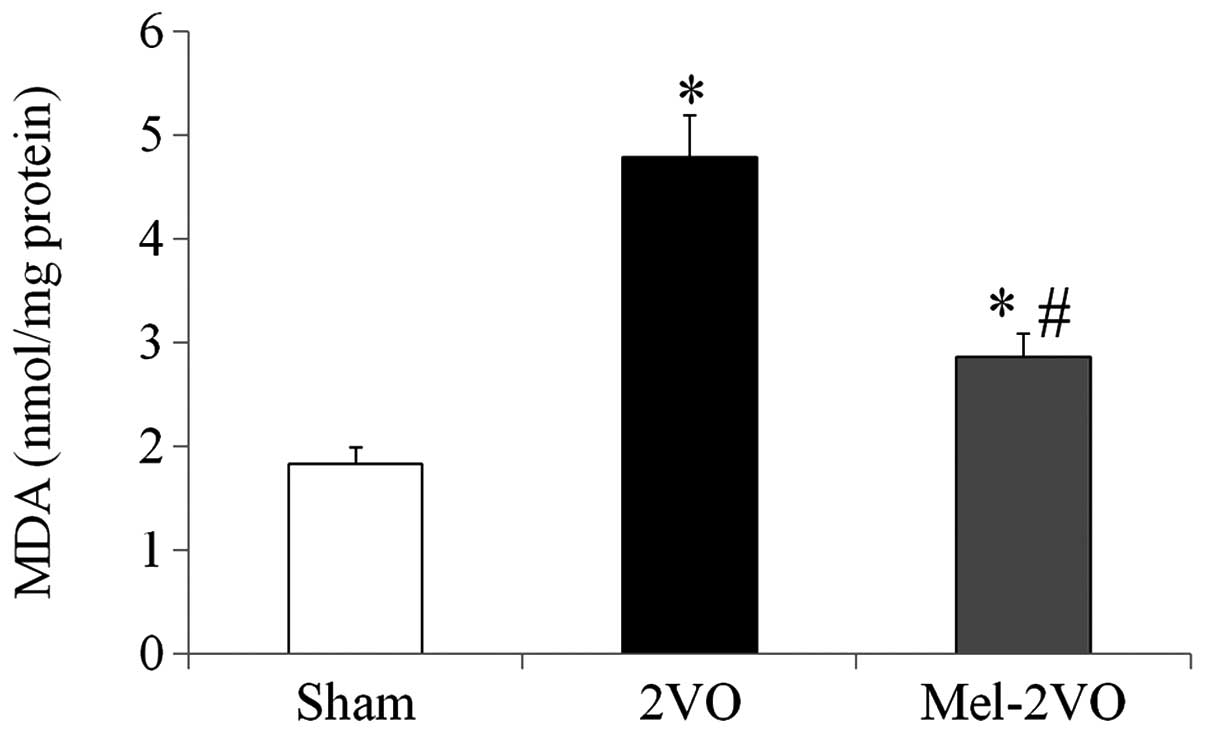
Quantification of MDA levels in the
rat hippocampus
In order to investigate the effects of melatonin on
CCH-induced lipid peroxidation, MDA levels were measured in the
hippocampal samples of all rats. Hippocampal MDA levels in the
sham-group were 1.83±0.16 nmol/mg protein, and were significantly
increased (2.6-fold; P<0.05) in the 2VO-group rats (Fig. 3). However, hippocampal MDA levels in
the mel-2VO-group were significantly reduced (60% of 2VO-group
levels), as compared with the 2VO-group (P<0.05, Fig. 3).
Microglial activation
Iba-1 immunohistochemistry in the hippocampal CA1
region (Fig. 4) demonstrated that,
in the sham-group, ramified Iba-1+ microglia containing a small
cytoplasm, known as resting microglia, were distributed throughout
the hippocampal CA1 region (Fig.
4A). In the CA1 region of the 2VO-group rats, Iba-1
immunoreactivity significantly increased, as compared with the sham
group rats (P<0.05, Fig. 4B and
D). In addition, numerous Iba-1+ microglia exhibited
hypertrophy and activation, with round forms, and were aggregated
in the stratum pyramidale of the CA1 region of the 2VO-group rats
(Fig. 4B). Following treatment with
melatonin, Iba-1 immunoreactivity in the mel-2VO-group
significantly decreased, as compared with the 2VO-group (P<0.05,
Fig. 4C and D); however, a higher
number of microglia exhibited activation in the mel-2VO-group rats,
as compared with the sham-group rats.
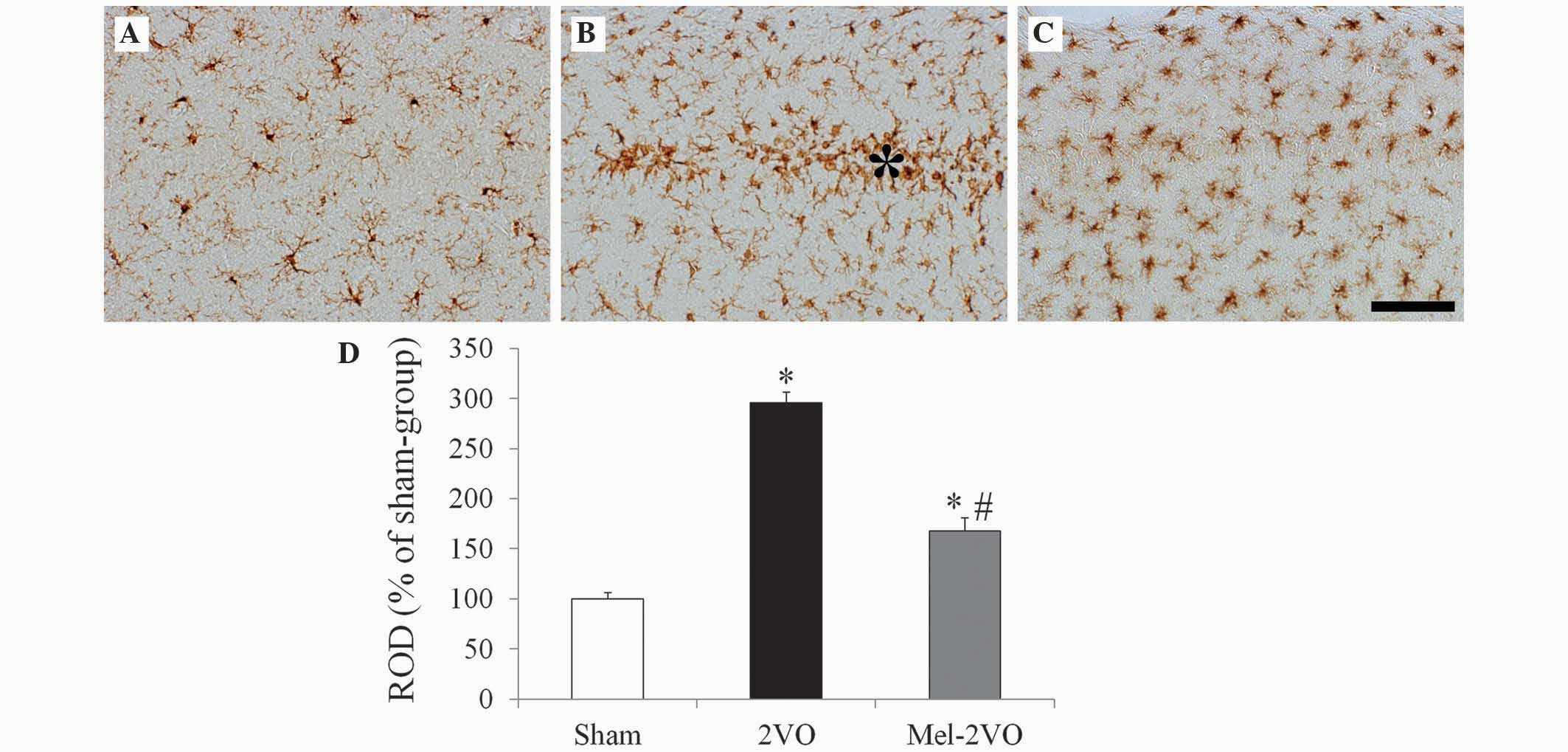 | Figure 4.Iba-1 immunohistochemistry in the
hippocampal CA1 region of the (A) sham-, (B) 2VO- and (C) mel-2VO-
groups. In the 2VO-group, Iba-1 immunoreactivity was markedly
increased, and the activated Iba-1+ microglia aggregated in the SP
(asterisk). In the mel-2VO-group, Iba-1 immunoreactivity was
significantly decreased, as compared with the 2VO-group rats (scale
bar=50 µm). (D) ROD was calculated as the percentage of Iba-1+
structures in the CA1 region of the sham-, 2VO- and mel-2VO-group
rats. Data are presented as the mean ± standard error of the mean.
Asterisk indicates SP. *P<0.05 vs. the sham-group,
#P<0.05 vs. the 2VO-group. Iba-1, ionized
calcium-binding adapter molecule-1; 2VO, bilateral common carotid
arteries occlusion-operated; ROD, relative optical density. |
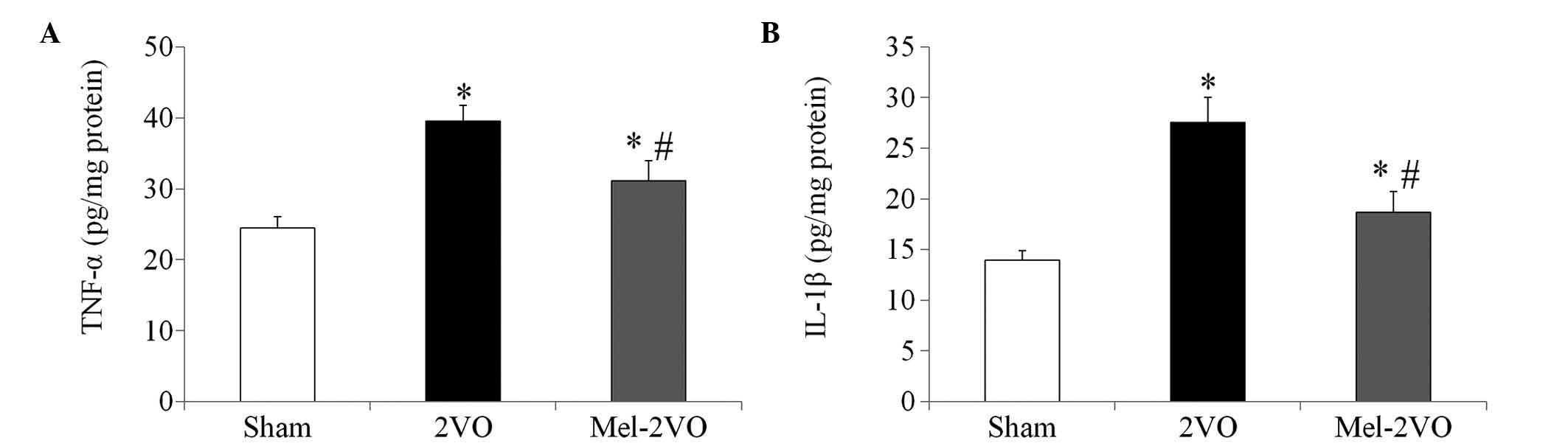
Quantification of TNF-α and IL-1β
protein expression levels
The TNF-α and IL-1β hippocampal protein expression
levels in the sham-group rats were 24.44±1.65 pg/mg and 13.95±0.92
pg/mg protein, respectively. In the 2VO-group, protein expression
levels of TNF-α and IL-1β were significantly increased, as compared
with those in the sham-group rats (P<0.05, Fig. 5). Conversely the TNF-α (31.11±2.85
pg/mg) and IL-1β (18.67±2.05 pg/mg) protein expression levels were
significantly decreased in the mel-2VO-group rats, as compared with
those in the 2VO-group rats (P<0.05, Fig. 5); however, TNF-α and IL-1β protein
expression levels were significantly higher than those in the
sham-group rats (P<0.05, Fig.
5).
Discussion
Previous studies have detected hippocampus-dependent
spatial learning and memory impairment in rat models of 2VO
(1,4,7–9,12,13,27).
The present study examined the effects of melatonin on CCH-induced
cognitive impairment, using the Morris water maze test, and
demonstrated that rats in the 2VO-group had significantly longer
escape latencies, as compared with the rats in the sham-group. In
addition, escape latencies of the rats in the mel-2VO-group were
significantly decreased, as compared with the 2VO-group rats. To
the best of our knowledge, the present study is the first to report
the effects of melatonin on CCH-induced cognitive impairment, and
demonstrated that melatonin treatment was able to improve
CCH-induced hippocampus-dependent spatial learning and memory
impairment. These results are consistent with those of a previous
study, which reported that post-ischemic melatonin treatment in a
rat model of global cerebral ischemia reduced escape latencies, as
compared with a sham-group (19).
Previous studies have detected significant
reductions in the number of pyramidal cells in the hippocampal CA1
region following 2VO, and therefore suggested that 2VO-induced
cognitive impairment may be associated with a loss of hippocampal
CA1 cells (1,7,12,13,27).
In the present study, a significant reduction in the number of
NeuN+ neurons, and a marked increase in the number of F-J B+ (a
marker for degenerating neurons) cells, were detected in the
stratum pyramidale of the hippocampal CA1 region in the 2VO-group
rats. In addition, the ability of melatonin to protect against
CCH-induced neuronal cell death in the hippocampal CA1 region was
investigated using NeuN immunohistochemistry and F-J B
histofluorescence staining. Melatonin treatment significantly
reduced CCH-induced neuronal cell death in the hippocampal CA1
region of 2VO-group rats. Ozacmak et al (6) previously reported that melatonin
treatment attenuated CCH-induced neuropathological alterations in
the hippocampus; concordantly, in the present study, the total
number of cells in the hippocampal CA1 region of rats in the
mel-2VO-group were significantly higher, as compared with in the
2VO-group rats. Therefore, the ability of melatonin to protect
against CCH-induced spatial learning and memory impairment may be
associated with the neuroprotective effects of melatonin in the
hippocampal CA1 region.
Oxidative stress is thought to be a major factor in
CCH-induced neuronal cell death (5,6,28). In the present study, the effects of
melatonin on CCH-induced oxidative stress in the rat hippocampus
were investigated. Melatonin treatment significantly reduced the
CCH-induced elevation of MDA levels in the hippocampus of the
2VO-rats, which is consistent with previous studies that reported
that the neuroprotective effects of melatonin may be associated
with its antioxidant properties in animal models of global and
focal cerebral ischemia (10,14,18).
Furthermore, these results align with a previous study, in which
the protective effects of melatonin against CCH-induced hippocampal
neuronal damage were associated with reduced oxidative stress, as
indicated by restored levels of MDA, superoxide dismutase and
glutathione following melatonin treatment (6).
Morphological and functional microglial alterations
are involved in the response to various changes in the neural
environment, including ischemic insults (25,29). It
was previously reported that CCH initiates significant microglial
activation and neuroinflammatory responses in various regions of
the rat brain, including the hippocampus (1,3,26). Furthermore, it has been demonstrated
that ischemia- and CCH-induced neuroinflammation promote the
expression of pro-inflammatory cytokines, including TNF-α and
IL-1β, and that these have important roles in the progression of
post-ischemic brain injury (26,30). In
addition, activated microglia were shown to secrete
pro-inflammatory cytokines, including TNF-α and IL-1β, in the
central nervous system (29). In our
previous study, we detected neuroprotective effects for melatonin
against ischemic damage, and suggested that these effects may be
associated with attenuation of microglial activation in the
hippocampal CA1 region following transient global cerebral ischemia
(10). Furthermore, previous studies
detected attenuation of transient focal cerebral ischemia- and
lipopolysaccharide-induced neuroinflammation following melatonin
treatment (20,21). In the present study, microglia in the
2VO-group rats were activated and aggregated in proximity to the
stratum pyramidale of the ischemic hippocampal CA1 region, whereas
this behavior was markedly reduced in the mel-2VO-group rats. In
addition, treatment of the 2VO rats with melatonin significantly
decreased the CCH-induced elevation of TNF-α and IL-1β levels in
the hippocampus. To the best of our knowledge, the present study is
the first to demonstrate an anti-inflammatory effect for melatonin
against CCH-induced pro-inflammatory cytokine expression, which
suggests that melatonin treatment may attenuate CCH-induced
neuroinflammation. Therefore, the results of the present study
suggested that the neuroprotective effects of melatonin against
CCH-induced neuronal cell damage may be associated with attenuation
of microglial activation and neuronal inflammatory responses.
In conclusion, the results of the present study
suggested that melatonin was able to improve CCH-induced cognitive
impairment, and that the neuroprotective effects of melatonin
against CCH-induced hippocampal neuronal cell damage were
associated with reduced oxidative stress, attenuation of microglial
activation and decreased production of pro-inflammatory
cytokines.
Acknowledgements
The present study was supported by the Basic Science
Research Program via the National Research Foundation of Korea,
funded by the Ministry of Science, ICT and Future Planning (grant
no. NRF-2012R1A1A1007298).
References
|
1
|
Farkas E, Luiten PG and Bari F: Permanent,
bilateral common carotid artery occlusion in the rat: A model for
chronic cerebral hypoperfusion-related neurodegenerative diseases.
Brain Res Rev. 54:162–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Román GC: Brain hypoperfusion: A critical
factor in vascular dementia. Neurol Res. 26:454–458. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Farkas E, Donka G, de Vos RA, Mihály A,
Bari F and Luiten PG: Experimental cerebral hypoperfusion induces
white matter injury and microglial activation in the rat brain.
Acta Neuropathol. 108:57–64. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
He XL, Wang YH, Bi MG and Du GH: Chrysin
improves cognitive deficits and brain damage induced by chronic
cerebral hypoperfusion in rats. Eur J Pharmacol. 680:41–48. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kasparová S, Brezová V, Valko M, Horecký
J, Mlynárik V, Liptaj T, Vancová O, Ulicná O and Dobrota D: Study
of the oxidative stress in a rat model of chronic brain
hypoperfusion. Neurochem Int. 46:601–611. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ozacmak VH, Barut F and Ozacmak HS:
Melatonin provides neuroprotection by reducing oxidative stress and
HSP70 expression during chronic cerebral hypoperfusion in
ovariectomized rats. J Pineal Res. 47:156–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xi Y, Wang M, Zhang W, Bai M, Du Y, Zhang
Z, Li Z and Miao J: Neuronal damage, central cholinergic
dysfunction and oxidative damage correlate with cognitive deficits
in rats with chronic cerebral hypoperfusion. Neurobiol Learn Mem.
109:7–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cechetti F, Pagnussat AS, Worm PV, Elsner
VR, Ben J, da Costa MS, Mestriner R, Weis SN and Netto CA: Chronic
brain hypoperfusion causes early glial activation and neuronal
death, and subsequent long-term memory impairment. Brain Res Bull.
87:109–116. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feng Z, Lu Y, Wu X, Zhao P, Li J, Peng B,
Qian Z and Zhu L: Ligustilide alleviates brain damage and improves
cognitive function in rats of chronic cerebral hypoperfusion. J
Ethnopharmacol. 144:313–321. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee CH, Yoo KY, Choi JH, Park OK, Hwang
IK, Kwon YG, Kim YM and Won MH: Melatonin's protective action
against ischemic neuronal damage is associated with up-regulation
of the MT2 melatonin receptor. J Neurosci Res. 88:2630–2640.
2010.PubMed/NCBI
|
|
11
|
Zhu B, Wang ZG, Ding J, Liu N, Wang DM,
Ding LC and Yang C: Chronic lipopolysaccharide exposure induces
cognitive dysfunction without affecting BDNF expression in the rat
hippocampus. Exp Ther Med. 7:750–754. 2014.PubMed/NCBI
|
|
12
|
De Jong GI, Farkas E, Stienstra CM, Plass
JR, Keijser JN, de la Torre JC and Luiten PG: Cerebral
hypoperfusion yields capillary damage in the hippocampal CA1 area
that correlates with spatial memory impairment. Neuroscience.
91:203–210. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sarti C, Pantoni L, Bartolini L and
Inzitari D: Cognitive impairment and chronic cerebral
hypoperfusion: What can be learned from experimental models. J
Neurol Sci. 203-204:263–266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reiter RJ, Tan DX, Osuna C and Gitto E:
Actions of melatonin in the reduction of oxidative stress. A
review. J Biomed Sci. 7:444–458. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Li L, Xiang C, Ma Z, Ma T and Zhu
S: Protective effect of melatonin against Adriamycin-induced
cardiotoxicity. Exp Ther Med. 5:1496–1500. 2013.PubMed/NCBI
|
|
16
|
Ma J, Shaw VE and Mitrofanis J: Does
melatonin help save dopaminergic cells in MPTP-treated mice?
Parkinsonism Relat Disord. 15:307–314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reiter RJ, Cabrera J, Sainz RM, Mayo JC,
Manchester LC and Tan DX: Melatonin as a pharmacological agent
against neuronal loss in experimental models of Huntington's
disease, Alzheimer's disease and parkinsonism. Ann NY Acad Sci.
890:471–485. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee EJ, Lee MY, Chen HY, Hsu YS, Wu TS,
Chen ST and Chang GL: Melatonin attenuates gray and white matter
damage in a mouse model of transient focal cerebral ischemia. J
Pineal Res. 38:42–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Letechipía-Vallejo G, López-Loeza E,
Espinoza-González V, González-Burgos I, Olvera-Cortés ME, Moralí G
and Cervantes M: Long-term morphological and functional evaluation
of the neuroprotective effects of post-ischemic treatment with
melatonin in rats. J Pineal Res. 42:138–146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee MY, Kuan YH, Chen HY, Chen TY, Chen
ST, Huang CC, Yang IP, Hsu YS, Wu TS and Lee EJ: Intravenous
administration of melatonin reduces the intracerebral cellular
inflammatory response following transient focal cerebral ischemia
in rats. J Pineal Res. 42:297–309. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tyagi E, Agrawal R, Nath C and Shukla R:
Effect of melatonin on neuroinflammation and acetylcholinesterase
activity induced by LPS in rat brain. Eur J Pharmacol. 640:206–210.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Institute of Laboratory Animal Research,
Committee for the Update of the Guide for the Care and Use of
Laboratory Animals and National Research Council: Guide for the
care and use of laboratory animals (8th). Washington, D.C., USA:
National Academies Press. 2202011.
|
|
23
|
Yoo DY, Kim W, Lee CH, Shin BN, Nam SM,
Choi JH, Won MH, Yoon YS and Hwang IK: Melatonin improves
D-galactose-induced aging effects on behavior, neurogenesis, and
lipid peroxidation in the mouse dentate gyrus via increasing pCREB
expression. J Pineal Res. 52:21–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmued LC and Hopkins KJ: Fluoro-Jade B:
A high affinity fluorescent marker for the localization of neuronal
degeneration. Brain Res. 874:123–130. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sugawara T, Lewén A, Noshita N, Gasche Y
and Chan PH: Effects of global ischemia duration on neuronal,
astroglial, oligodendroglial, and microglial reactions in the
vulnerable hippocampal CA1 subregion in rats. J Neurotrauma.
19:85–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qu J, Zhou Q, Du Y, Zhang W, Bai M, Zhang
Z, Xi Y, Li Z and Miao J: Rutin protects against cognitive deficits
and brain damage in rats with chronic cerebral hypoperfusion. Br J
Pharmacol. 171:3702–3715. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pappas BA, de la Torre JC, Davidson CM,
Keyes MT and Fortin T: Chronic reduction of cerebral blood flow in
the adult rat: Late-emerging CA1 cell loss and memory dysfunction.
Brain Res. 708:50–58. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Y, He Y, Guan Q, Liu W, Han H and Nie
Z: Disrupted iron metabolism and ensuing oxidative stress may
mediate cognitive dysfunction induced by chronic cerebral
hypoperfusion. Biol Trace Elem Res. 150:242–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stoll G, Jander S and Schroeter M:
Inflammation and glial responses in ischemic brain lesions. Prog
Neurobiol. 56:149–171. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dirnagl U, Iadecola C and Moskowitz MA:
Pathobiology of ischaemic stroke: An integrated view. Trends
Neurosci. 22:391–397. 1999. View Article : Google Scholar : PubMed/NCBI
|